

Sirtuin 1 in Host Defense during Infection

Abstract
:1. Introduction
2. Overview of SIRTs
3. Function of SIRT1 during Infections
3.1. SIRT1 in Bacterial Infections
3.2. SIRT1 in Viral Infections
3.3. SIRT1 in Parasitic Infections
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nogueiras, R.; Habegger, K.M.; Chaudhary, N.; Finan, B.; Banks, A.S.; Dietrich, M.O.; Horvath, T.L.; Sinclair, D.A.; Pfluger, P.T.; Tschop, M.H. Sirtuin 1 and sirtuin 3: Physiological modulators of metabolism. Physiol. Rev. 2012, 92, 1479–1514. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.A.; Madsen, A.S.; Olsen, C.A.; Hirschey, M.D. Metabolic control by sirtuins and other enzymes that sense NAD+, NADH, or their ratio. Biochim. Biophys. Acta Bioenerg. 2017, 1858, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, H.B.S.; Williams, C.; King, S.J.; Allison, S.J. Nicotinamide adenine dinucleotide (NAD+): Essential redox metabolite, co-substrate and an anti-cancer and anti-ageing therapeutic target. Biochem. Soc. Trans. 2020, 48, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Figarola-Centurion, I.; Escoto-Delgadillo, M.; Gonzalez-Enriquez, G.V.; Gutierrez-Sevilla, J.E.; Vazquez-Valls, E.; Torres-Mendoza, B.M. Sirtuins Modulation: A Promising Strategy for HIV-Associated Neurocognitive Impairments. Int. J. Mol. Sci. 2022, 23, 643. [Google Scholar] [CrossRef] [PubMed]
- Sadia, K.; Ashraf, M.Z.; Mishra, A. Therapeutic Role of Sirtuins Targeting Unfolded Protein Response, Coagulation, and Inflammation in Hypoxia-Induced Thrombosis. Front. Physiol. 2021, 12, 733453. [Google Scholar] [CrossRef]
- Park, S.; Mori, R.; Shimokawa, I. Do sirtuins promote mammalian longevity? A critical review on its relevance to the longevity effect induced by calorie restriction. Mol. Cells 2013, 35, 474–480. [Google Scholar] [CrossRef]
- Hamaidi, I.; Kim, S. Sirtuins are crucial regulators of T cell metabolism and functions. Exp. Mol. Med. 2022, 54, 207–215. [Google Scholar] [CrossRef]
- Teixeira, C.S.S.; Cerqueira, N.; Gomes, P.; Sousa, S.F. A Molecular Perspective on Sirtuin Activity. Int. J. Mol. Sci. 2020, 21, 8609. [Google Scholar] [CrossRef]
- Hawse, W.F.; Wolberger, C. Structure-based mechanism of ADP-ribosylation by sirtuins. J. Biol. Chem. 2009, 284, 33654–33661. [Google Scholar] [CrossRef]
- Bringman-Rodenbarger, L.R.; Guo, A.H.; Lyssiotis, C.A.; Lombard, D.B. Emerging Roles for SIRT5 in Metabolism and Cancer. Antioxid. Redox Signal. 2018, 28, 677–690. [Google Scholar] [CrossRef]
- Lin, L.; Wen, S.H.; Guo, S.Z.; Su, X.Y.; Wu, H.J.; Chong, L.; Zhang, H.L.; Zhang, W.X.; Li, C.C. Role of SIRT1 in Streptococcus pneumoniae-induced human beta-defensin-2 and interleukin-8 expression in A549 cell. Mol. Cell. Biochem. 2014, 394, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Bai, Z.; Li, C.; Sheng, C.; Zhao, X. EV71 infection induces cell apoptosis through ROS generation and SIRT1 activation. J. Cell. Biochem. 2020, 121, 4321–4331. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, B.; Gao, W.; An, Y.; Dong, G.; Jia, J.; Yang, Q. Helicobacter pylori inhibits autophagic flux and promotes its intracellular survival and colonization by down-regulating SIRT1. J. Cell. Mol. Med. 2021, 25, 3348–3360. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Hu, J.; Chen, Y.J.; Ge, B. Role of Sirt1 in innate immune mechanisms against Mycobacterium tuberculosis via the inhibition of TAK1 activation. Arch. Biochem. Biophys. 2019, 667, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Hale, B.G. Antiviral immunity triggered by infection-induced host transposable elements. Curr. Opin. Virol. 2022, 52, 211–216. [Google Scholar] [CrossRef]
- King, A.N.; de Mets, F.; Brinsmade, S.R. Who’s in control? Regulation of metabolism and pathogenesis in space and time. Curr. Opin. Microbiol. 2020, 55, 88–96. [Google Scholar] [CrossRef]
- Gan, J.; Giogha, C.; Hartland, E.L. Molecular mechanisms employed by enteric bacterial pathogens to antagonise host innate immunity. Curr. Opin. Microbiol. 2021, 59, 58–64. [Google Scholar] [CrossRef]
- Davies, L.C.; Jenkins, S.J.; Allen, J.E.; Taylor, P.R. Tissue-resident macrophages. Nat. Immunol. 2013, 14, 986–995. [Google Scholar] [CrossRef]
- Barnaba, V. T Cell Memory in Infection, Cancer, and Autoimmunity. Front. Immunol. 2021, 12, 811968. [Google Scholar] [CrossRef]
- Roy, R.K.; Yadav, R.; Jain, A.; Tripathi, V.; Jain, M.; Singh, S.; Prakash, H. Yin and yang of immunological memory in controlling infections: Overriding self defence mechanisms. Int. Rev. Immunol. 2022, 41, 240–252. [Google Scholar] [CrossRef]
- Janssen, E.M.; Lemmens, E.E.; Wolfe, T.; Christen, U.; von Herrath, M.G.; Schoenberger, S.P. CD4+ T cells are required for secondary expansion and memory in CD8+ T lymphocytes. Nature 2003, 421, 852–856. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Buechler, N.L.; Woodruff, A.G.; Long, D.L.; Zabalawi, M.; Yoza, B.K.; McCall, C.E.; Vachharajani, V. Sirtuins and Immuno-Metabolism of Sepsis. Int. J. Mol. Sci. 2018, 19, 2738. [Google Scholar] [CrossRef] [PubMed]
- Li, X. SIRT1 and energy metabolism. Acta. Biochim. Biophys. Sin. 2013, 45, 51–60. [Google Scholar] [CrossRef]
- Tanno, M.; Sakamoto, J.; Miura, T.; Shimamoto, K.; Horio, Y. Nucleocytoplasmic shuttling of the NAD+-dependent histone deacetylase SIRT1. J. Biol. Chem. 2007, 282, 6823–6832. [Google Scholar] [CrossRef] [PubMed]
- Byles, V.; Chmilewski, L.K.; Wang, J.; Zhu, L.; Forman, L.W.; Faller, D.V.; Dai, Y. Aberrant cytoplasm localization and protein stability of SIRT1 is regulated by PI3K/IGF-1R signaling in human cancer cells. Int. J. Biol. Sci. 2010, 6, 599–612. [Google Scholar] [CrossRef]
- Pande, S.; Raisuddin, S. Molecular and cellular regulatory roles of sirtuin protein. Crit. Rev. Food Sci. Nutr. 2022, 1–19. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, Y.; Wang, Y.; Chao, Y.; Zhang, J.; Jia, Y.; Tie, J.; Hu, D. Regulation of SIRT1 and Its Roles in Inflammation. Front. Immunol. 2022, 13, 831168. [Google Scholar] [CrossRef]
- Tang, Z.; Wang, P.; Dong, C.; Zhang, J.; Wang, X.; Pei, H. Oxidative Stress Signaling Mediated Pathogenesis of Diabetic Cardiomyopathy. Oxid. Med. Cell. Longev. 2022, 2022, 5913374. [Google Scholar] [CrossRef]
- Shen, P.; Deng, X.; Chen, Z.; Ba, X.; Qin, K.; Huang, Y.; Huang, Y.; Li, T.; Yan, J.; Tu, S. SIRT1: A Potential Therapeutic Target in Autoimmune Diseases. Front. Immunol. 2021, 12, 779177. [Google Scholar] [CrossRef]
- Xu, Y.; Wan, W. Acetylation in the regulation of autophagy. Autophagy 2022, 1–9. [Google Scholar] [CrossRef]
- Utani, K.; Fu, H.; Jang, S.M.; Marks, A.B.; Smith, O.K.; Zhang, Y.; Redon, C.E.; Shimizu, N.; Aladjem, M.I. Phosphorylated SIRT1 associates with replication origins to prevent excess replication initiation and preserve genomic stability. Nucleic Acids Res. 2017, 45, 7807–7824. [Google Scholar] [CrossRef] [PubMed]
- Alves-Fernandes, D.K.; Jasiulionis, M.G. The Role of SIRT1 on DNA Damage Response and Epigenetic Alterations in Cancer. Int. J. Mol. Sci. 2019, 20, 3153. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.H.; Cao, L.; Mostoslavsky, R.; Lombard, D.B.; Liu, J.; Bruns, N.E.; Tsokos, M.; Alt, F.W.; Finkel, T. A role for the NAD-dependent deacetylase Sirt1 in the regulation of autophagy. Proc. Natl. Acad. Sci. USA 2008, 105, 3374–3379. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.L. Sirt1 and the Mitochondria. Mol. Cells 2016, 39, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Sarikhani, M.; Maity, S.; Mishra, S.; Jain, A.; Tamta, A.K.; Ravi, V.; Kondapalli, M.S.; Desingu, P.A.; Khan, D.; Kumar, S.; et al. SIRT2 deacetylase represses NFAT transcription factor to maintain cardiac homeostasis. J. Biol. Chem. 2018, 293, 5281–5294. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, J.; Hong, T.; Chen, X.; Cui, L. SIRT2: Controversy and multiple roles in disease and physiology. Ageing Res. Rev. 2019, 55, 100961. [Google Scholar] [CrossRef]
- Liu, G.; Park, S.H.; Imbesi, M.; Nathan, W.J.; Zou, X.; Zhu, Y.; Jiang, H.; Parisiadou, L.; Gius, D. Loss of NAD-Dependent Protein Deacetylase Sirtuin-2 Alters Mitochondrial Protein Acetylation and Dysregulates Mitophagy. Antioxid. Redox Signal. 2017, 26, 849–863. [Google Scholar] [CrossRef]
- Amjad, S.; Nisar, S.; Bhat, A.A.; Shah, A.R.; Frenneaux, M.P.; Fakhro, K.; Haris, M.; Reddy, R.; Patay, Z.; Baur, J.; et al. Role of NAD+ in regulating cellular and metabolic signaling pathways. Mol. Metab. 2021, 49, 101195. [Google Scholar] [CrossRef]
- Wang, C.H.; Wei, Y.H. Roles of Mitochondrial Sirtuins in Mitochondrial Function, Redox Homeostasis, Insulin Resistance and Type 2 Diabetes. Int. J. Mol. Sci. 2020, 21, 5266. [Google Scholar] [CrossRef]
- Michishita, E.; Park, J.Y.; Burneskis, J.M.; Barrett, J.C.; Horikawa, I. Evolutionarily conserved and nonconserved cellular localizations and functions of human SIRT proteins. Mol. Biol. Cell. 2005, 16, 4623–4635. [Google Scholar] [CrossRef] [Green Version]
- Mendes, K.L.; Lelis, D.F.; Santos, S.H.S. Nuclear sirtuins and inflammatory signaling pathways. Cytokine Growth Factor Rev. 2017, 38, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Leite, J.A.; Ghirotto, B.; Targhetta, V.P.; de Lima, J.; Camara, N.O.S. Sirtuins as pharmacological targets in neurodegenerative and neuropsychiatric disorders. Br. J. Pharmacol. 2022, 179, 1496–1511. [Google Scholar] [CrossRef] [PubMed]
- Khayatan, D.; Razavi, S.M.; Arab, Z.N.; Khanahmadi, M.; Momtaz, S.; Butler, A.E.; Montecucco, F.; Markina, Y.V.; Abdolghaffari, A.H.; Sahebkar, A. Regulatory Effects of Statins on SIRT1 and Other Sirtuins in Cardiovascular Diseases. Life 2022, 12, 760. [Google Scholar] [CrossRef] [PubMed]
- Zhao, E.; Hou, J.; Ke, X.; Abbas, M.N.; Kausar, S.; Zhang, L.; Cui, H. The Roles of Sirtuin Family Proteins in Cancer Progression. Cancers 2019, 11, 1949. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.Y.; Lin, H. Sirtuin Modulators in Cellular and Animal Models of Human Diseases. Front. Pharmacol. 2021, 12, 735044. [Google Scholar] [CrossRef]
- Kim, S.Y.; Yang, C.S.; Lee, H.M.; Kim, J.K.; Kim, Y.S.; Kim, Y.R.; Kim, J.S.; Kim, T.S.; Yuk, J.M.; Dufour, C.R.; et al. ESRRA (estrogen-related receptor alpha) is a key coordinator of transcriptional and post-translational activation of autophagy to promote innate host defense. Autophagy 2018, 14, 152–168. [Google Scholar] [CrossRef]
- Ou, X.; Lee, M.R.; Huang, X.; Messina-Graham, S.; Broxmeyer, H.E. SIRT1 positively regulates autophagy and mitochondria function in embryonic stem cells under oxidative stress. Stem Cells 2014, 32, 1183–1194. [Google Scholar] [CrossRef]
- Kim, S.R.; Lee, K.S.; Park, S.J.; Min, K.H.; Choe, Y.H.; Moon, H.; Yoo, W.H.; Chae, H.J.; Han, M.K.; Lee, Y.C. Involvement of sirtuin 1 in airway inflammation and hyperresponsiveness of allergic airway disease. J. Allergy Clin. Immunol. 2010, 125, 449–460 e414. [Google Scholar] [CrossRef]
- Isaacs-Ten, A.; Moreno-Gonzalez, M.; Bone, C.; Martens, A.; Bernuzzi, F.; Ludwig, T.; Hellmich, C.; Hiller, K.; Rushworth, S.A.; Beraza, N. Metabolic Regulation of Macrophages by SIRT1 Determines Activation During Cholestatic Liver Disease in Mice. Cell. Mol. Gastroenterol. Hepatol. 2022, 13, 1019–1039. [Google Scholar] [CrossRef]
- Vachharajani, V.T.; Liu, T.; Wang, X.; Hoth, J.J.; Yoza, B.K.; McCall, C.E. Sirtuins Link Inflammation and Metabolism. J. Immunol. Res. 2016, 2016, 8167273. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.F.; Vachharajani, V.T.; Yoza, B.K.; McCall, C.E. NAD+-dependent sirtuin 1 and 6 proteins coordinate a switch from glucose to fatty acid oxidation during the acute inflammatory response. J. Biol. Chem. 2012, 287, 25758–25769. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.F.; Yoza, B.K.; El Gazzar, M.; Vachharajani, V.T.; McCall, C.E. NAD+-dependent SIRT1 deacetylase participates in epigenetic reprogramming during endotoxin tolerance. J. Biol. Chem 2011, 286, 9856–9864. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhao, C.; Kong, P.; Sun, H.; Sun, Z.; Bian, G.; Sun, Y.; Guo, L. Treatment with NAD+ inhibited experimental autoimmune encephalomyelitis by activating AMPK/SIRT1 signaling pathway and modulating Th1/Th17 immune responses in mice. Int. Immunopharmacol. 2016, 39, 287–294. [Google Scholar] [CrossRef]
- Liu, G.; Bi, Y.; Xue, L.; Zhang, Y.; Yang, H.; Chen, X.; Lu, Y.; Zhang, Z.; Liu, H.; Wang, X.; et al. Dendritic cell SIRT1-HIF1alpha axis programs the differentiation of CD4+ T cells through IL-12 and TGF-beta1. Proc. Natl. Acad. Sci. USA 2015, 112, E957–E965. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kundu, M.; Viollet, B.; Guan, K.L. AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat. Cell Biol. 2011, 13, 132–141. [Google Scholar] [CrossRef]
- Wu, Y.; Li, X.; Zhu, J.X.; Xie, W.; Le, W.; Fan, Z.; Jankovic, J.; Pan, T. Resveratrol-activated AMPK/SIRT1/autophagy in cellular models of Parkinson’s disease. Neurosignals 2011, 19, 163–174. [Google Scholar] [CrossRef]
- Luo, G.; Jian, Z.; Zhu, Y.; Zhu, Y.; Chen, B.; Ma, R.; Tang, F.; Xiao, Y. Sirt1 promotes autophagy and inhibits apoptosis to protect cardiomyocytes from hypoxic stress. Int. J. Mol. Med. 2019, 43, 2033–2043. [Google Scholar] [CrossRef]
- Chakraborty, S.; Datta, S.; Ghosh, S. Induction of autophagy under nitrosative stress: A complex regulatory interplay between SIRT1 and AMPK in MCF7 cells. Cell. Signal. 2019, 64, 109411. [Google Scholar] [CrossRef]
- Elesela, S.; Morris, S.B.; Narayanan, S.; Kumar, S.; Lombard, D.B.; Lukacs, N.W. Sirtuin 1 regulates mitochondrial function and immune homeostasis in respiratory syncytial virus infected dendritic cells. PLoS Pathog. 2020, 16, e1008319. [Google Scholar] [CrossRef]
- Kauppinen, A.; Suuronen, T.; Ojala, J.; Kaarniranta, K.; Salminen, A. Antagonistic crosstalk between NF-κB and SIRT1 in the regulation of inflammation and metabolic disorders. Cell. Signal. 2013, 25, 1939–1948. [Google Scholar] [CrossRef]
- Paik, S.; Jo, E.K. An Interplay between Autophagy and Immunometabolism for Host Defense Against Mycobacterial Infection. Front. Immunol. 2020, 11, 603951. [Google Scholar] [CrossRef] [PubMed]
- Riffelmacher, T.; Richter, F.C.; Simon, A.K. Autophagy dictates metabolism and differentiation of inflammatory immune cells. Autophagy 2018, 14, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.C.; Guarente, L. SIRT1 and other sirtuins in metabolism. Trends Endocrinol. Metab. 2014, 25, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Giblin, W.; Skinner, M.E.; Lombard, D.B. Sirtuins: Guardians of mammalian healthspan. Trends Genet. 2014, 30, 271–286. [Google Scholar] [CrossRef] [PubMed]
- Crotty Alexander, L.E.; Marsh, B.J.; Timmer, A.M.; Lin, A.E.; Zainabadi, K.; Czopik, A.; Guarente, L.; Nizet, V. Myeloid cell sirtuin-1 expression does not alter host immune responses to Gram-negative endotoxemia or Gram-positive bacterial infection. PLoS ONE 2013, 8, e84481. [Google Scholar] [CrossRef]
- Ganesan, R.; Hos, N.J.; Gutierrez, S.; Fischer, J.; Stepek, J.M.; Daglidu, E.; Kronke, M.; Robinson, N. Salmonella Typhimurium disrupts Sirt1/AMPK checkpoint control of mTOR to impair autophagy. PLoS Pathog. 2017, 13, e1006227. [Google Scholar] [CrossRef]
- Xu, W.; Tian, K.; Li, X.; Zhang, S. IL-9 blockade attenuates inflammation in a murine model of methicillin-resistant Staphylococcus aureus pneumonia. Acta. Biochim. Biophys. Sin. 2020, 52, 133–140. [Google Scholar] [CrossRef]
- Tian, K.; Xu, W. MiR-155 regulates Th9 differentiation in children with methicillin-resistant Staphylococcus aureus pneumonia by targeting SIRT1. Hum. Immunol. 2021, 82, 775–781. [Google Scholar] [CrossRef]
- Cheng, C.Y.; Gutierrez, N.M.; Marzuki, M.B.; Lu, X.; Foreman, T.W.; Paleja, B.; Lee, B.; Balachander, A.; Chen, J.; Tsenova, L.; et al. Host sirtuin 1 regulates mycobacterial immunopathogenesis and represents a therapeutic target against tuberculosis. Sci. Immunol. 2017, 2, 9. [Google Scholar] [CrossRef]
- Kwon, H.S.; Brent, M.M.; Getachew, R.; Jayakumar, P.; Chen, L.F.; Schnolzer, M.; McBurney, M.W.; Marmorstein, R.; Greene, W.C.; Ott, M. Human immunodeficiency virus type 1 Tat protein inhibits the SIRT1 deacetylase and induces T cell hyperactivation. Cell Host Microbe 2008, 3, 158–167. [Google Scholar] [CrossRef] [Green Version]
- Gruevska, A.; Moragrega, A.B.; Galindo, M.J.; Esplugues, J.V.; Blas-Garcia, A.; Apostolova, N. Down-Regulation of the Longevity-Associated Protein SIRT1 in Peripheral Blood Mononuclear Cells of Treated HIV Patients. Cells 2022, 11, 348. [Google Scholar] [CrossRef]
- Bordoni, V.; Tartaglia, E.; Refolo, G.; Sacchi, A.; Grassi, G.; Antinori, A.; Fimia, G.M.; Agrati, C. Per2 Upregulation in Circulating Hematopoietic Progenitor Cells During Chronic HIV Infection. Front. Cell. Infect. Microbiol. 2020, 10, 362. [Google Scholar] [CrossRef] [PubMed]
- Jurkowska, K.; Szymanska, B.; Knysz, B.; Piwowar, A. The Effect of Antiretroviral Therapy on SIRT1, SIRT3 and SIRT6 Expression in HIV-Infected Patients. Molecules 2022, 27, 1358. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.A.; Seong, R.K.; Shin, O.S. Enhanced Viral Replication by Cellular Replicative Senescence. Immune Netw. 2016, 16, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, G.Y.; Ren, J.P.; Wang, L.; Zhao, J.; Ning, S.B.; Zhang, Y.; Lian, J.Q.; Huang, C.X.; Jia, Z.S.; et al. Protection of CD4+ T cells from hepatitis C virus infection-associated senescence via DeltaNp63-miR-181a-Sirt1 pathway. J. Leukoc. Biol. 2016, 100, 1201–1211. [Google Scholar] [CrossRef]
- Kim, C.; Jin, J.; Ye, Z.; Jadhav, R.R.; Gustafson, C.E.; Hu, B.; Cao, W.; Tian, L.; Weyand, C.M.; Goronzy, J.J. Histone deficiency and accelerated replication stress in T cell aging. J. Clin. Investig. 2021, 131, e143632. [Google Scholar] [CrossRef]
- Owczarczyk, A.B.; Schaller, M.A.; Reed, M.; Rasky, A.J.; Lombard, D.B.; Lukacs, N.W. Sirtuin 1 Regulates Dendritic Cell Activation and Autophagy during Respiratory Syncytial Virus-Induced Immune Responses. J. Immunol. 2015, 195, 1637–1646. [Google Scholar] [CrossRef]
- Bordoni, V.; Tartaglia, E.; Sacchi, A.; Fimia, G.M.; Cimini, E.; Casetti, R.; Notari, S.; Grassi, G.; Marchioni, L.; Bibas, M.; et al. The unbalanced p53/SIRT1 axis may impact lymphocyte homeostasis in COVID-19 patients. Int. J. Infect. Dis. 2021, 105, 49–53. [Google Scholar] [CrossRef]
- Karkischenko, V.N.; Skvortsova, V.I.; Gasanov, M.T.; Fokin, Y.V.; Nesterov, M.S.; Petrova, N.V.; Alimkina, O.V.; Pomytkin, I.A. Inhaled [D-Ala(2)]-Dynorphin 1-6 Prevents Hyperacetylation and Release of High Mobility Group Box 1 in a Mouse Model of Acute Lung Injury. J. Immunol. Res. 2021, 2021, 4414544. [Google Scholar] [CrossRef]
- Wan, X.; Garg, N.J. Sirtuin Control of Mitochondrial Dysfunction, Oxidative Stress, and Inflammation in Chagas Disease Models. Front. Cell. Infect. Microbiol. 2021, 11, 693051. [Google Scholar] [CrossRef]
- Wan, X.; Wen, J.J.; Koo, S.J.; Liang, L.Y.; Garg, N.J. SIRT1-PGC1α-NFκB Pathway of Oxidative and Inflammatory Stress during Trypanosoma cruzi Infection: Benefits of SIRT1-Targeted Therapy in Improving Heart Function in Chagas Disease. PLoS Pathog. 2016, 12, e1005954. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.; Chowdhury, I.H.; Jie, Z.; Choudhuri, S.; Garg, N.J. Origin of Monocytes/Macrophages Contributing to Chronic Inflammation in Chagas Disease: SIRT1 Inhibition of FAK-NFκB-Dependent Proliferation and Proinflammatory Activation of Macrophages. Cells 2019, 9, 80. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Munoz, D.; Sanchez, A.; Perez-Benavente, S.; Contreras-Jurado, C.; Montero-Pedrazuela, A.; Toledo-Castillo, M.; Gutierrez-Hernandez, M.; Rodrigues-Diez, R.; Folgueira, C.; Briones, A.M.; et al. Hypothyroidism confers tolerance to cerebral malaria. Sci. Adv. 2022, 8, eabj7110. [Google Scholar] [CrossRef] [PubMed]
- Moreira, D.; Rodrigues, V.; Abengozar, M.; Rivas, L.; Rial, E.; Laforge, M.; Li, X.; Foretz, M.; Viollet, B.; Estaquier, J.; et al. Leishmania infantum modulates host macrophage mitochondrial metabolism by hijacking the SIRT1-AMPK axis. PLoS Pathog. 2015, 11, e1004684. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Saha, S.; Gupta, P.; Ukil, A.; Das, P.K. Crosstalk of PD-1 signaling with the SIRT1/FOXO-1 axis during the progression of visceral leishmaniasis. J. Cell. Sci. 2019, 132, jcs226274. [Google Scholar] [CrossRef]
- Xie, H.; Lei, N.; Gong, A.Y.; Chen, X.M.; Hu, G. Cryptosporidium parvum induces SIRT1 expression in host epithelial cells through downregulating let-7i. Hum. Immunol. 2014, 75, 760–765. [Google Scholar] [CrossRef] [Green Version]


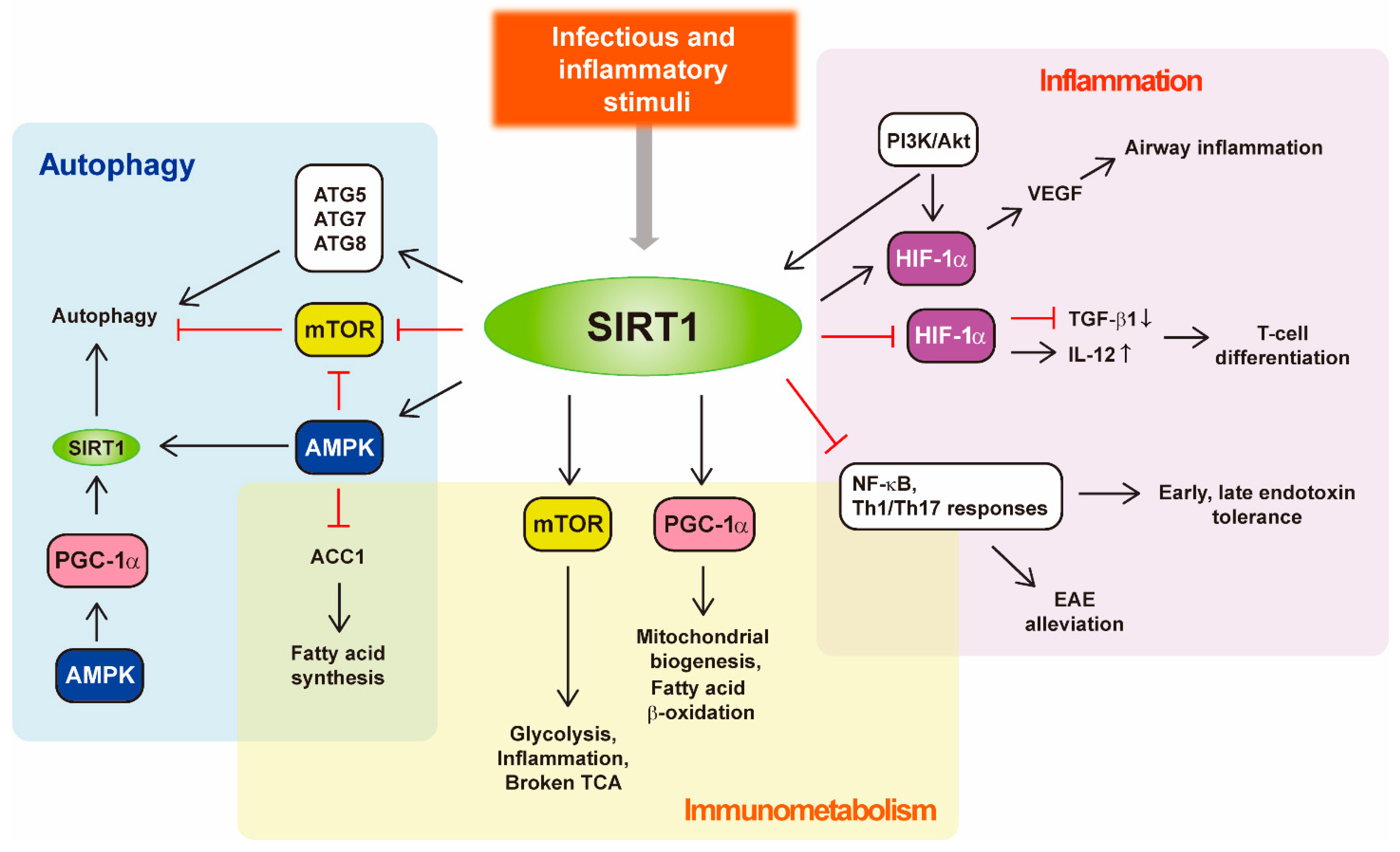{kind=link}
| Pathogen | Cells/Study Model | SIRT1 after Infection | Intervention | Outcome | Mechanism of Action | Ref. |
|---|---|---|---|---|---|---|
| Bacterial Infection | ||||||
| Helicobacter pylori | Human gastric cell lines, gastric tissues | ↓ | RUNX3 knockdown or overexpression, SRT1720 | Increased intracellular survival and colonization | Inhibition of SIRT1 in a RUNX3-dependent manner, and inhibition of autophagy flux | [13] |
| Salmonella Typhimurium | Murine BMDMs | ↓ | Sirt1 overexpression, Torin 1, Atg7−/− | Compromised autophagic host cell defense | AKT-mTOR-dependent degradation of SIRT1 and inhibition of AMPK activation and autophagy by virulence factor SPI2 | [66] |
| Streptococcus pneumoniae | A549 cell line | ↑ | Resveratrol, Nicotinamide | Induction of hBD2 and IL-8 | SIRT1-mediated induction of hBD2 through p38 MAPK and IL-8 through ERK | [11] |
| Staphylococcus aureus | BALF from children with MRSA, CD4+ T cells | ↓ * | miR-155 mimic and inhibitor, SIRT1 overexpression | Th9 differentiation | miR-155-mediated negative regulation of SIRT1 | [68] |
| Mycobacterium tuberculosis | THP-1 cells, hMDMs, in vitro mouse model | ↓ | SRT1720, SIRT1 KO | Higher TB pathogenesis | SIRT1 activation-mediated induction of autophagy and phagosome-lysosome fusion, and deacetylation of RelA/p65 | [69] |
| HEK293T cells, human PBMCs, Mouse PMs | ↓ | TLR2−/−, Sirt1−/−, Resveratrol | Negative regulation of inflammatory responses | Enhanced TAK1 phosphorylation/ubiquitination leading to activation of p65/MAPK signaling pathway | [14] | |
| BMDMs, RAW264.7 cells, HEK293T cells | Not reported | AICAR, Resveratrol, XCT790, esrra−/− | Antibacterial autophagy | AMPK or SIRT1 activation leading to induction of ESRRA | [46] | |
| Viral Infection | ||||||
| Human immunodeficiency virus (HIV) | Jurkat T cells, HeLa, HEK293, MEF cells | ↓ | SIRT1−/−, Tat overexpression, Nicotinamide | Immune cell hyperactivation | Interaction of Tat with SIRT1 to block its deacetylase activity and superinduction of T cell activation and HIV transcription via NF-kB | [70] |
| PBMCs from HIV+ and healthy individuals | ↓ | Resveratrol | Impaired immune reconstitution and accelerated aging process | Negative regulation of Per2 by Sirt1 in HPCs | [72] | |
| Hepatitis C virus (HCV) | PBMCs from healthy and HCV infected patients | ↑ | SIRT1 and ΔNp63 knockdown, miR-181a precursor | T cell senescence and viral persistence | Upregulation of ΔNp63 by HCV leading to inhibition of miR-181a, thereby increasing Sirt1 and DUSP6 in CD4+ T cells | [75] |
| Lymphocytic choriomeningitis virus (LCMV) | Human primary T cells | Not reported | EX-527, SIRT1 knockdown, miR-181a−/− | Diminished cell cycle progression and replication stress | Reduction of histone gene by miR-181a-dependent increase of SIRT1 | [76] |
| Respiratory syncytial virus (RSV) | mouse BMDCs, AECs, in vivo mouse model | ↑ | EX-527, Sirt1 KO, SIRT1 knockdown, SRT1720 | Antiviral immune response | Induction of autophagy, induction of Th1 immune response, suppression of Th2 and Th17 responses | [77] |
| BMDCs, in vivo mouse model | Not reported | SIRT−/−, C75 | Altered immune homeostasis | In SIRT1-deficient DCs—reduction of mitochondrial function/respiration andincreased fatty acid synthesis through acetyl Co A pathway | [59] | |
| COVID-19 | PBMCs from COVID-19 patients | ↓ | - | Altered lymphocyte homeostasis | Increased p53 and reduced SIRT1, reduced expression of IL17R and BLNK | [78] |
| Parasitic Infection | ||||||
| Trypanosoma cruzi | Chagas mice, RAW264.7 cells | Not reported | SRT1720, PF-562271 (iFAK) | Inhibition of inflammation during Chagas disease | Suppression of FAK phosphorylation, FAK-dependent increase of Pu.1 and Runx1, and NF-κB activity | [82] |
| Plasmodium berghei ANKA (PbA) | ECM mice | ↓ | SRT1720, EX-527, Hypothyroid mice | Improved disease outcome | Sirt1 as a mediator of the action of the thyroid hormones | [83] |
| Leishmania infantum | BMDMs, PMs, in vivo mouse model | ↓ | SIRT1 KO, LKB1 KO, SRT1720 | Parasitic growth | Macrophages switch from early glycolytic metabolism to oxidative phosphorylation through SIRT1 and LKB1/AMPK | [84] |
| Leishmania donovani | RAW264.7 cells, In vivo mouse model | ↑ | Sirtinol, AS1842856, SIRT1 knockdown | Parasitic survival | Deacetylation and inactivation of FOXO-1 by SIRT1 to prevent apoptosis | [85] |
| Cryptosporidium parvum | H69 cells | ↑ | Pre-let-7i, Anti-let-7i | - | Reduction in let-7i leading to inhibition of miRNA-mediated translational suppression of SIRT1 to induce its protein level | [86] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.K.; Silwal, P.; Jo, E.-K. Sirtuin 1 in Host Defense during Infection. Cells 2022, 11, 2921. https://doi.org/10.3390/cells11182921
Kim JK, Silwal P, Jo E-K. Sirtuin 1 in Host Defense during Infection. Cells. 2022; 11(18):2921. https://doi.org/10.3390/cells11182921
Chicago/Turabian StyleKim, Jin Kyung, Prashanta Silwal, and Eun-Kyeong Jo. 2022. "Sirtuin 1 in Host Defense during Infection" Cells 11, no. 18: 2921. https://doi.org/10.3390/cells11182921