
Dysregulated Neutrophil Phenotype and Function in Hospitalised Non-ICU COVID-19 Pneumonia

 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Methods
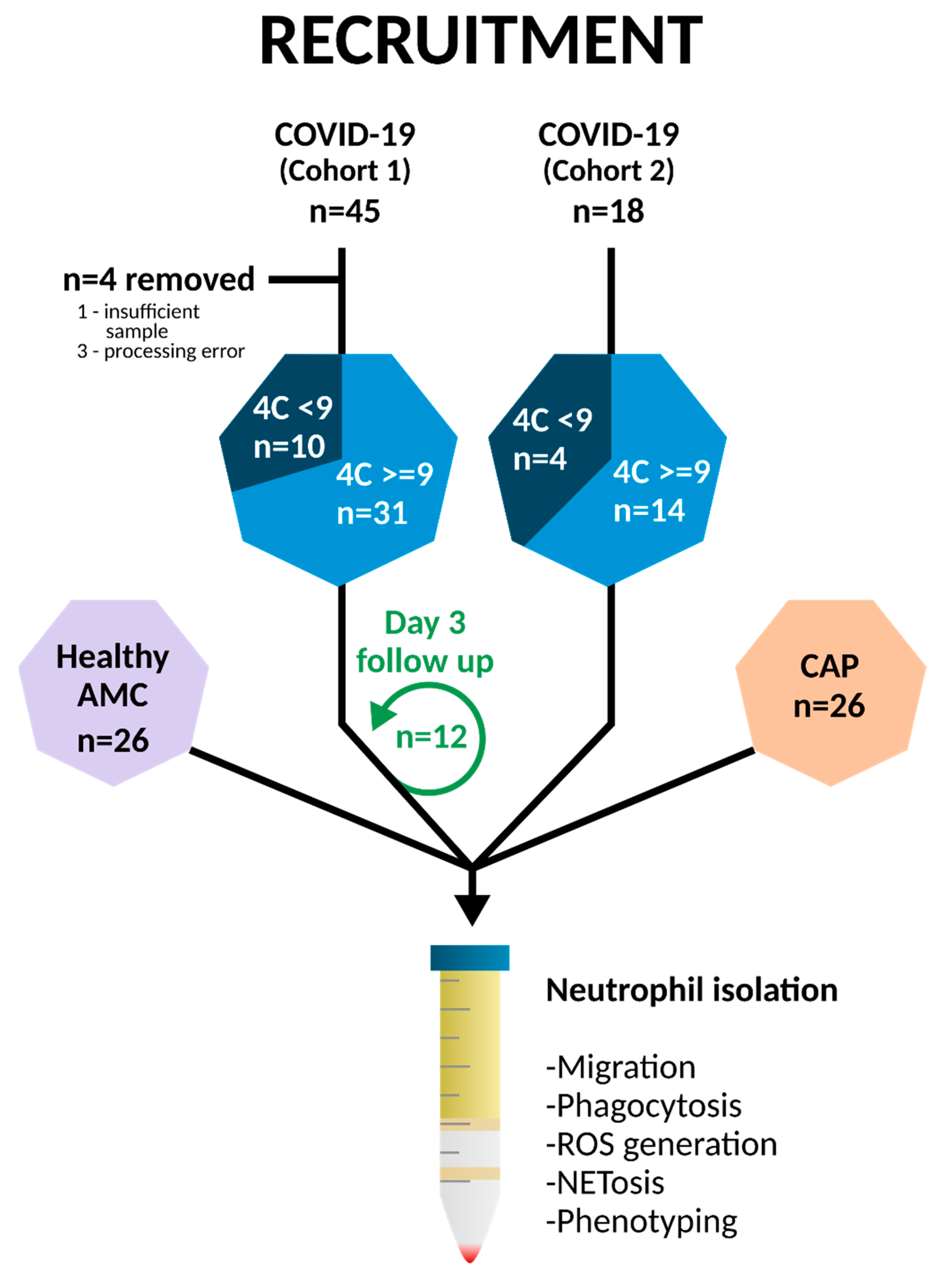
2.1. Healthy Donor and Patient Recruitment
2.2. Neutrophil Phenotypic and Functional Analysis
2.3. Statistics
3. Results
3.1. Clinical Characterisation
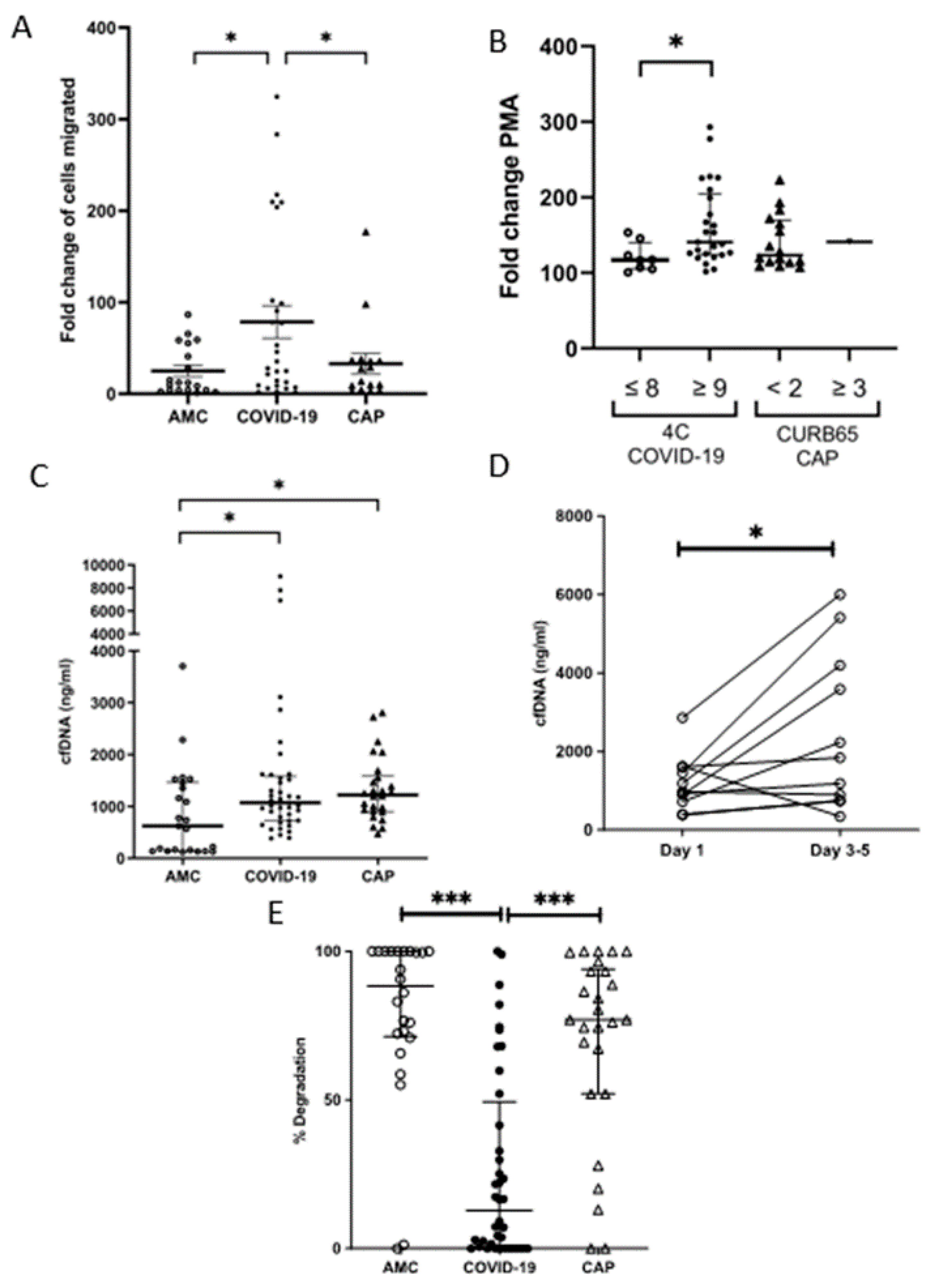
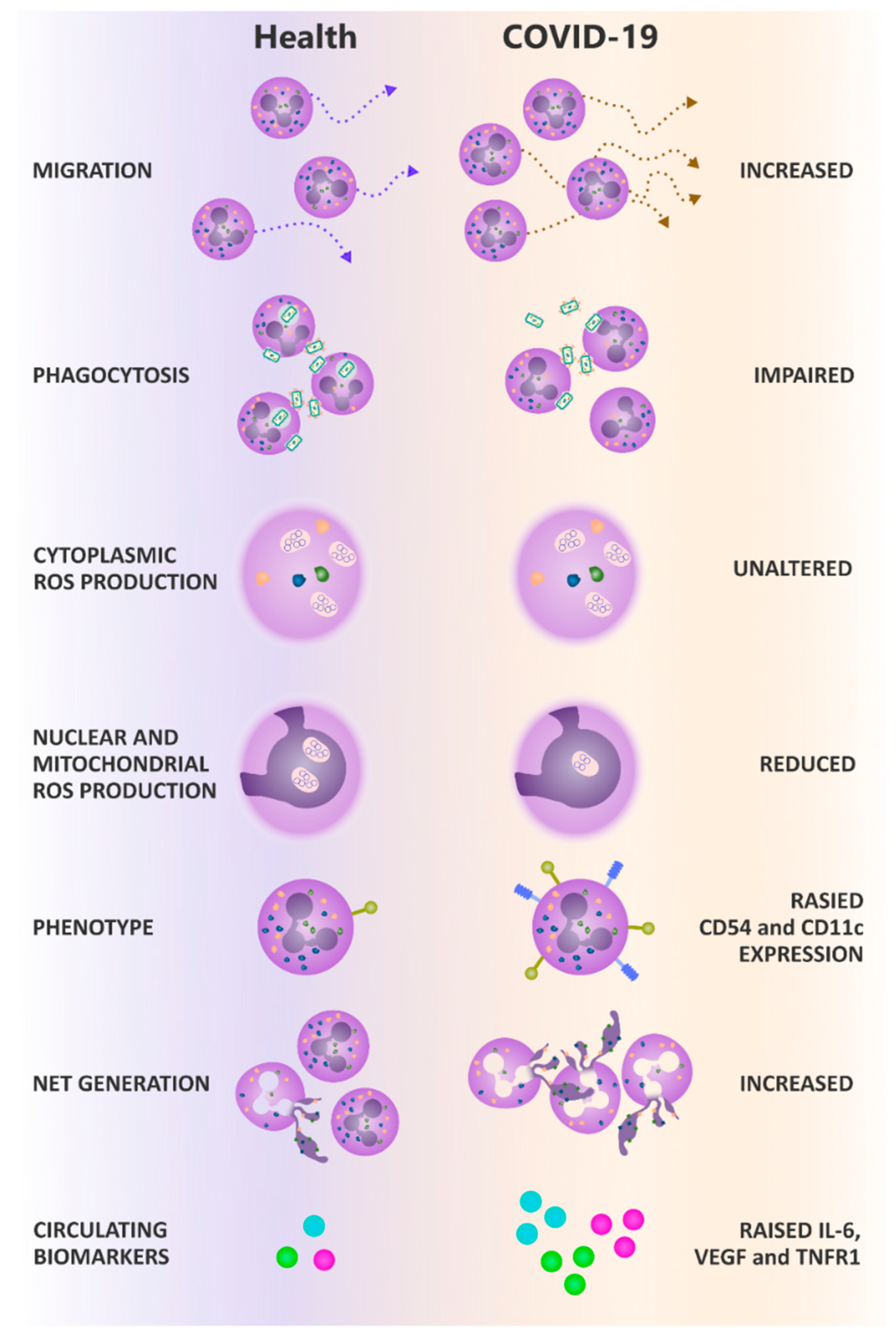
3.2. Neutrophil Migration Is Elevated through a Transwell System in COVID-19
3.3. Neutrophil Phagocytosis Is Impaired in COVID-19
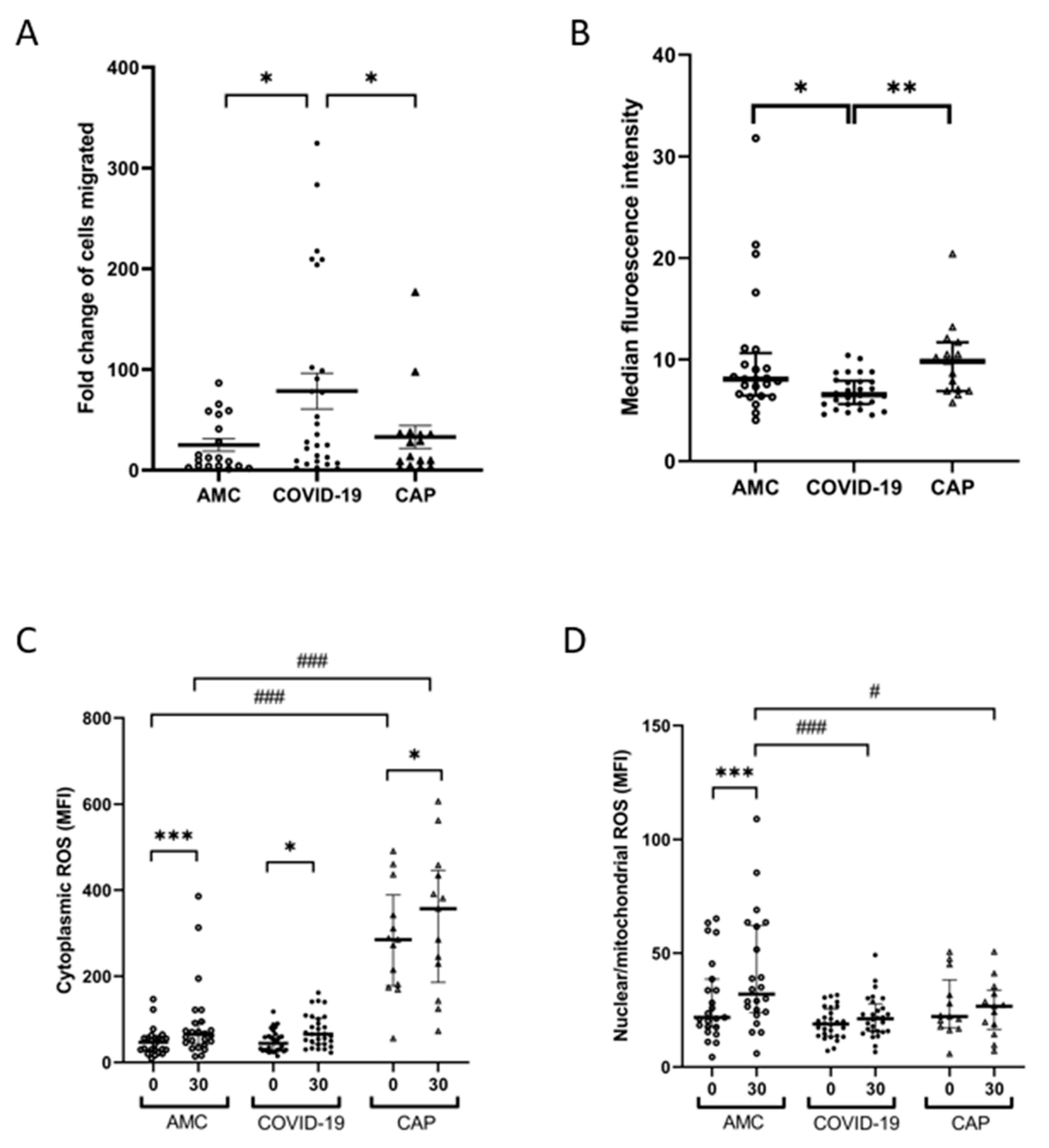
3.4. Neutrophil Derived ROS Generation following Phagocytosis Is Impaired in COVID-19
3.5. Neutrophil NETosis Is Elevated in COVID-19
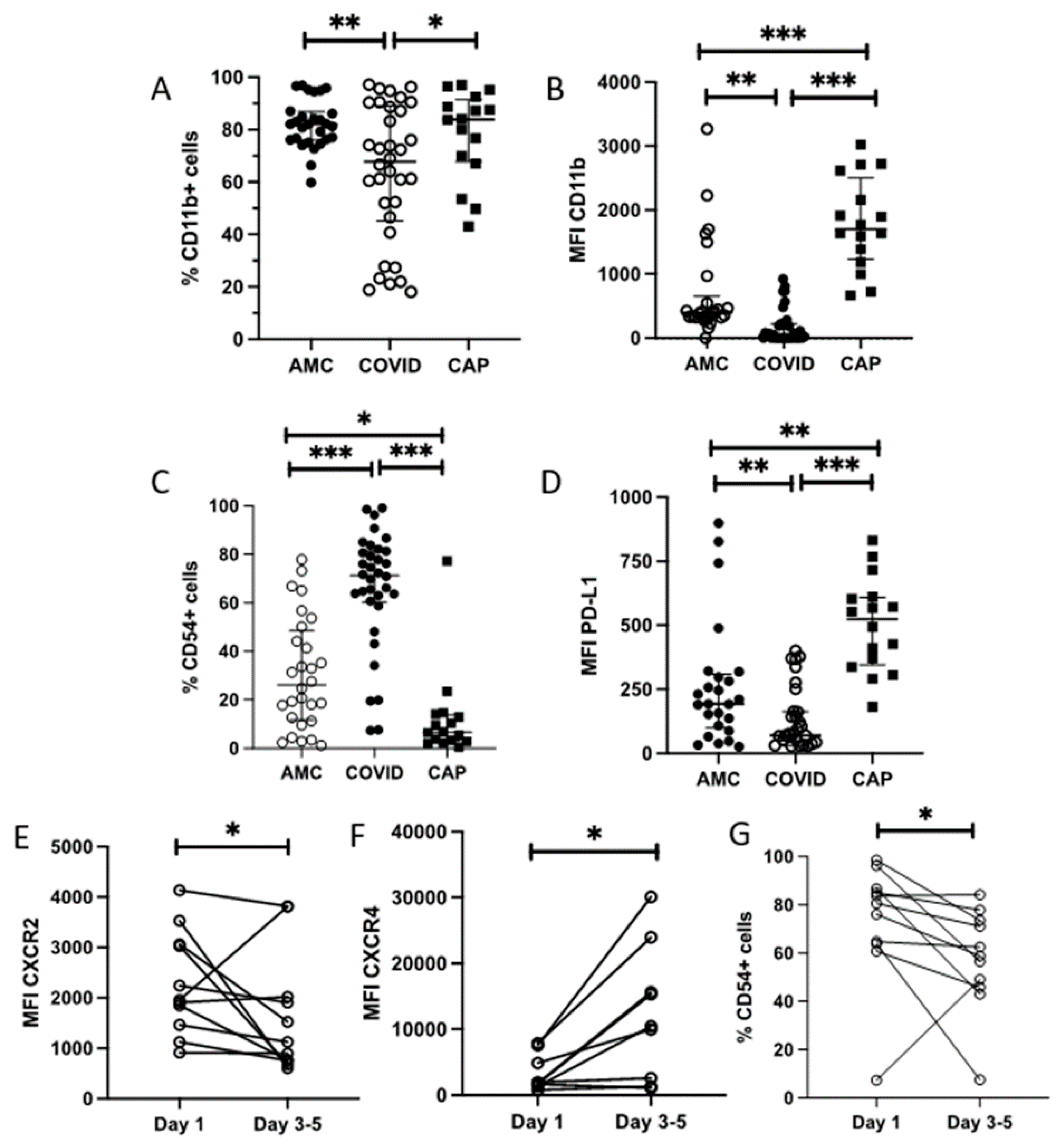
3.6. Neutrophil Phenotype Is Altered in COVID-19
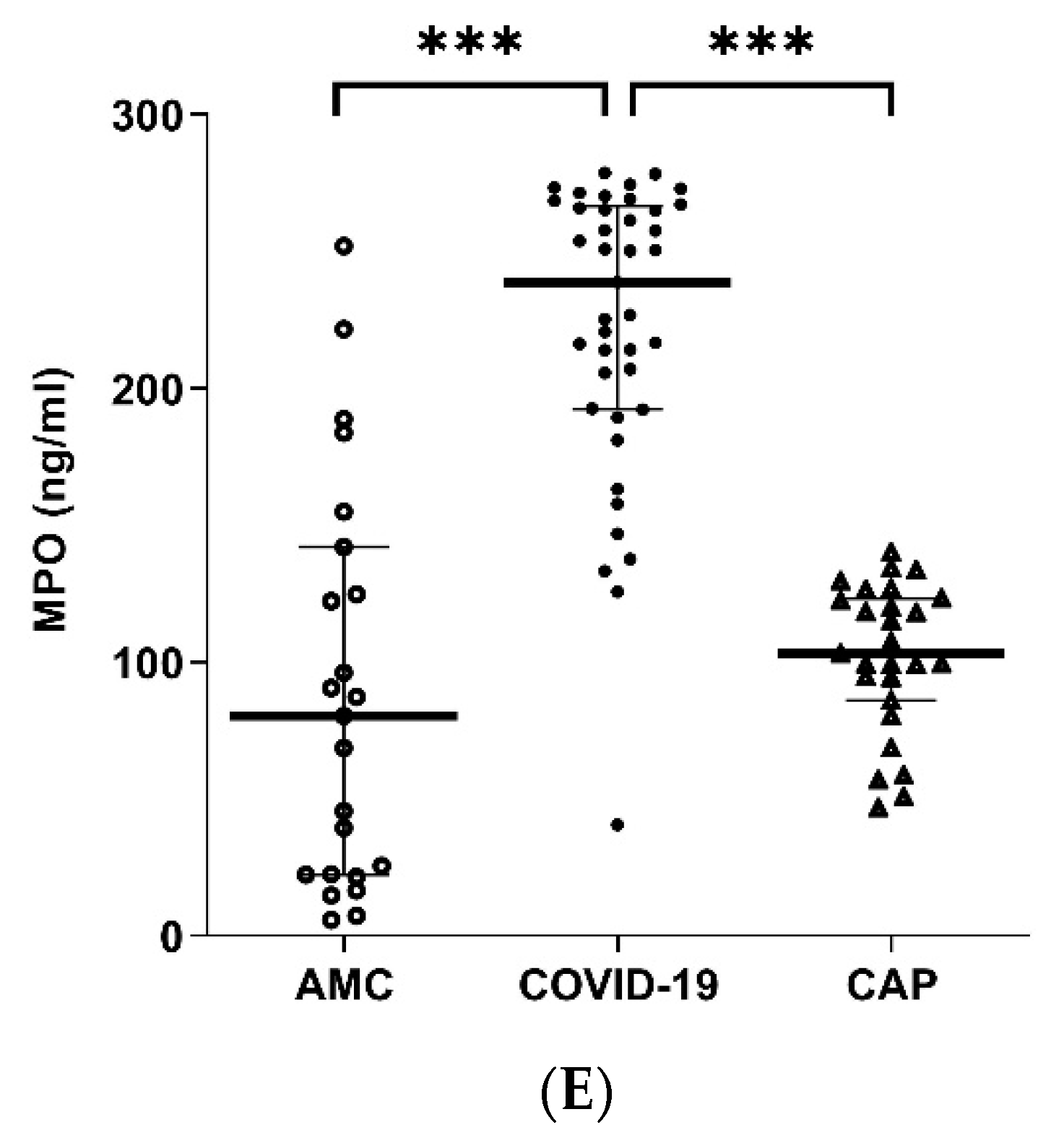
3.7. Systemic Inflammatory Mediators Are Elevated in COVID-19
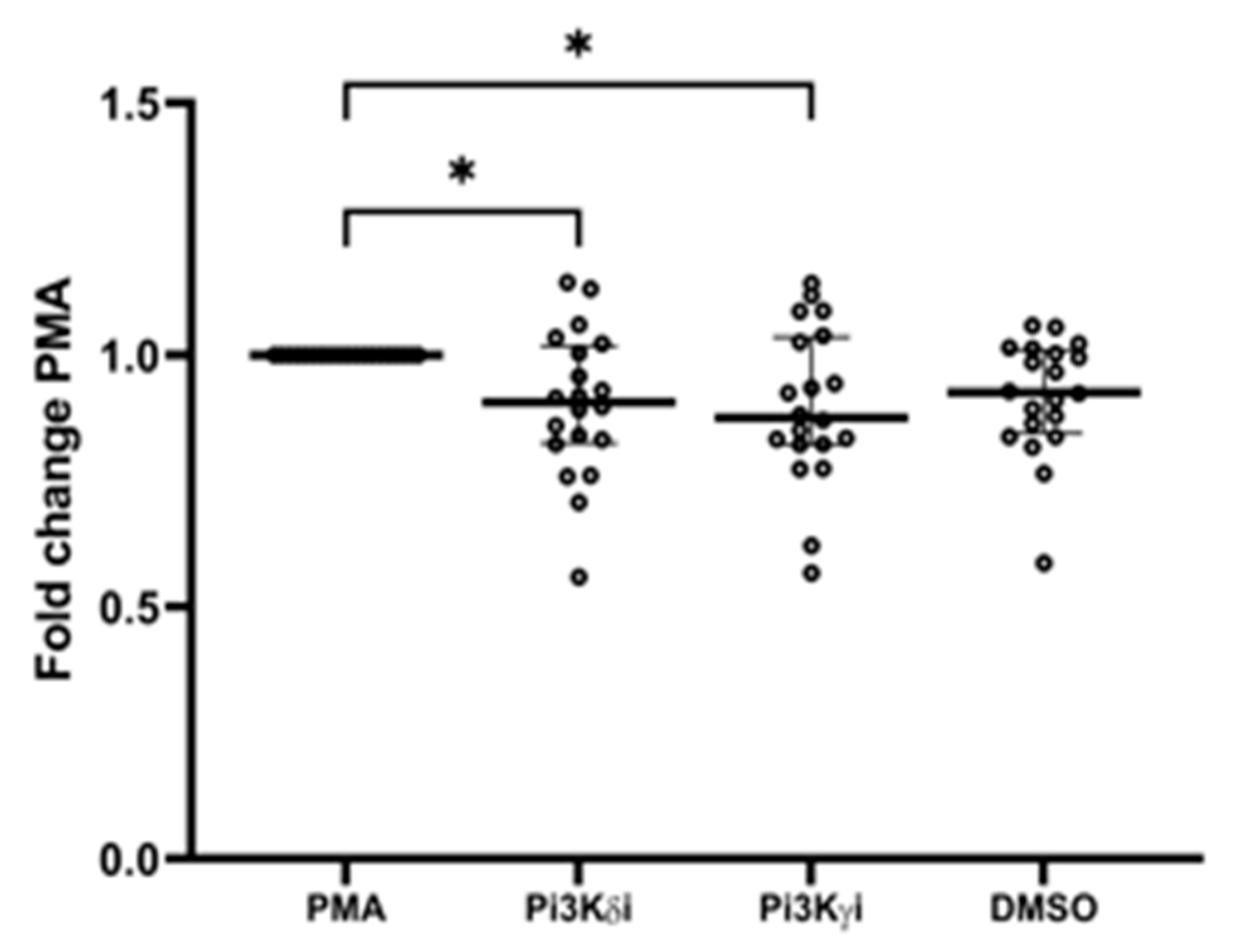
3.8. Pi3K Inhibitors
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cucinotta, D.; Vanelli, M. WHO Declares COVID-19 a Pandemic. Acta Biomed. 2020, 91, 157–160. [Google Scholar]
- Kolb, M.; Margetts, P.J.; Anthony, D.C.; Pitossi, F.; Gauldie, J. Transient expression of IL-1beta induces acute lung injury and chronic repair leading to pulmonary fibrosis. J. Clin. Invest. 2001, 107, 1529–1536. [Google Scholar] [CrossRef] [PubMed]
- Dorward, D.A.; Russell, C.D.; Um, I.H.; Elshani, M.; Armstrong, S.D.; Penrice-Randal, R.; Millar, T.; Lerpiniere, C.E.B.; Tagliavini, G.; Hartley, C.S.; et al. Tissue-Specific Immunopathology in Fatal COVID-19. Am. J. Respir. Crit. Care Med. 2021, 203, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Galani, I.E.; Andreakos, E. Neutrophils in viral infections: Current concepts and caveats. J. Leukoc. Biol. 2015, 98, 557–564. [Google Scholar] [CrossRef]
- Aulakh, G.K. Neutrophils in the lung: “The first responders”. Cell Tissue Res. 2017, 371, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Hu, C.; Luo, L.; Fang, F.; Chen, Y.; Li, J.; Peng, Z.; Pan, H. Clinical features and short-term outcomes of 221 patients with COVID-19 in Wuhan, China. J. Clin. Virol. 2020, 127, 104364. [Google Scholar] [CrossRef]
- Printz, C. Poor COVID-19 outcomes and deaths linked to advanced age and pre-existing conditions. Cancer 2021, 127, 497. [Google Scholar] [CrossRef]
- Chen, Y.; Klein, S.L.; Garibaldi, B.T.; Li, H.; Wu, C.; Osevala, N.M.; Li, T.; Margolick, J.B.; Pawelec, G.; Leng, S.X. Aging in COVID-19: Vulnerability, immunity and intervention. Ageing Res. Rev. 2021, 65, 101205. [Google Scholar] [CrossRef]
- Sapey, E.; Greenwood, H.; Walton, G.; Mann, E.; Love, A.; Aaronson, N.; Insall, R.H.; Stockley, R.A.; Lord, J.M. Phosphoinositide 3-kinase inhibition restores neutrophil accuracy in the elderly: Toward targeted treatments for immunosenescence. Blood 2014, 123, 239–248. [Google Scholar] [CrossRef]
- Butcher, S.K.; Chahal, H.; Nayak, L.; Sinclair, A.; Henriquez, N.V.; Sapey, E.; O’Mahony, D.; Lord, J.M. Senescence in innate immune responses: Reduced neutrophil phagocytic capacity and CD16 expression in elderly humans. J. Leukoc. Biol. 2001, 70, 881–886. [Google Scholar]
- Hazeldine, J.; Harris, P.; Chapple, I.L.; Grant, M.; Greenwood, H.; Livesey, A.; Sapey, E.; Lord, J.M. Impaired neutrophil extracellular trap formation: A novel defect in the innate immune system of aged individuals. Aging Cell 2014, 13, 690–698. [Google Scholar] [CrossRef] [PubMed]
- Grudzinska, F.S.; Brodlie, M.; Scholefield, B.; Jackson, T.; Scott, A.; Thickett, D.; Sapey, E. Neutrophils in community-acquired pneumonia: Parallels in dysfunction at the extremes of age. Thorax 2020, 75, 164–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, J.M.; Sapey, E.; Parekh, D.; Scott, A.; Dosanjh, D.; Gao, F.; Thickett, D. Sepsis Induces a Dysregulated Neutrophil Phenotype That Is Associated with Increased Mortality. Mediat. Inflamm. 2018, 2018, 4065362. [Google Scholar] [CrossRef] [PubMed]
- Juss, J.K.; House, D.; Amour, A.; Begg, M.; Herre, J.; Storisteanu, D.M.L.; Hoenderdos, K.; Bradley, G.; Lennon, M.; Summers, C.; et al. Acute Respiratory Distress Syndrome Neutrophils Have a Distinct Phenotype and Are Resistant to Phosphoinositide 3-Kinase Inhibition. Am. J. Respir. Crit. Care Med. 2016, 194, 961–973. [Google Scholar] [CrossRef] [PubMed]
- Sapey, E.; Patel, J.M.; Greenwood, H.L.; Walton, G.M.; Hazeldine, J.; Sadhra, C.; Parekh, D.; Dancer, R.C.; Nightingale, P.; Lord, J.M.; et al. Pulmonary infections in the elderly lead to impaired neutrophil targetting, which is improved by simvastatin. Am. J. Respir. Crit. Care Med. 2017, 196, 1325–1336. [Google Scholar] [CrossRef]
- Greenwood, J.; Steinman, L.; Zamvil, S.S. Statin therapy and autoimmune disease: From protein prenylation to immunomodulation. Nat. Rev. Immunol. 2006, 6, 358–370. [Google Scholar] [CrossRef] [PubMed]
- Sapey, E.; Patel, J.; Greenwood, H.; Walton, G.; Grudzinska, F.; Parekh, D.; Mahida, R.Y.; Dancer, R.C.A.; Lugg, S.T.; Howells, P.A.; et al. Simvastatin Improves Neutrophil Function and Clinical Outcomes in Pneumonia. A pilot randomized controlled clinical trial. Am. J. Respir. Crit. Care Med. 2019, 200, 1282–1293. [Google Scholar] [CrossRef]
- Drew, W.; Wilson, D.V.; Sapey, E. Inflammation and neutrophil immunosenescence in health and disease: Targeted treatments to improve clinical outcomes in the elderly. Exp. Gerontol. 2018, 105, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Rosales, C. Neutrophil: A Cell with Many Roles in Inflammation or Several Cell Types? Front. Physiol. 2018, 9, 113. [Google Scholar] [CrossRef]
- Wang, J.; Li, Q.; Yin, Y.; Zhang, Y.; Cao, Y.; Lin, X.; Huang, L.; Hoffmann, D.; Lu, M.; Qiu, Y. Excessive Neutrophils and Neutrophil Extracellular Traps in COVID-19. Front. Immunol. 2020, 11, 2063. [Google Scholar] [CrossRef]
- Aschenbrenner, A.C.; Mouktaroudi, M.; Krämer, B.; Oestreich, M.; Antonakos, N.; Nuesch-Germano, M.; Gkizeli, K.; Bonaguro, L.; Reusch, N.; Baßler, K.; et al. Disease severity-specific neutrophil signatures in blood transcriptomes stratify COVID-19 patients. Genome Med. 2021, 13, 7. [Google Scholar] [CrossRef] [PubMed]
- Schulte-Schrepping, J.; Reusch, N.; Paclik, D.; Baßler, K.; Schlickeiser, S.; Zhang, B.; Krämer, B.; Krammer, T.; Brumhard, S.; Bonaguro, L.; et al. Severe COVID-19 Is Marked by a Dysregulated Myeloid Cell Compartment. Cell 2020, 182, 1419–1440.e23. [Google Scholar] [CrossRef] [PubMed]
- Middleton, E.A.; He, X.-Y.; Denorme, F.; Campbell, R.A.; Ng, D.; Salvatore, S.P.; Mostyka, M.; Baxter-Stoltzfus, A.; Borczuk, A.C.; Loda, M.; et al. Neutrophil extracellular traps contribute to immunothrombosis in COVID-19 acute respiratory distress syndrome. Blood 2020, 136, 1169–1179. [Google Scholar] [CrossRef]
- Skendros, P.; Mitsios, A.; Chrysanthopoulou, A.; Mastellos, D.C.; Metallidis, S.; Rafailidis, P.; Ntinopoulou, M.; Sertaridou, E.; Tsironidou, V.; Tsigalou, C.; et al. Complement and tissue factor-enriched neutrophil extracellular traps are key drivers in COVID-19 immunothrombosis. J. Clin. Invest. 2020, 130, 6151–6157. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Yalavarthi, S.; Shi, H.; Gockman, K.; Zuo, M.; Madison, J.A.; Blair, C.; Weber, A.; Barnes, B.J.; Egeblad, M.; et al. Neutrophil extracellular traps in COVID-19. JCI Insight 2020, 5, e138999. [Google Scholar] [CrossRef] [PubMed]
- Arcanjo, A.; Logullo, J.; Menezes, C.C.B.; Giangiarulo, T.C.D.S.C.; Dos Reis, M.C.; De Castro, G.M.M.; Fontes, Y.D.S.; Todeschini, A.R.; Freire-De-Lima, L.; Decoté-Ricardo, D.; et al. The emerging role of neutrophil extracellular traps in severe acute respiratory syndrome coronavirus 2 (COVID-19). Sci. Rep. 2020, 10, 19630. [Google Scholar] [CrossRef]
- Leppkes, M.; Knopf, J.; Naschberger, E.; Lindemann, A.; Singh, J.; Herrmann, I.; Stürzl, M.; Staats, L.; Mahajan, A.; Schauer, C.; et al. Vascular occlusion by neutrophil extracellular traps in COVID-19. EBioMedicine 2020, 58, 102925. [Google Scholar] [CrossRef]
- Hazeldine, J.; Lord, J.M. Neutrophils and COVID-19: Active Participants and Rational Therapeutic Targets. Front. Immunol. 2021, 12, 680134. [Google Scholar] [CrossRef]
- Sapey, E.; Gallier, S.; Mainey, C.; Nightingale, P.; McNulty, D.; Crothers, H.; Evison, F.; Reeves, K.; Pagano, D.; Denniston, A.K.; et al. Ethnicity and risk of death in patients hospitalised for COVID-19 infection in the UK: An observational cohort study in an urban catchment area. BMJ Open Respir. Res. 2020, 7, e000644. [Google Scholar] [CrossRef]
- Knight, S.R.; Ho, A.; Pius, R.; Buchan, I.; Carson, G.; Drake, T.M.; Dunning, J.; Fairfield, C.J.; Gamble, C.; Green, C.A.; et al. Risk stratification of patients admitted to hospital with COVID-19 using the ISARIC WHO Clinical Characterisation Protocol: Development and validation of the 4C Mortality Score. BMJ 2020, 370, m3339. [Google Scholar] [CrossRef]
- National Institute for Health and Care Excellence. COVID-19 Rapid Guideline: Managing COVID-19; National Institute for Health and Care Excellence: London, UK, 2021.
- Force, A.D.T.; Ranieri, V.M.; Rubenfeld, G.D.; Thompson, B.; Ferguson, N.; Caldwell, E.; Fan, E.; Camporota, L.; Slutsky, A.S. Acute respiratory distress syndrome: The Berlin Definition. JAMA 2012, 307, 2526–2533. [Google Scholar]
- Liu, C.L.; Tangsombatvisit, S.; Rosenberg, J.M.; Mandelbaum, G.; Gillespie, E.C.; Gozani, O.P.; Alizadeh, A.A.; Utz, P.J. Specific post-translational histone modifications of neutrophil extracellular traps as immunogens and potential targets of lupus autoantibodies. Arthritis Res. 2012, 14, R25. [Google Scholar] [CrossRef] [PubMed]
- Sinha, P.; Delucchi, K.L.; McAuley, D.F.; O’Kane, C.M.; Matthay, A.M.; Calfee, C.S. Development and validation of parsimonious algorithms to classify acute respiratory distress syndrome phenotypes: A secondary analysis of randomised controlled trials. Lancet Respir. Med. 2020, 8, 247–257. [Google Scholar] [CrossRef]
- Injarabian, L.; Devin, A.; Ransac, S.; Marteyn, B.S. Neutrophil Metabolic Shift during their Lifecycle: Impact on their Survival and Activation. Int. J. Mol. Sci. 2019, 21, 287. [Google Scholar] [CrossRef]
- Masso-Silva, J.A.; Moshensky, A.; Lam, M.T.; Odish, M.; Patel, A.; Xu, L.; Hansen, E.; Trescott, S.; Nguyen, C.; Kim, R.; et al. Increased peripheral blood neutrophil activation phenotypes and NETosis in critically ill COVID-19 patients: A case series and review of the literature. Clin. Infect. Dis 2021, 74, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Hazeldine, J.; Dinsdale, R.J.; Harrison, P.; Lord, J.M. Traumatic Injury and Exposure to Mitochondrial-Derived Damage Associated Molecular Patterns Suppresses Neutrophil Extracellular Trap Formation. Front. Immunol. 2019, 10, 685. [Google Scholar] [CrossRef] [PubMed]
- Vorobjeva, N.V.; Chernyak, B.V. NETosis: Molecular Mechanisms, Role in Physiology and Pathology. Biochemistry 2020, 85, 1178–1190. [Google Scholar] [CrossRef]
- McIlroy, D.J.; Minahan, K.; Keely, S.; Lott, N.; Hansbro, P.; Smith, D.W.; Balogh, Z.J. Reduced deoxyribonuclease enzyme activity in response to high postinjury mitochondrial DNA concentration provides a therapeutic target for Systemic Inflammatory Response Syndrome. J. Trauma Acute Care Surg. 2018, 85, 354–358. [Google Scholar] [CrossRef]
- Feldman, C.; Anderson, R. The role of co-infections and secondary infections in patients with COVID-19. Pneumonia 2021, 13, 5. [Google Scholar] [CrossRef]
- Buehler, P.K.; Zinkernagel, A.S.; Hofmaenner, D.A.; García, P.D.W.; Acevedo, C.T.; Gómez-Mejia, A.; Shambat, S.M.; Andreoni, F.; Maibach, M.A.; Bartussek, J.; et al. Bacterial pulmonary superinfections are associated with unfavourable outcomes in critically ill COVID-19 patients. medRxiv 2020. [Google Scholar] [CrossRef]
- Zhu, X.; Ge, Y.; Wu, T.; Zhao, K.; Chen, Y.; Wu, B.; Zhu, F.; Zhu, B.; Cui, L. Co-infection with respiratory pathogens among COVID-2019 cases. Virus Res. 2020, 285, 198005. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, G.T.; Green, E.R.; Mecsas, J. Neutrophils to the ROScue: Mechanisms of NADPH Oxidase Activation and Bacterial Resistance. Front. Cell Infect. Microbiol. 2017, 7, 373. [Google Scholar] [CrossRef]
- Veras, F.P.; Pontelli, M.C.; Silva, C.M.; Toller-Kawahisa, J.E.; de Lima, M.; Nascimento, D.C.; Schneider, A.H.; Caetité, D.; Tavares, L.A.; Paiva, I.M.; et al. SARS-CoV-2-triggered neutrophil extracellular traps mediate COVID-19 pathology. J. Exp. Med. 2020, 217, e20201129. [Google Scholar] [CrossRef]
- Ebrahimi, F.; Giaglis, S.; Hahn, S.; Blum, C.A.; Baumgartner, C.; Kutz, A.; Van Breda, S.V.; Mueller, B.; Schuetz, P.; Christ-Crain, M.; et al. Markers of neutrophil extracellular traps predict adverse outcome in community-acquired pneumonia: Secondary analysis of a randomised controlled trial. Eur. Respir. J. 2018, 51, 1701389. [Google Scholar] [CrossRef] [PubMed]
- de Buhr, N.; Parplys, A.C.; Schroeder, M.; Henneck, T.; Schaumburg, B.; Stanelle-Bertram, S.; Jarczak, D.; Nierhaus, A.; Hiller, J.; Peine, S. Impaired Degradation of Neutrophil Extracellular Traps: A Possible Severity Factor of Elderly Male COVID-19 Patients. J. Innate Immun. 2022, 14, 461–476. [Google Scholar] [CrossRef] [PubMed]
- Messner, C.B.; Demichev, V.; Wendisch, D.; Michalick, L.; White, M.; Freiwald, A.; Textoris-Taube, K.; Vernardis, S.I.; Egger, A.-S.; Kreidl, M.; et al. Ultra-High-Throughput Clinical Proteomics Reveals Classifiers of COVID-19 Infection. Cell Syst. 2020, 11, 11–24.e4. [Google Scholar] [CrossRef]
- Overmyer, K.A.; Shishkova, E.; Miller, I.J.; Balnis, J.; Bernstein, M.N.; Peters-Clarke, T.M.; Meyer, J.G.; Quan, Q.; Muehlbauer, L.K.; Trujillo, E.A.; et al. Large-Scale Multi-omic Analysis of COVID-19 Severity. Cell Syst. 2021, 12, 23–40.e7. [Google Scholar] [CrossRef]
- Shen, B.; Yi, X.; Sun, Y.; Bi, X.; Du, J.; Zhang, C.; Quan, S.; Zhang, F.; Sun, R.; Qian, L.; et al. Proteomic and Metabolomic Characterization of COVID-19 Patient Sera. Cell 2020, 182, 59–72.e15. [Google Scholar] [CrossRef]
- Adrover, J.M.; Carrau, L.; Daßler-Plenker, J.; Bram, Y.; Chandar, V.; Houghton, S.; Redmond, D.; Merrill, J.R.; Shevik, M.; Lyons, S.K.; et al. Disulfiram inhibits neutrophil extraelular trap formation protecting rodents from acute lung injury and SARS-CoV2 infection. J. Clin. Investig. 2022, 7, e157342. [Google Scholar]
- Andrew, N.; Insall, R.H. Chemotaxis in shallow gradients is mediated independently of PtdIns 3-kinase by biased choices between random protrusions. Nat. Cell Biol. 2007, 9, 193–200. [Google Scholar] [CrossRef]
- Lee, W.L.; Harrison, R.E.; Grinstein, S. Phagocytosis by neutrophils. Microbes Infect. 2003, 5, 1299–1306. [Google Scholar] [CrossRef]
- Stephens, L.; Ellson, C.; Hawkins, P. Roles of PI3Ks in leukocyte chemotaxis and phagocytosis. Curr. Opin. Cell Biol. 2002, 14, 203–213. [Google Scholar] [CrossRef]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2018, 18, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Erpenbeck, L.; Gruhn, A.L.; Kudryasheva, G.; Günay, G.; Meyer, D.; Busse, J.; Neubert, E.; Schön, M.P.; Rehfeldt, F.; Kruss, S. Effect of Adhesion and Substrate Elasticity on Neutrophil Extracellular Trap Formation. Front. Immunol. 2019, 10, 2320. [Google Scholar] [CrossRef] [Green Version]
- DeSouza-Vieira, T.; Guimarães-Costa, A.; Rochael, N.C.; Lira, M.N.; Nascimento, M.T.; Lima-Gomez, P.D.S.; Mariante, R.M.; Persechini, P.M.; Saraiva, E.M. Neutrophil extracellular traps release induced by Leishmania: Role of PI3Kγ, ERK, PI3Kσ, PKC, and [Ca2+]. J. Leukoc. Biol. 2016, 100, 801–810. [Google Scholar] [CrossRef]
- Palma, G.; Pasqua, T.; Silvestri, G.; Rocca, C.; Gualtieri, P.; Barbieri, A.; De Bartolo, A.; De Lorenzo, A.; Angelone, T.; Avolio, E.; et al. PI3Kδ Inhibition as a Potential Therapeutic Target in COVID-19. Front. Immunol. 2020, 11, 2094. [Google Scholar] [CrossRef]
- Lewis, S.M.; Treacher, D.F.; Edgeworth, J.; Mahalingam, G.; Brown, C.S.; Mare, A.T.; Stacey, M.; Beale, R.; Brown, K.A. Expression of CD11c and EMR2 on neutrophils: Potential diagnostic biomarkers for sepsis and systemic inflammation. Clin. Exp. Immunol. 2015, 182, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Marini, O.; Costa, S.; Bevilacqua, D.; Calzetti, F.; Tamassia, N.; Spina, C.; De Sabata, D.; Tinazzi, E.; Lunardi, C.; Scupoli, M.T.; et al. Mature CD10(+) and immature CD10(−) neutrophils present in G-CSF-treated donors display opposite effects on T cells. Blood 2017, 129, 1343–1356. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Burdon, P.C.; Bridger, G.; Gutierrez-Ramos, J.-C.; Williams, T.J.; Rankin, S.M. Chemokines acting via CXCR2 and CXCR4 control the release of neutrophils from the bone marrow and their return following senescence. Immunity 2003, 19, 583–593. [Google Scholar] [CrossRef]
- Wilk, A.J.; Rustagi, A.; Zhao, N.Q.; Roque, J.; Martínez-Colón, G.J.; McKechnie, J.L.; Ivison, G.T.; Ranganath, T.; Vergara, R.; Hollis, T.; et al. A single-cell atlas of the peripheral immune response in patients with severe COVID-19. Nat. Med. 2020, 26, 1070–1076. [Google Scholar] [CrossRef]
- Buckley, C.D.; Ross, E.; McGettrick, H.; Osborne, C.E.; Haworth, O.; Schmutz, C.; Stone, P.C.W.; Salmon, M.; Matharu, N.M.; Vohra, R.K.; et al. Identification of a phenotypically and functionally distinct population of long-lived neutrophils in a model of reverse endothelial migration. J. Leukoc. Biol. 2006, 79, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Metzemaekers, M.; Cambier, S.; Blanter, M.; Vandooren, J.; de Carvalho, A.C.; Malengier-Devlies, B.; Vanderbeke, L.; Jacobs, C.; Coenen, S.; Martens, E.; et al. Kinetics of peripheral blood neutrophils in severe coronavirus disease 2019. Clin. Transl. Immunol. 2021, 10, e1271. [Google Scholar] [CrossRef] [PubMed]
- Carvelli, J.; Demaria, O.; Vély, F.; Batista, L.; Benmansour, N.C.; Fares, J.; Carpentier, S.; Thibult, M.-L.; Morel, A.; Remark, R.; et al. Association of COVID-19 inflammation with activation of the C5a-C5aR1 axis. Nature 2020, 588, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Shambat, S.M.; Gómez-Mejia, A.; Schweizer, T.A.; Huemer, M.; Chang, C.C.; Acevedo, C.; Pijuan, J.B.; Vulin, C.; Miroshnikova, N.; Hofmänner, D.A.; et al. Neutrophil and monocyte dysfunctional effector response towards bacterial challenge in critically-ill COVID-19 patients. bioRxiv 2020. [Google Scholar] [CrossRef]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.C.; Uhl, S.; Hoagland, D.; Møller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020, 181, 1036–1045.e9. [Google Scholar] [CrossRef]
- Thwaites, R.S.; Uruchurtu, A.S.S.; Siggins, M.K.; Liew, F.; Russell, C.D.; Moore, S.C.; Fairfield, C.; Carter, E.; Abrams, S.; Short, C.-E.; et al. Inflammatory profiles across the spectrum of disease reveal a distinct role for GM-CSF in severe COVID-19. Sci. Immunol. 2021, 6, eabg9873. [Google Scholar] [CrossRef]
- Zuo, Y.; Zuo, M.; Yalavarthi, S.; Gockman, K.; Madison, J.A.; Shi, H.; Woodard, W.; Lezak, S.P.; Lugogo, N.L.; Knight, J.S.; et al. Neutrophil extracellular traps and thrombosis in COVID-19. J. Thromb. Thrombolysis 2021, 51, 446–453. [Google Scholar] [CrossRef]
- Loyer, C.; Lapostolle, A.; Urbina, T.; Elabbadi, A.; Lavillegrand, J.-R.; Chaigneau, T.; Simoes, C.; Dessajan, J.; Desnos, C.; Morin-Brureau, M.; et al. Impairment of neutrophil functions and homeostasis in COVID-19 patients: Association with disease severity. Crit. Care 2022, 26, 155. [Google Scholar] [CrossRef]
- Parekh, D.; Dancer, R.C.A.; Scott, A.; D’Souza, V.K.; Howells, P.A.; Mahida, R.Y.; Tang, J.C.Y.; Cooper, M.S.; Fraser, W.D.; Tan, L. Vitamin D to Prevent Lung Injury Following Esophagectomy-A Randomized, Placebo-Controlled Trial. Crit Care Med. 2018, 46, e1128–e1135. [Google Scholar] [CrossRef]
- Dockrell, D.H.; Lee, M.; Lynch, D.H.; Read, R.C. Immune-mediated phagocytosis and killing of Streptococcus pneumoniae are associated with direct and bystander macrophage apoptosis. J. Infect. Dis. 2001, 184, 713–722. [Google Scholar] [CrossRef]
- Bewley, M.A.; Belchamber, K.B.R.; Chana, K.K.; Budd, R.C.; Donaldson, G.; Wedzicha, J.A.; Brightling, C.E.; Kilty, I.; Donnelly, L.E.; Barnes, P.J.; et al. Differential Effects of p38, MAPK, PI3K or Rho Kinase Inhibitors on Bacterial Phagocytosis and Efferocytosis by Macrophages in COPD. PLoS ONE 2016, 11, e0163139. [Google Scholar] [CrossRef] [PubMed]
- van Overveld, F.J.; Demkow, U.A.; Górecka, D.; Zielinski, J.; de Backer, W.A. Inhibitory capacity of different steroids on neutrophil migration across a bilayer of endothelial and bronchial epithelial cells. Eur. J. Pharmacol. 2003, 477, 261–267. [Google Scholar] [CrossRef]
- Zentay, Z.; Sharaf, M.; Qadir, M.; Drafta, D.; Davidson, D. Mechanism for dexamethasone inhibition of neutrophil migration upon exposure to lipopolysaccharide in vitro: Role of neutrophil interleukin-8 release. Pediatric Res. 1999, 46, 406. [Google Scholar] [CrossRef] [PubMed]
- Weijtens, O.; van der Sluijs, F.A.; Schoemaker, R.C.; Lentjes, E.G.; Cohen, A.F.; Romijn, F.P.; van Meurs, J.C. Peribulbar corticosteroid injection: Vitreal and serum concentrations after dexamethasone disodium phosphate injection. Am. J. Ophthalmol. 1997, 123, 358–363. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AMC n = 26 | COVID-19 n = 41 | CAP n = 26 | COVID-19 Pi3K n = 18 | |
|---|---|---|---|---|
| Male:Female | 10:16 | 26:15 | 15:11 | 9:9 |
| White:Non-white | 24:2 | 31:10 | 23:3 | 13:5 |
| Died (%) | 0 (0%) | 10 (24%) | 2 (8%) | 5 (28%) |
| Age | 70 (61.0–78.0) | 71.5 (58.0–84.0) | 67.5 (54.5–85.0) | 70.7 (51.0–83.0) |
| BMI (kg/m2) | 26.4 (24.2–32.8) | 27.9 (23.3–41.1) | 30.1 (23.2–27.2) | |
| Comorbidities | ||||
| Cardiovascular | 4 (15.3%) | 12 (29.3%) | 7 (26.9%) | 7 (38.9%) |
| Respiratory | 0 (0%) | 1 (2.4%) | 4 (15.4%) | 2 (11.1%) |
| Endocrine | 10 (38.4%) | 16 (39.0%) | 3 (11.5%)* | 9 (50.0%) |
| Hypertension | 13 (50%) | 19 (46.3%) | 10 (38.5%) | 9 (50.0%) |
| Other | 11 (42.3%) | 24 (58.5%) | 15 (57.7%) | 8 (44.4%) |
| WCC (×109/L) | 8.2 (6.3–12.0) | 13.4 (10.7–16.4) *** | 7.6 (4.6–11.9) | |
| Neutrophils (×109/L) | 6.4 (4.4–8.6) | 11.2 (7.8–13.6) ** | 5.8 (2.9–9.4) | |
| CRP (mg/L) | 103.0 (63.0–165.0) | 119.0 (42.0–396.0) | 74 (24.3–161.3) | |
| NLR | 5.4 (3.8–10.8) | 8.5 (5.9–52.7) | 6.3 (2.4–10.2) | |
| Worst NEWS2 | 6.0 (5.0–7.0) | 5.0 (3.0–12.0) | 6.0 (3.0–8.0) | |
| HS Troponin I (ng/L) | 14.5 (5.0–31.3) | 17.5 (4.0–318.0) | 7.0 (4.0–31.5) | |
| D-dimer (ng/mL) | 382.0 (218.0–829.5) | 659.0 (270.5–1510.0) | 493.0 (247.0–890.0) | |
| Ferritin (ug/L) | 1082 (428.3–1525.0) | 110.0 (78.8–225.3) * | 251.5 (185.8–1230.3) | |
| Vitamin D (nmol/L) | 35.6 (23.0–51.8) | 45.3 (23.8–73.2) | 36.6 (22.6–60.1) | |
| Dexamethasone | 38 (92.6%) | 0 (0%) | 10 (55.6%) * | |
| Admission | ||||
| 4C | 12.0 (9.0–14.0) | 13 (11.0–15.8) | ||
| qSOFA | 1.0 (1.0–1.5) | 1.0 (1.0–1.0) | 1.0 (1.0–1.0) | |
| CURB-65 | 2.0 (1.0–3.0) | 2.0 (2.0–2.0) | 2.0 (2.0–3.0) | |
| Length of stay (days) | 5.5 (3.0–12.0) | 4 (3.0–7.5) | 6.5 (3.3–14.8) |
| Receptors | % Expression | MFI | ||||||
|---|---|---|---|---|---|---|---|---|
| AMC n = 26 | COVID n = 34 | CAP n = 16 | p Value | AMC n = 26 | COVID n = 34 | CAP n = 16 | p Value | |
| CD10 | 94.3 (11.52) | 95.3 (13.6) | 74.7 *# (60.6) | * <0.0006 # <0.0006 | 669 (511) | 836 (518) | 701 (486) | 0.304 |
| CD11b | 81.6 (15.25) | 67.8 *$ (43.9) | 83.9 (23.8) | * 0.0014 $ 0.046 | 399 (331) | 40 *$ (223) | 1701 * (1268) | * 0.0026 $ <0.0001 * <0.0001 |
| CD54 | 26.2 (37.0) | 71.3 *$ (21.2) | 6.6 (10.9) | * <0.0001 $ <0.0001 | 33 (48) | 73 (58) | 147 *# (77) | * 0.0001 # 0.0012 |
| CD62L | 23.3 (58.8) | 31.0 (15.3) | 49.9 (41.4) | 0.384 | 68 (400) | 94 (121) | 723 *# (519) | * 0.0005 # <0.0001 |
| CXCR2 | 99.9 (0.1) | 100 (0.2) | 98.5 (1.6) | 0.741 | 2482 (1415) | 2031 (1600) | 2839 (2839) | 0.439 |
| CXCR4 | 93.9 (16.4) | 96.7 (9.07) | 54.3 *# (35.0) | * <0.0001 # <0.0001 | 1917 (4039) | 1724$ (4177) | 1090 (1376) | $ 0.028 |
| CD66b | 99.7 (0.5) | 99.8 (0.3) | 97.6 (1.6) | 0.298 | 324 (367) | 186 (161) | 1631 *# (1493) | * <0.0001 # <0.0001 |
| CD11c | 99.2 (2.9) | 99.9 (1.05) | 76.1 *# (64.5) | * <0.0065 # <0.0001 | 392 (2120) | 1148 (670) | 4197 *# (6040) | * <0.0001 # <0.0001 |
| PD-L1 | 63.8 (32) | 98 (27.9) | 8.7 *# (12.9) | * <0.0001 # <0.0001 | 193 (210) | 70 *$ (132) | 524 * (264) | * 0.006 $ <0.0001 * 0.0006 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belchamber, K.B.R.; Thein, O.S.; Hazeldine, J.; Grudzinska, F.S.; Faniyi, A.A.; Hughes, M.J.; Jasper, A.E.; Yip, K.P.; Crowley, L.E.; Lugg, S.T.; et al. Dysregulated Neutrophil Phenotype and Function in Hospitalised Non-ICU COVID-19 Pneumonia. Cells 2022, 11, 2901. https://doi.org/10.3390/cells11182901
Belchamber KBR, Thein OS, Hazeldine J, Grudzinska FS, Faniyi AA, Hughes MJ, Jasper AE, Yip KP, Crowley LE, Lugg ST, et al. Dysregulated Neutrophil Phenotype and Function in Hospitalised Non-ICU COVID-19 Pneumonia. Cells. 2022; 11(18):2901. https://doi.org/10.3390/cells11182901
Chicago/Turabian StyleBelchamber, Kylie B. R., Onn S. Thein, Jon Hazeldine, Frances S. Grudzinska, Aduragbemi A. Faniyi, Michael J. Hughes, Alice E. Jasper, Kay Por Yip, Louise E. Crowley, Sebastian T. Lugg, and et al. 2022. "Dysregulated Neutrophil Phenotype and Function in Hospitalised Non-ICU COVID-19 Pneumonia" Cells 11, no. 18: 2901. https://doi.org/10.3390/cells11182901