
Peripheral Nerve Regeneration–Adipose-Tissue-Derived Stem Cells Differentiated by a Three-Step Protocol Promote Neurite Elongation via NGF Secretion
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. ASC Cell Culture
2.2. Differentiation Protocols
2.3. NG108-15 Cell Culture
2.4. Coculture
2.5. RNA Isolation/Reverse Transcription PCR
2.6. Western Blot
2.7. ELISA
2.8. Neurite-Outgrowth Assay
2.9. Surface Coating
2.10. Statistical Analysis
3. Results
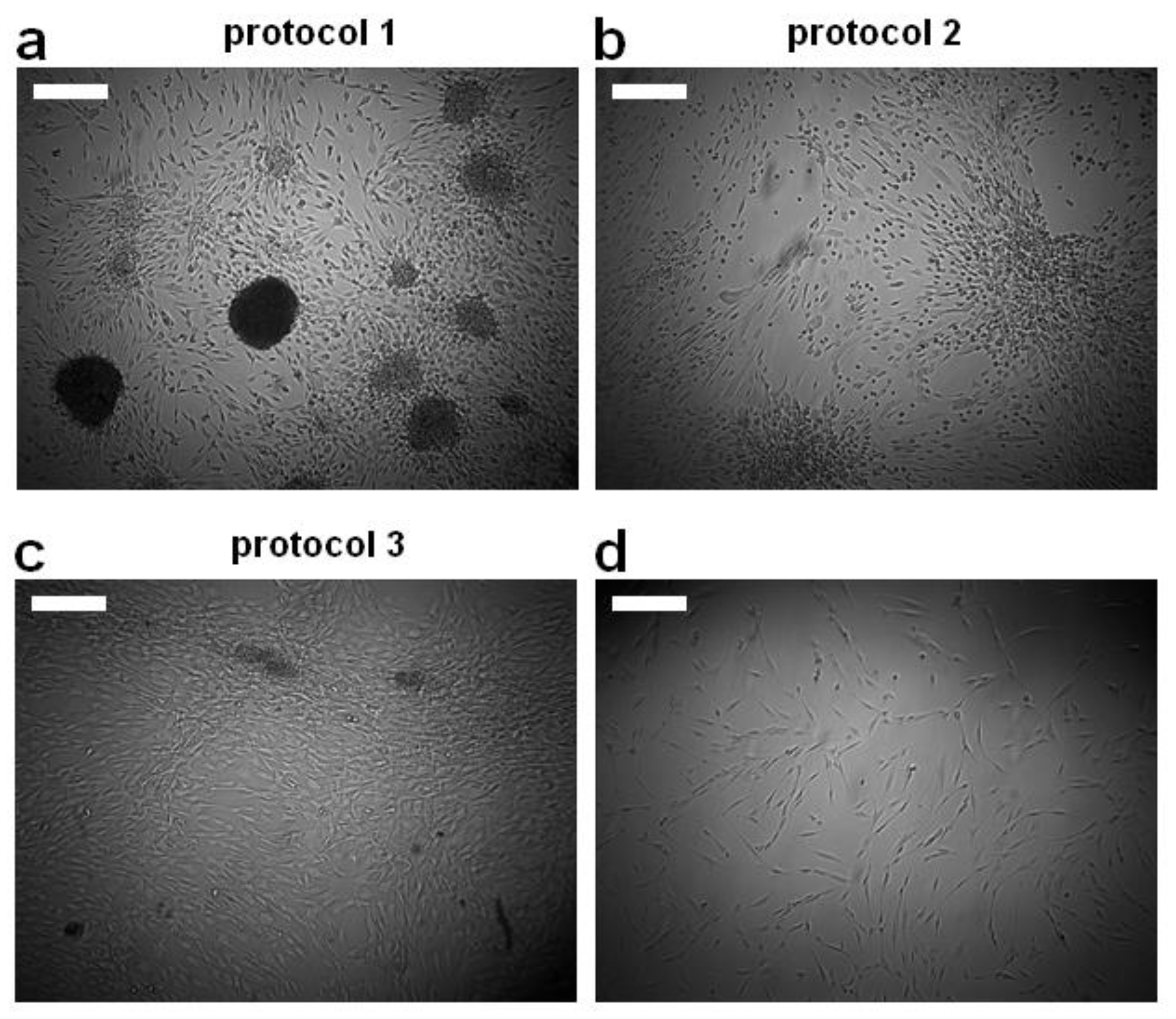
3.1. Morphological Appearance
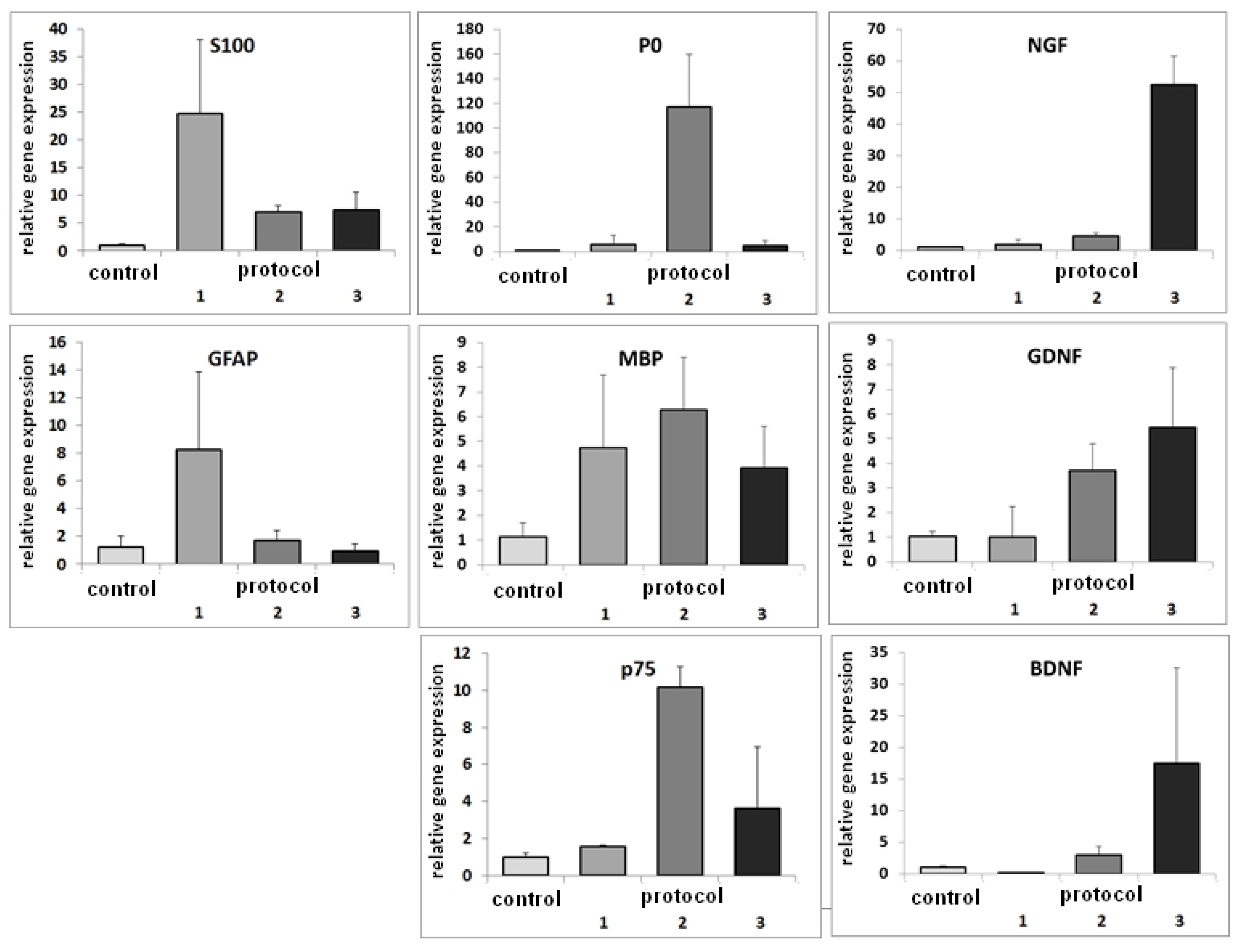
3.2. RT-PCR after Differentiation of ASCs with Basic Protocols
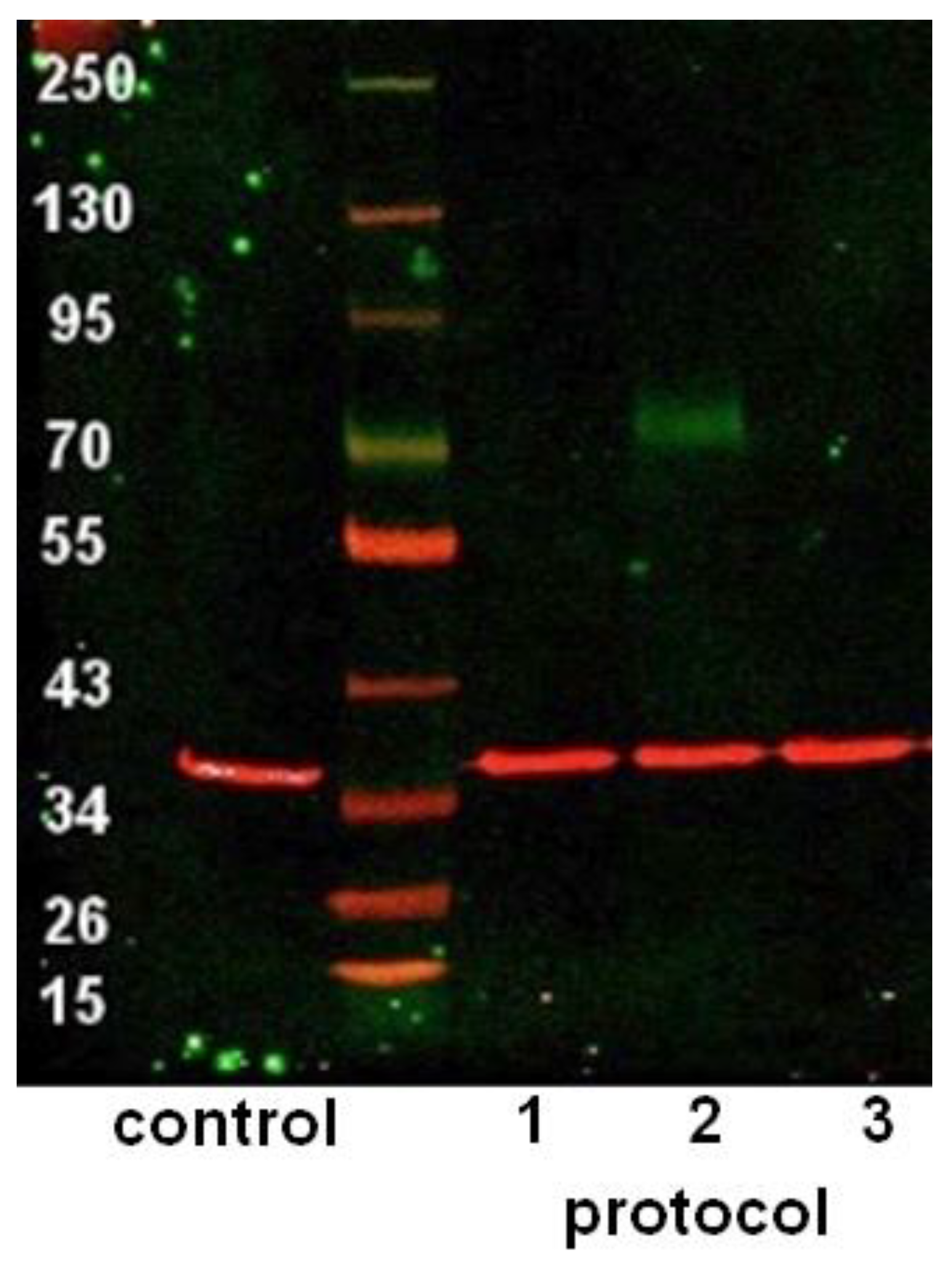
3.3. Western Blot after Differentiation of ASCs with Basic Protocols
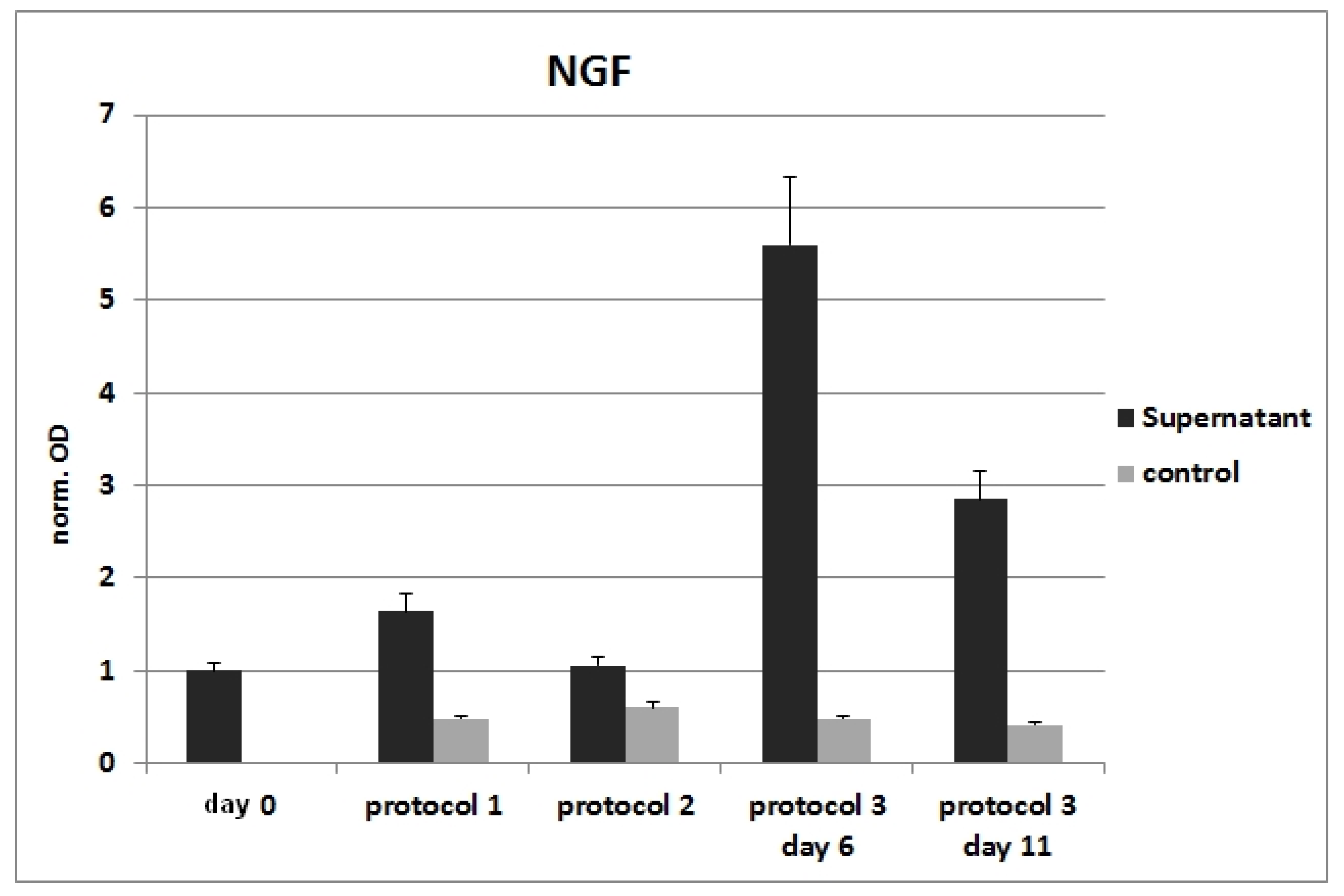
3.4. ELISA after Differentiation with Basic Protocols
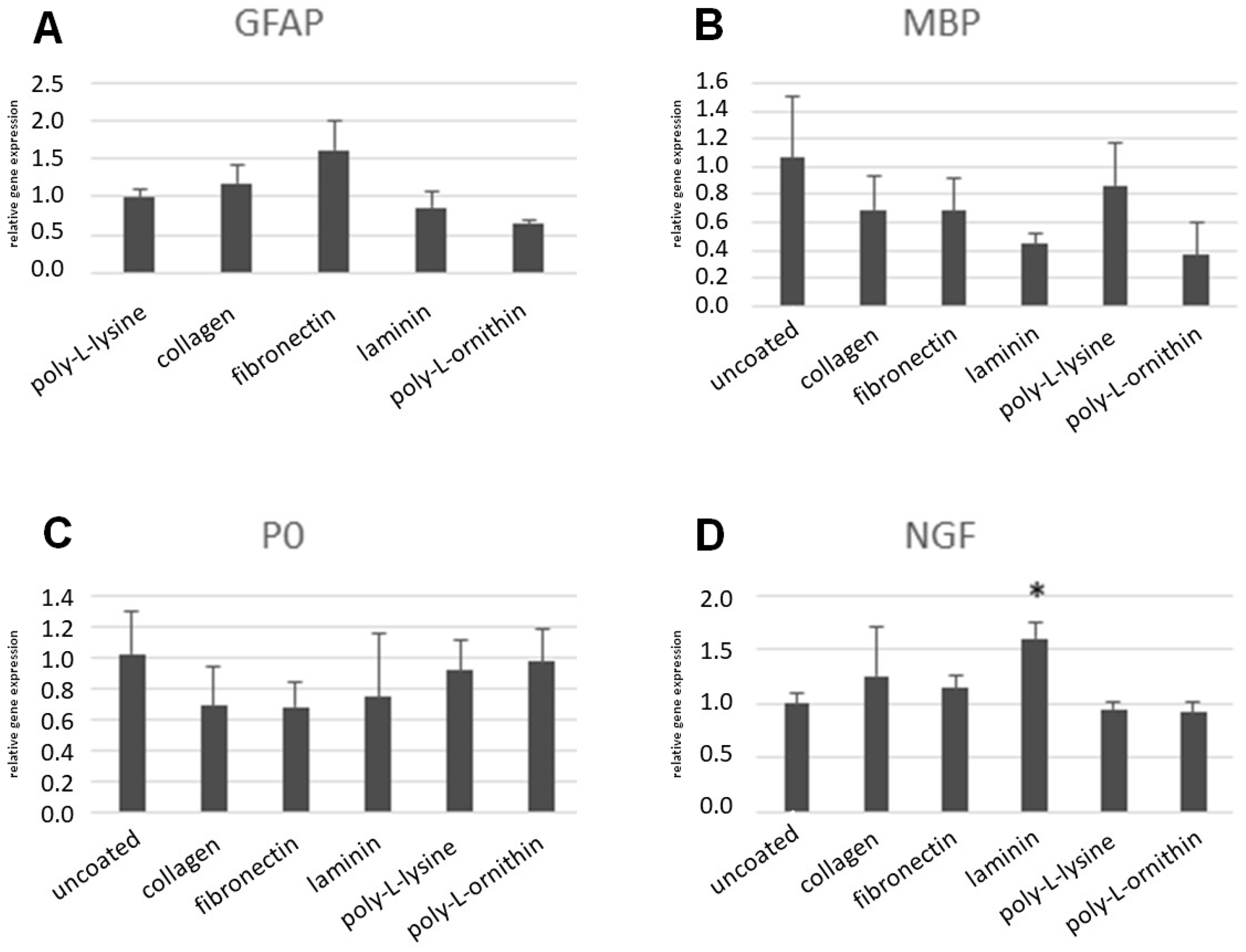
3.5. RT-PCR after Differentiation of ASCs on Coated Surfaces
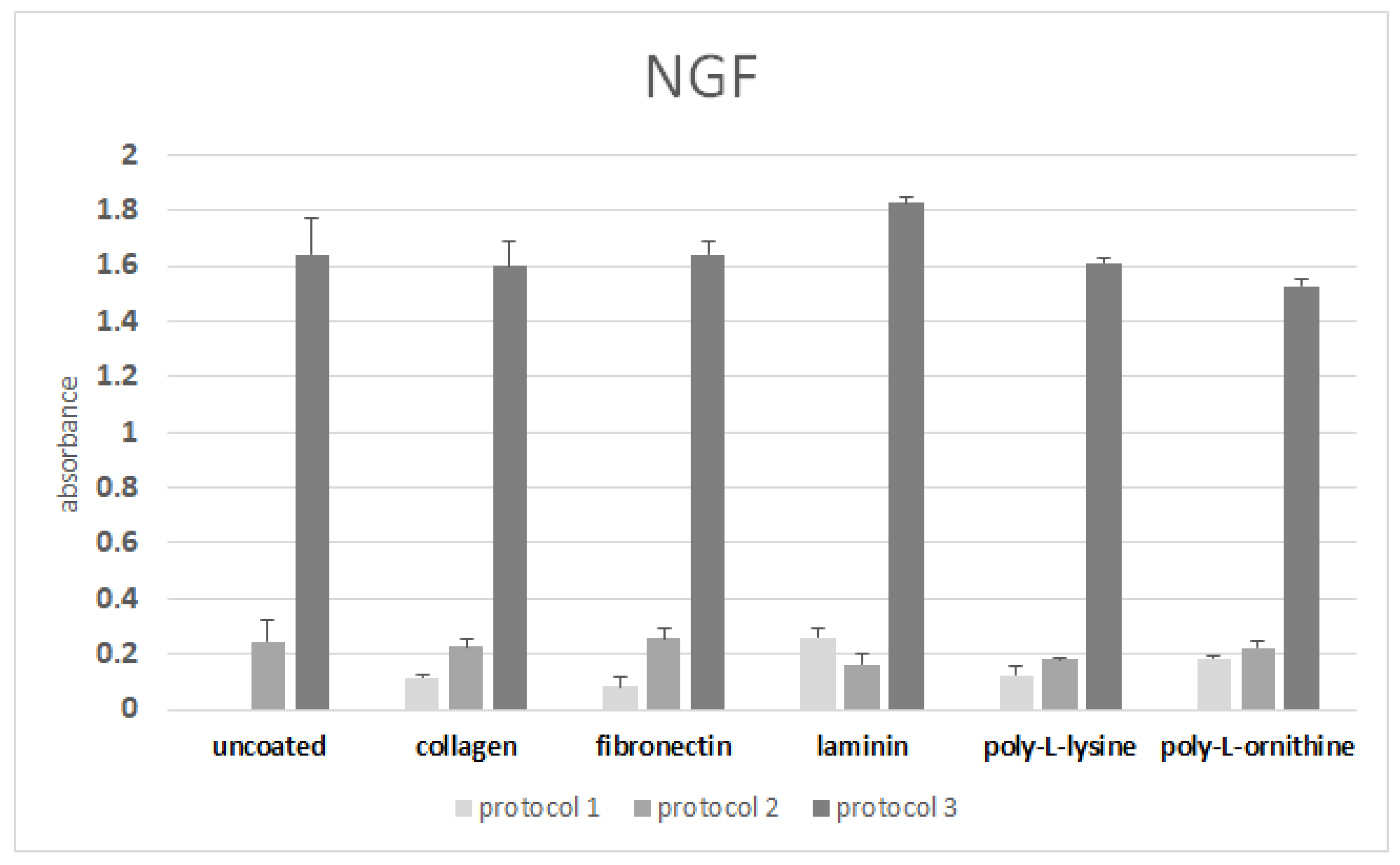
3.6. ELISA after Differentiation on Coated Surfaces
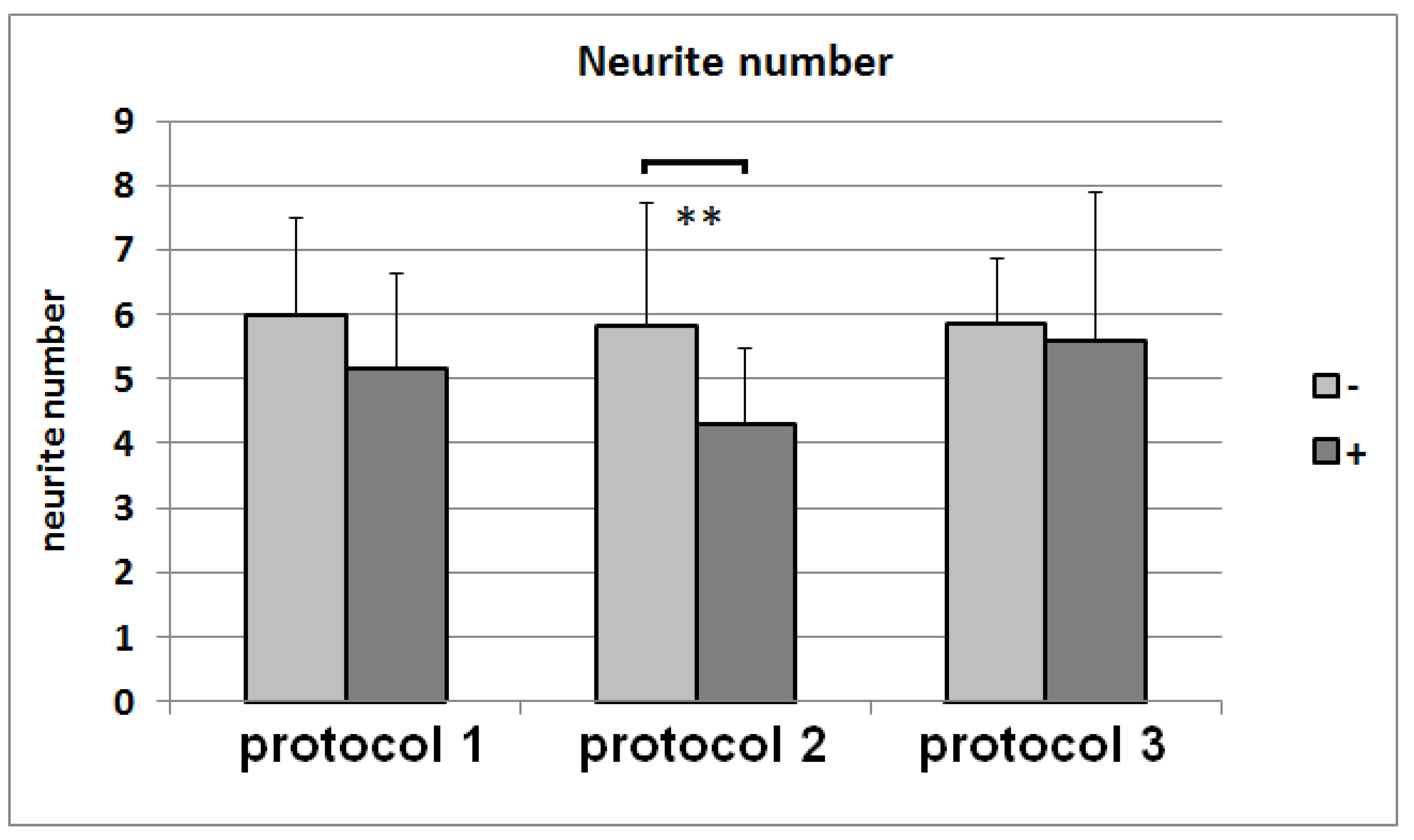
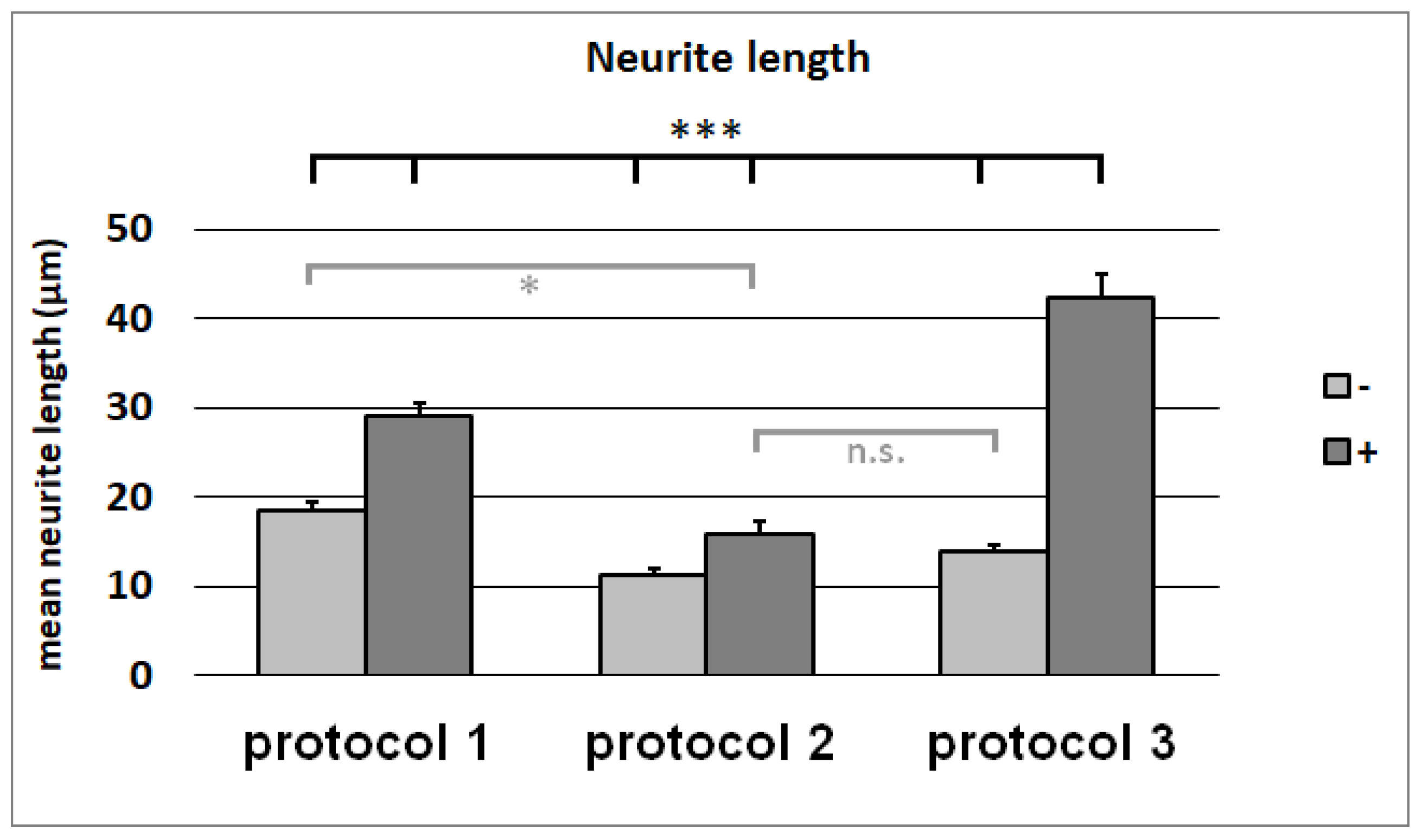
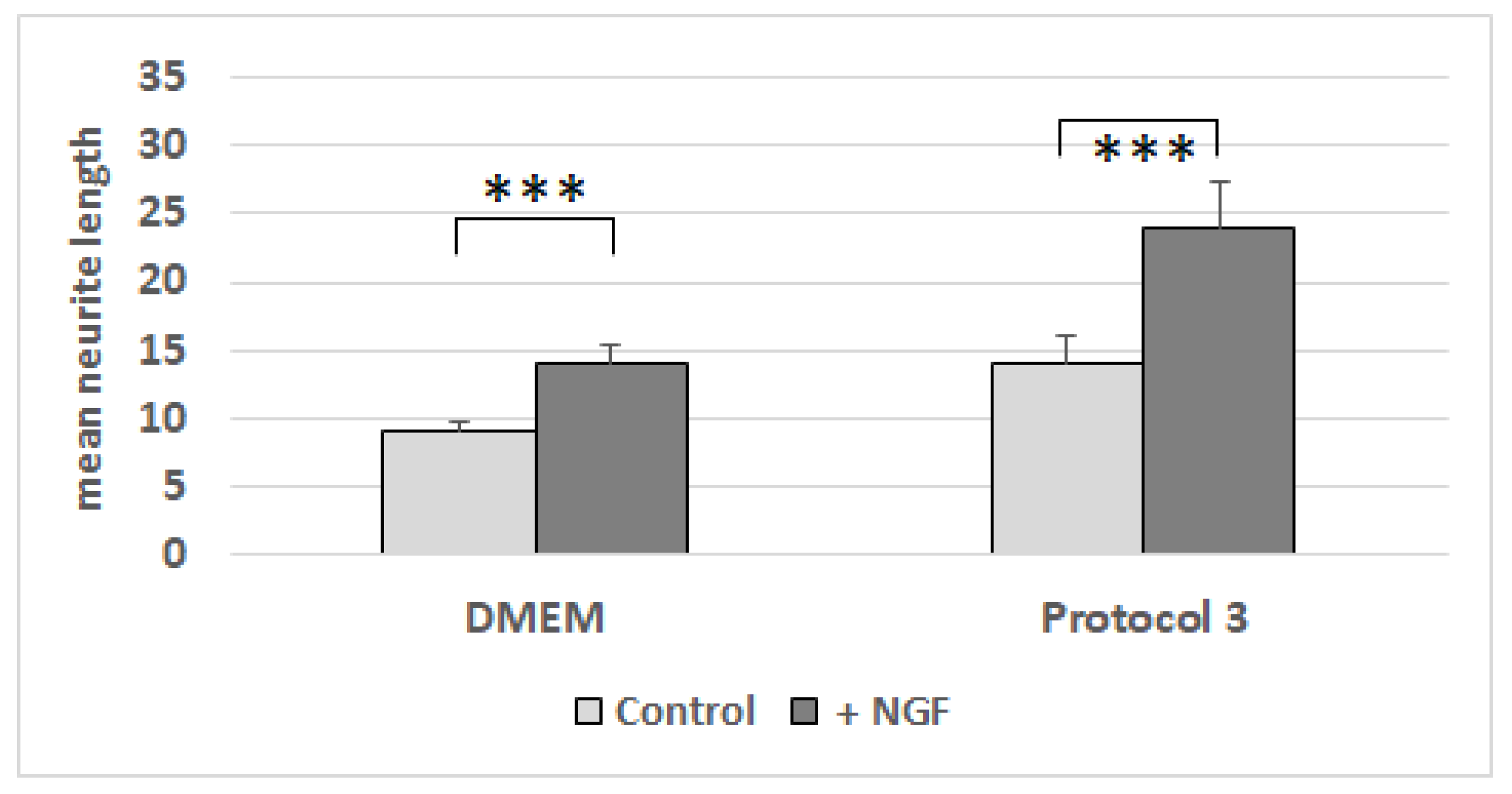
3.7. Neurite-Outgrowth Assay
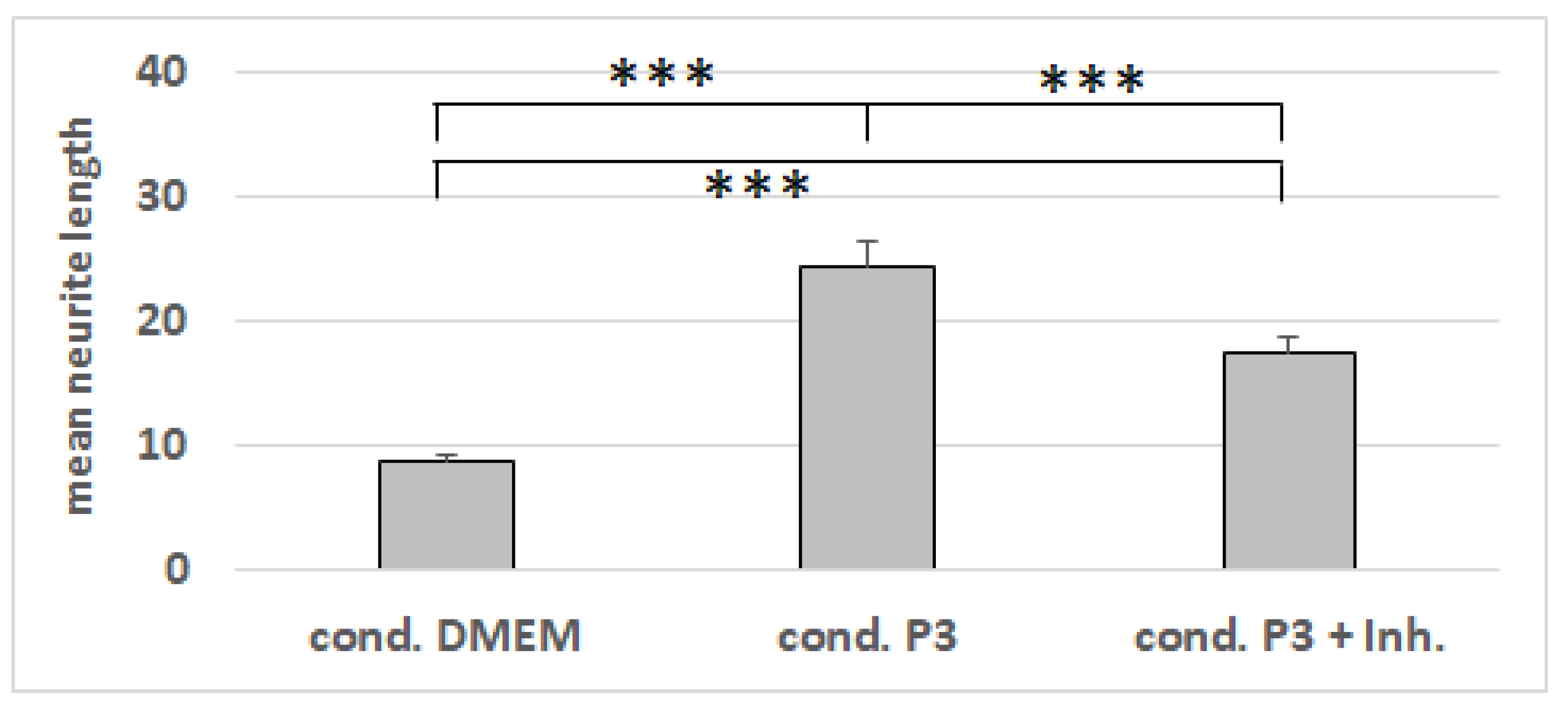
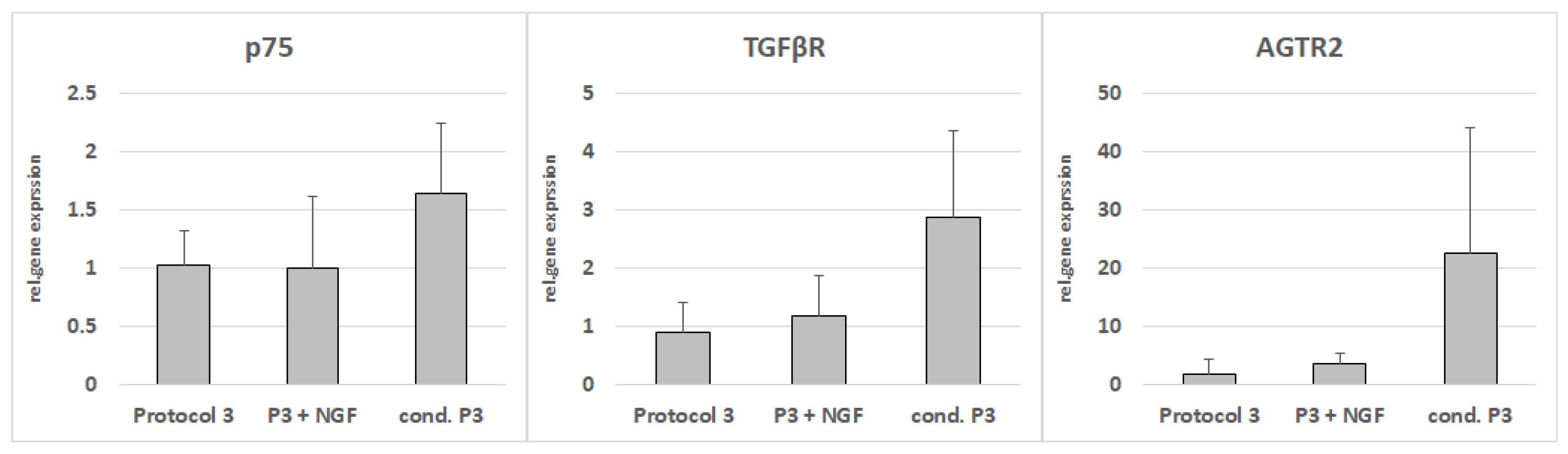
3.8. RT-PCR from NG108-15 Cells after Cultivation with Conditioned Media
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bhatheja, K.; Field, J. Schwann Cells: Origins and Role in Axonal Maintenance and Regeneration. Int. J. Biochem. Cell Biol. 2006, 38, 1995–1999. [Google Scholar] [CrossRef]
- Richner, M.; Ulrichsen, M.; Elmegaard, S.L.; Dieu, R.; Pallesen, L.T.; Vaegter, C.B. Peripheral Nerve Injury Modulates Neurotrophin Signaling in the Peripheral and Central Nervous System. Mol. Neurobiol. 2014, 50, 945–970. [Google Scholar] [CrossRef]
- Jessen, K.R.; Mirsky, R. Negative Regulation of Myelination: Relevance for Development, Injury, and Demyelinating Disease. Glia 2008, 56, 1552–1565. [Google Scholar] [CrossRef]
- Arthur-Farraj, P.J.; Latouche, M.; Wilton, D.K.; Quintes, S.; Chabrol, E.; Banerjee, A.; Woodhoo, A.; Jenkins, B.; Rahman, M.; Turmaine, M.; et al. C-Jun Reprograms Schwann Cells of Injured Nerves to Generate a Repair Cell Essential for Regeneration. Neuron 2012, 75, 633–647. [Google Scholar] [CrossRef]
- Jessen, K.R.; Mirsky, R.; Lloyd, A.C. Schwann Cells: Development and Role in Nerve Repair. Cold Spring Harb. Perspect. Biol. 2015, 7, a020487. [Google Scholar] [CrossRef]
- Dai, L.-G.; Huang, G.-S.; Hsu, S. Sciatic Nerve Regeneration by Cocultured Schwann Cells and Stem Cells on Microporous Nerve Conduits. Cell Transplant. 2013, 22, 2029–2039. [Google Scholar] [CrossRef]
- Gonzalez-Perez, F.; Hernández, J.; Heimann, C.; Phillips, J.B.; Udina, E.; Navarro, X. Schwann Cells and Mesenchymal Stem Cells in Laminin- or Fibronectin-Aligned Matrices and Regeneration across a Critical Size Defect of 15 Mm in the Rat Sciatic Nerve. J. Neurosurg. Spine 2018, 28, 109–118. [Google Scholar] [CrossRef]
- Hsu, M.-N.; Liao, H.-T.; Li, K.-C.; Chen, H.-H.; Yen, T.-C.; Makarevich, P.; Parfyonova, Y.; Hu, Y.-C. Adipose-Derived Stem Cell Sheets Functionalized by Hybrid Baculovirus for Prolonged GDNF Expression and Improved Nerve Regeneration. Biomaterials 2017, 140, 189–200. [Google Scholar] [CrossRef]
- Klein, S.M.; Vykoukal, J.; Li, D.-P.; Pan, H.-L.; Zeitler, K.; Alt, E.; Geis, S.; Felthaus, O.; Prantl, L. Peripheral Motor and Sensory Nerve Conduction Following Transplantation of Undifferentiated Autologous Adipose Tissue-Derived Stem Cells in a Biodegradable U.S. Food and Drug Administration-Approved Nerve Conduit. Plast. Reconstr. Surg. 2016, 138, 132–139. [Google Scholar] [CrossRef]
- Orbay, H.; Uysal, A.C.; Hyakusoku, H.; Mizuno, H. Differentiated and Undifferentiated Adipose-Derived Stem Cells Improve Function in Rats with Peripheral Nerve Gaps. J. Plast. Reconstr. Aesthetic Surg. JPRAS 2012, 65, 657–664. [Google Scholar] [CrossRef]
- Reid, A.J.; Sun, M.; Wiberg, M.; Downes, S.; Terenghi, G.; Kingham, P.J. Nerve Repair with Adipose-Derived Stem Cells Protects Dorsal Root Ganglia Neurons from Apoptosis. Neuroscience 2011, 199, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Yokoi, T.; Uemura, T.; Takamatsu, K.; Shintani, K.; Onode, E.; Okada, M.; Hidaka, N.; Nakamura, H. Bioabsorbable Nerve Conduits Coated with Induced Pluripotent Stem Cell-Derived Neurospheres Enhance Axonal Regeneration in Sciatic Nerve Defects in Aged Mice. J. Biomed. Mater. Res. B Appl. Biomater. 2017, 106, 1752–1758. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Liu, Z.; Liu, L.; Zhao, C.; Xiong, F.; Zhou, C.; Li, Y.; Shan, Y.; Peng, F.; Zhang, C. Neurospheres from Rat Adipose-Derived Stem Cells Could Be Induced into Functional Schwann Cell-like Cells in Vitro. BMC Neurosci. 2008, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Felthaus, O.; Ernst, W.; Driemel, O.; Reichert, T.E.; Schmalz, G.; Morsczeck, C. TGF-Beta Stimulates Glial-like Differentiation in Murine Dental Follicle Precursor Cells (MDFPCs). Neurosci. Lett. 2010, 471, 179–184. [Google Scholar] [CrossRef]
- Kingham, P.J.; Kalbermatten, D.F.; Mahay, D.; Armstrong, S.J.; Wiberg, M.; Terenghi, G. Adipose-Derived Stem Cells Differentiate into a Schwann Cell Phenotype and Promote Neurite Outgrowth in Vitro. Exp. Neurol. 2007, 207, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.; Prantl, L.; Vykoukal, J.; Loibl, M.; Felthaus, O. Differential Effects of Coating Materials on Viability and Migration of Schwann Cells. Materials 2016, 9, 150. [Google Scholar] [CrossRef]
- Urrutia, D.N.; Caviedes, P.; Mardones, R.; Minguell, J.J.; Vega-Letter, A.M.; Jofre, C.M. Comparative Study of the Neural Differentiation Capacity of Mesenchymal Stromal Cells from Different Tissue Sources: An Approach for Their Use in Neural Regeneration Therapies. PLoS ONE 2019, 14, e0213032. [Google Scholar] [CrossRef]
- Zhou, L.N.; Wang, J.C.; Zilundu, P.L.M.; Wang, Y.Q.; Guo, W.P.; Zhang, S.X.; Luo, H.; Zhou, J.H.; Deng, R.D.; Chen, D.F. A Comparison of the Use of Adipose-Derived and Bone Marrow-Derived Stem Cells for Peripheral Nerve Regeneration in Vitro and in Vivo. Stem Cell Res. Ther. 2020, 11, 153. [Google Scholar] [CrossRef]
- Wu, S.-H.; Liao, Y.-T.; Hsueh, K.-K.; Huang, H.-K.; Chen, T.-M.; Chiang, E.-R.; Hsu, S.; Tseng, T.-C.; Wang, J.-P. Adipose-Derived Mesenchymal Stem Cells from a Hypoxic Culture Improve Neuronal Differentiation and Nerve Repair. Front. Cell Dev. Biol. 2021, 9, 658099. [Google Scholar] [CrossRef]
- Acheson, A.; Barker, P.A.; Alderson, R.F.; Miller, F.D.; Murphy, R.A. Detection of Brain-Derived Neurotrophic Factor-like Activity in Fibroblasts and Schwann Cells: Inhibition by Antibodies to NGF. Neuron 1991, 7, 265–275. [Google Scholar] [CrossRef]
- Taniuchi, M.; Clark, H.B.; Johnson, J.E. Induction of Nerve Growth Factor Receptor in Schwann Cells after Axotomy., Induction of Nerve Growth Factor Receptor in Schwann Cells after Axotomy. Proc. Natl. Acad. Sci. USA 1986, 83, 4094–4098. [Google Scholar] [CrossRef] [PubMed]
- Neet, K.E.; Campenot, R.B. Receptor Binding, Internalization, and Retrograde Transport of Neurotrophic Factors. Cell. Mol. Life Sci. CMLS 2001, 58, 1021–1035. [Google Scholar] [CrossRef]
- Tricaud, N.; Park, H.T. Wallerian Demyelination: Chronicle of a Cellular Cataclysm. Cell. Mol. Life Sci. CMLS 2017, 74, 4049–4057. [Google Scholar] [CrossRef]
- Chen, Z.-L.; Yu, W.-M.; Strickland, S. Peripheral Regeneration. Annu. Rev. Neurosci. 2007, 30, 209–233. [Google Scholar] [CrossRef]
- Bhangra, K.S.; Busuttil, F.; Phillips, J.B.; Rahim, A.A. Using Stem Cells to Grow Artificial Tissue for Peripheral Nerve Repair. Stem Cells Int. 2016, 2016. [Google Scholar] [CrossRef]
- Clements, M.P.; Byrne, E.; Camarillo Guerrero, L.F.; Cattin, A.-L.; Zakka, L.; Ashraf, A.; Burden, J.J.; Khadayate, S.; Lloyd, A.C.; Marguerat, S.; et al. The Wound Microenvironment Reprograms Schwann Cells to Invasive Mesenchymal-like Cells to Drive Peripheral Nerve Regeneration. Neuron 2017, 96, 98–114. [Google Scholar] [CrossRef]
- Boyd, J.G.; Gordon, T. Neurotrophic Factors and Their Receptors in Axonal Regeneration and Functional Recovery after Peripheral Nerve Injury. Mol. Neurobiol. 2003, 27, 277–324. [Google Scholar] [CrossRef]
- Aszmann, O.C.; Korak, K.J.; Kropf, N.; Fine, E.; Aebischer, P.; Frey, M. Simultaneous GDNF and BDNF Application Leads to Increased Motoneuron Survival and Improved Functional Outcome in an Experimental Model for Obstetric Brachial Plexus Lesions. Plast. Reconstr. Surg. 2002, 110, 1066–1072. [Google Scholar] [CrossRef]
- Li, R.; Wu, J.; Lin, Z.; Nangle, M.R.; Li, Y.; Cai, P.; Liu, D.; Ye, L.; Xiao, Z.; He, C.; et al. Single Injection of a Novel Nerve Growth Factor Coacervate Improves Structural and Functional Regeneration after Sciatic Nerve Injury in Adult Rats. Exp. Neurol. 2017, 288, 1–10. [Google Scholar] [CrossRef]
- Baumgartner, B.J.; Shine, H.D. Neuroprotection of Spinal Motoneurons Following Targeted Transduction with an Adenoviral Vector Carrying the Gene for Glial Cell Line-Derived Neurotrophic Factor. Exp. Neurol. 1998, 153, 102–112. [Google Scholar] [CrossRef]
- Yan, Q.; Elliott, J.; Snider, W.D. Brain-Derived Neurotrophic Factor Rescues Spinal Motor Neurons from Axotomy-Induced Cell Death. Nature 1992, 360, 753–755. [Google Scholar] [CrossRef]
- Morgan, L.; Jessen, K.R.; Mirsky, R. The Effects of CAMP on Differentiation of Cultured Schwann Cells: Progression from an Early Phenotype (04+) to a Myelin Phenotype (P0+, GFAP-, N-CAM-, NGF-Receptor-) Depends on Growth Inhibition. J. Cell Biol. 1991, 112, 457–467. [Google Scholar] [CrossRef]
- Jessen, K.R.; Mirsky, R. The Repair Schwann Cell and Its Function in Regenerating Nerves., The Repair Schwann Cell and Its Function in Regenerating Nerves. J. Physiol. 2016, 594, 3521–3531. [Google Scholar] [CrossRef] [PubMed]
- Salzer, J.L. Schwann Cell Myelination. Cold Spring Harb. Perspect. Biol. 2015, 7, a020529. [Google Scholar] [CrossRef] [PubMed]
- Di Summa, P.G.; Kingham, P.J.; Raffoul, W.; Wiberg, M.; Terenghi, G.; Kalbermatten, D.F. Adipose-Derived Stem Cells Enhance Peripheral Nerve Regeneration. J. Plast. Reconstr. Aesthetic Surg. JPRAS 2010, 63, 1544–1552. [Google Scholar] [CrossRef]
- Shimizu, S.; Kitada, M.; Ishikawa, H.; Itokazu, Y.; Wakao, S.; Dezawa, M. Peripheral Nerve Regeneration by the in Vitro Differentiated-Human Bone Marrow Stromal Cells with Schwann Cell Property. Biochem. Biophys. Res. Commun. 2007, 359, 915–920. [Google Scholar] [CrossRef]
- Dadon-Nachum, M.; Sadan, O.; Srugo, I.; Melamed, E.; Offen, D. Differentiated Mesenchymal Stem Cells for Sciatic Nerve Injury. Stem Cell Rev. 2011, 7, 664–671. [Google Scholar] [CrossRef]
- Li, Y.; Xu, W.; Cheng, L.-Y. Adipose-Derived Mesenchymal Stem Cells Accelerate Nerve Regeneration and Functional Recovery in a Rat Model of Recurrent Laryngeal Nerve Injury. Neural Regen. Res. 2017, 12, 1544–1550. [Google Scholar] [CrossRef]
- Cai, S.; Tsui, Y.-P.; Tam, K.-W.; Shea, G.K.-H.; Chang, R.S.-K.; Ao, Q.; Shum, D.K.-Y.; Chan, Y.-S. Directed Differentiation of Human Bone Marrow Stromal Cells to Fate-Committed Schwann Cells. Stem Cell Rep. 2017, 9, 1097–1108. [Google Scholar] [CrossRef]
- Yu, W.-M.; Chen, Z.-L.; North, A.J.; Strickland, S. Laminin Is Required for Schwann Cell Morphogenesis. J. Cell Sci. 2009, 122, 929–936. [Google Scholar] [CrossRef] [Green Version]
- Zarinfard, G.; Tadjalli, M.; Razavi, S.; Kazemi, M. Effect of Laminin on Neurotrophic Factors Expression in Schwann-Like Cells Induced from Human Adipose-Derived Stem Cells In Vitro. J. Mol. Neurosci. 2016, 60, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Guimond, M.-O.; Roberge, C.; Gallo-Payet, N. Fyn Is Involved in Angiotensin II Type 2 Receptor-Induced Neurite Outgrowth, but Not in P42/P44mapk in NG108-15 Cells. Mol. Cell. Neurosci. 2010, 45, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Mazini, L.; Ezzoubi, M.; Malka, G. Overview of Current Adipose-Derived Stem Cell (ADSCs) Processing Involved in Therapeutic Advancements: Flow Chart and Regulation Updates before and after COVID-19. Stem Cell Res. Ther. 2021, 12, 1. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protocol 1 [13] | Protocol 2 [14] | Protocol 3 [15] | |
|---|---|---|---|
| Original protocol composition | EGF bFGF ---------------------------------- all-trans-retinoic acid neuregulin 1 PDGF forskolin | N2 Supplement bFGF EGF all-trans-retinoic acid TGF-beta1 | β-mercaptoethanol --------------------------------- all-trans-retinoic acid --------------------------------- PDGF bFGF forskolin neuregulin 1 |
| Additionally tested supplements | FK-506 GAS-6 heregulin beta 1 melatonin TGF-beta 1 | FK-506 forskolin GAS-6 heregulin beta 1 melatonin neuregulin 1 PDGF | FK-506 GAS-6 heregulin beta 1 melatonin TGF-beta 1 |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| AGTR2 | 5’-GGTCTGCTGGGATTGCCTTAATG-3’ | 5’-ACTTGGTCACGGGTAATTCTGTTCT-3’ |
| BDNF | 5’-AATAATGTCTGACCCCAGTGCC-3’ | 5’-ATTGTTGTCACGCTCCTGGT-3’ |
| GAPDH | 5’-GGAGCGAGATCCCTCCAAAAT-3’ | 5’-GGCTGTTGTCATACTTCTCATGG-3’ |
| GDNF | 5’-CGCTGACCAGTGACTCCAATA-3’ | 5’-CTCTGCGACCTTTCCCTCTG-3’ |
| GFAP | 5’-GACACCTGGGTACCATTCCG-3’ | 5’-CATCTTGGAGCTTCTGCCTCA-3’ |
| MBP | 5’-TCTGGCAAGGACTCACACAC-3’ | 5’-TCTGCCTCCGTAGCCAAATC-3’ |
| NGF | 5’-GGCCACTCTGAGGTGCATAG-3’ | 5’-CTGTGTACGGTTCTGCCTGT-3’ |
| P0 | 5’-GGCCATTGTGGTTTACACGG-3’ | 5’-GGAAGATTGAAATGGCATCTCGG-3’ |
| p75 | 5’-AATGCGAAGAGATCCCTGGTC-3’ | 5’-GGAATGAGGTTGTCGGTGGT-3’ |
| S100B | 5’-AGAGGGTGACAAGCACAAGC-3’ | 5’-TCCTGCTCTTTGATTTCCTCCAG-3’ |
| TGFβR | 5’-ATTGGCGGAATCCACGAAGA-3’ | 5’-GTAGAACAACCGGCCTCCAA-3’ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klein, S.; Siegmund, A.; Eigenberger, A.; Hartmann, V.; Langewost, F.; Hammer, N.; Anker, A.; Klein, K.; Morsczeck, C.; Prantl, L.; et al. Peripheral Nerve Regeneration–Adipose-Tissue-Derived Stem Cells Differentiated by a Three-Step Protocol Promote Neurite Elongation via NGF Secretion. Cells 2022, 11, 2887. https://doi.org/10.3390/cells11182887
Klein S, Siegmund A, Eigenberger A, Hartmann V, Langewost F, Hammer N, Anker A, Klein K, Morsczeck C, Prantl L, et al. Peripheral Nerve Regeneration–Adipose-Tissue-Derived Stem Cells Differentiated by a Three-Step Protocol Promote Neurite Elongation via NGF Secretion. Cells. 2022; 11(18):2887. https://doi.org/10.3390/cells11182887
Chicago/Turabian StyleKlein, Silvan, Andreas Siegmund, Andreas Eigenberger, Valerie Hartmann, Felix Langewost, Nicolas Hammer, Alexandra Anker, Konstantin Klein, Christian Morsczeck, Lukas Prantl, and et al. 2022. "Peripheral Nerve Regeneration–Adipose-Tissue-Derived Stem Cells Differentiated by a Three-Step Protocol Promote Neurite Elongation via NGF Secretion" Cells 11, no. 18: 2887. https://doi.org/10.3390/cells11182887