
Sugarcane Streak Mosaic Virus P1 Attenuates Plant Antiviral Immunity and Enhances Potato Virus X Infection in Nicotiana benthamiana

 , and
, and 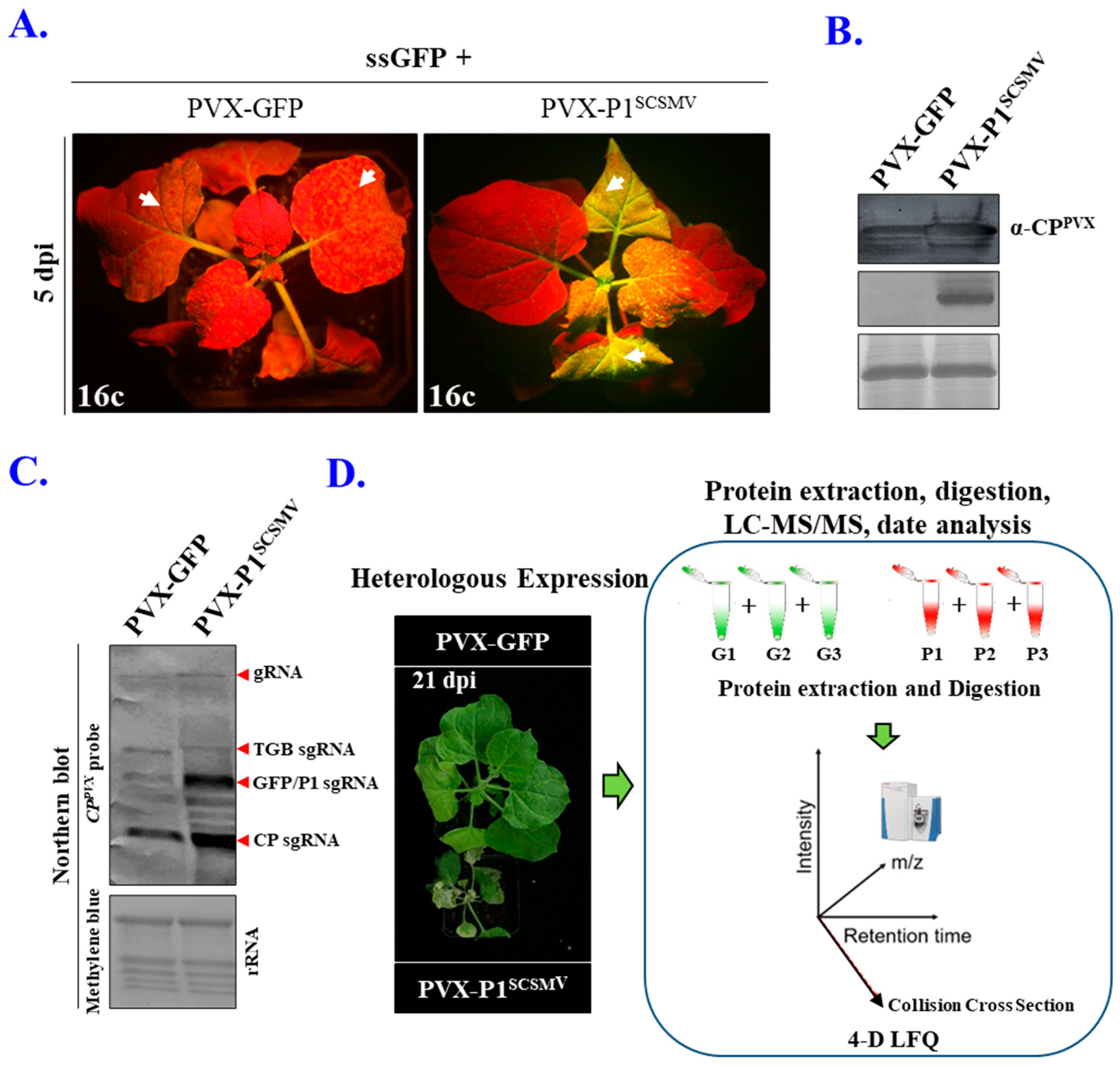{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viral-Derived Vector Construction
2.2. Plant Growth Conditions and Virus Inoculation
2.3. Antiserum Preparation
2.4. Measurement of the Photosynthesis
2.5. Agroinfiltration and GFP Imaging to Investigate the RNA Silencing Suppressor Activity
2.6. Western Blotting and Coomassie Brilliant Blue (CBB) Staining
2.7. Real-Time PCR Quantification of the Gene Expression
2.8. Northern Blotting and DIG-Labelled PVX Specific DNA Probe Preparation
2.9. Extraction of Total Proteins from Plant Tissue
2.10. Digestion, LC-MS/MS, Database Searching, and Bioinformatics Analysis
2.11. Generation of P1SCSMV Transgenic N. benthamiana Plants
2.12. Statistical Analysis
3. Results
3.1. Heterologous and Cis-Expression of P1SCSMV Enhances Viral Pathogenicity
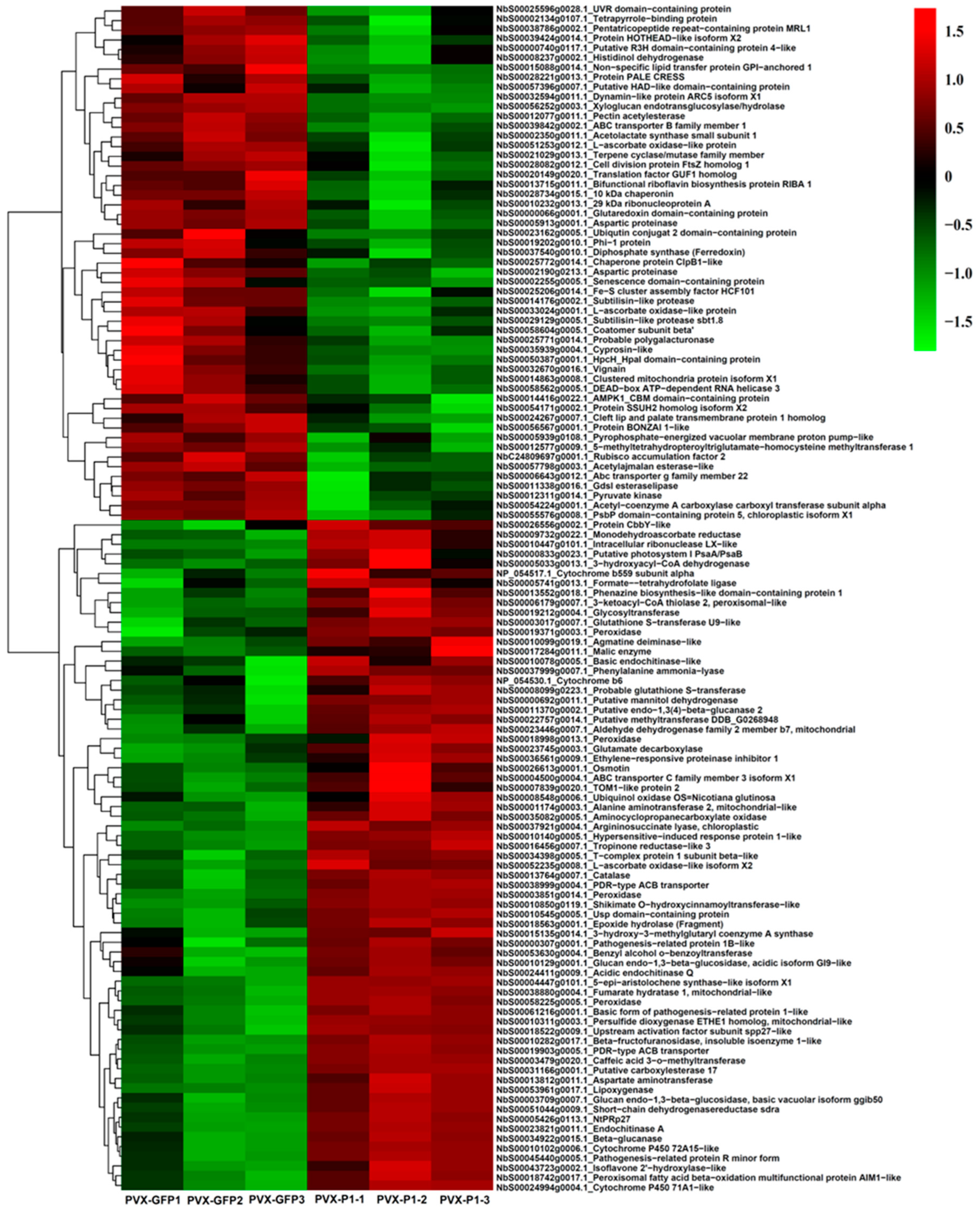
3.2. Identification, Quantification, and Analyses of DAPs after 4D Label-Free LC-MS/MS
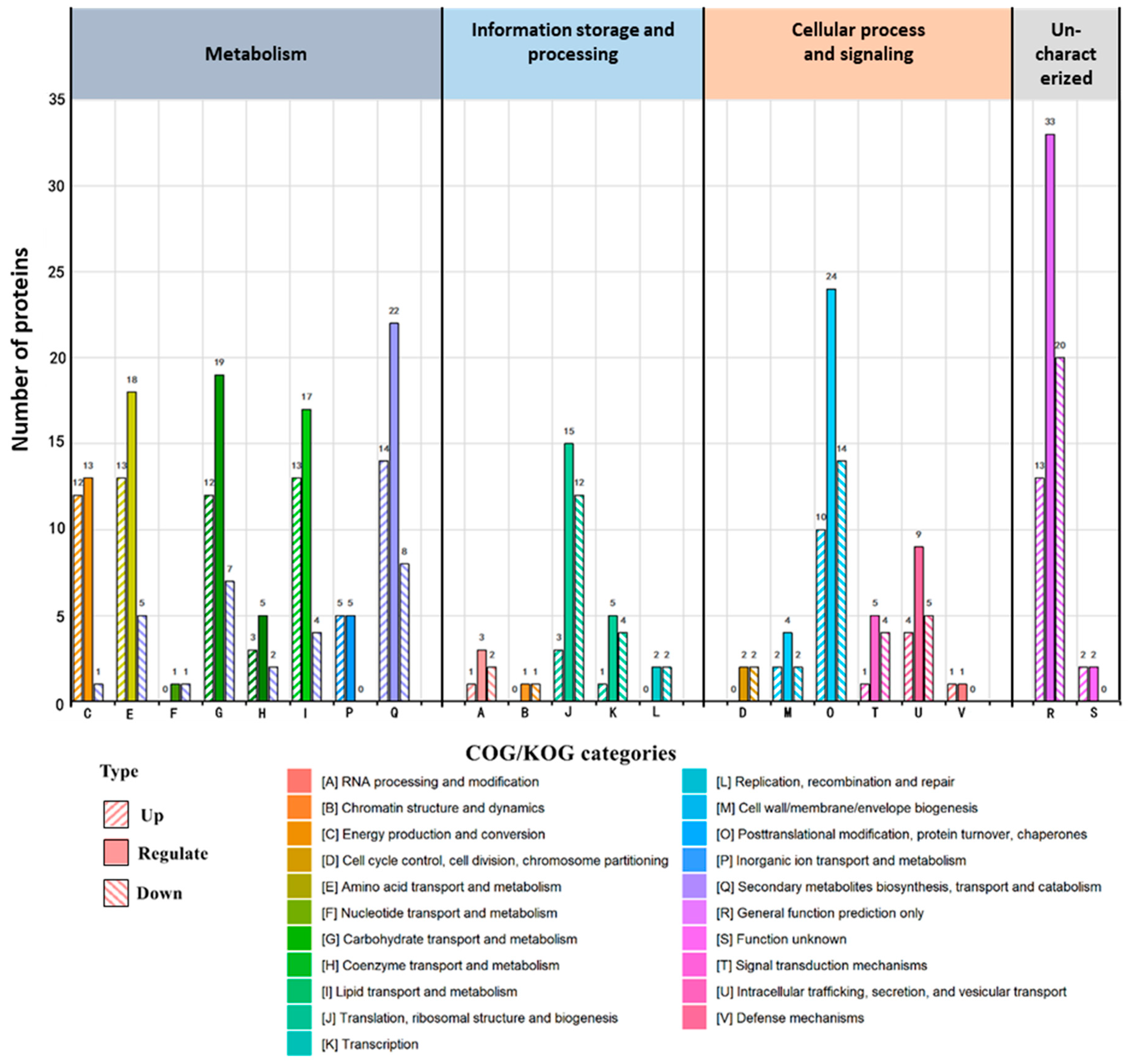
3.3. COG/KOG Annotation and KEGG Enrichment Analyses of DAPs
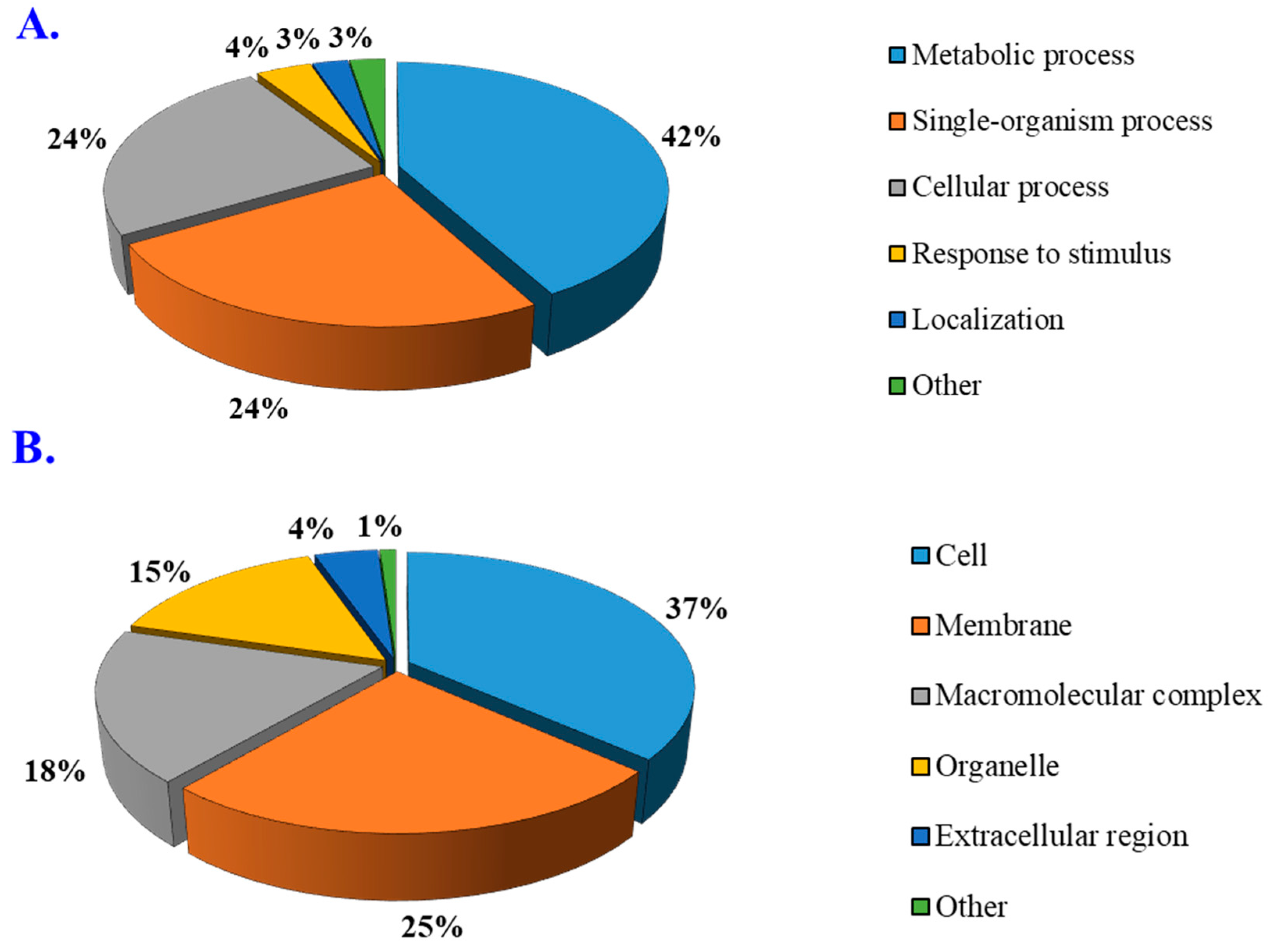
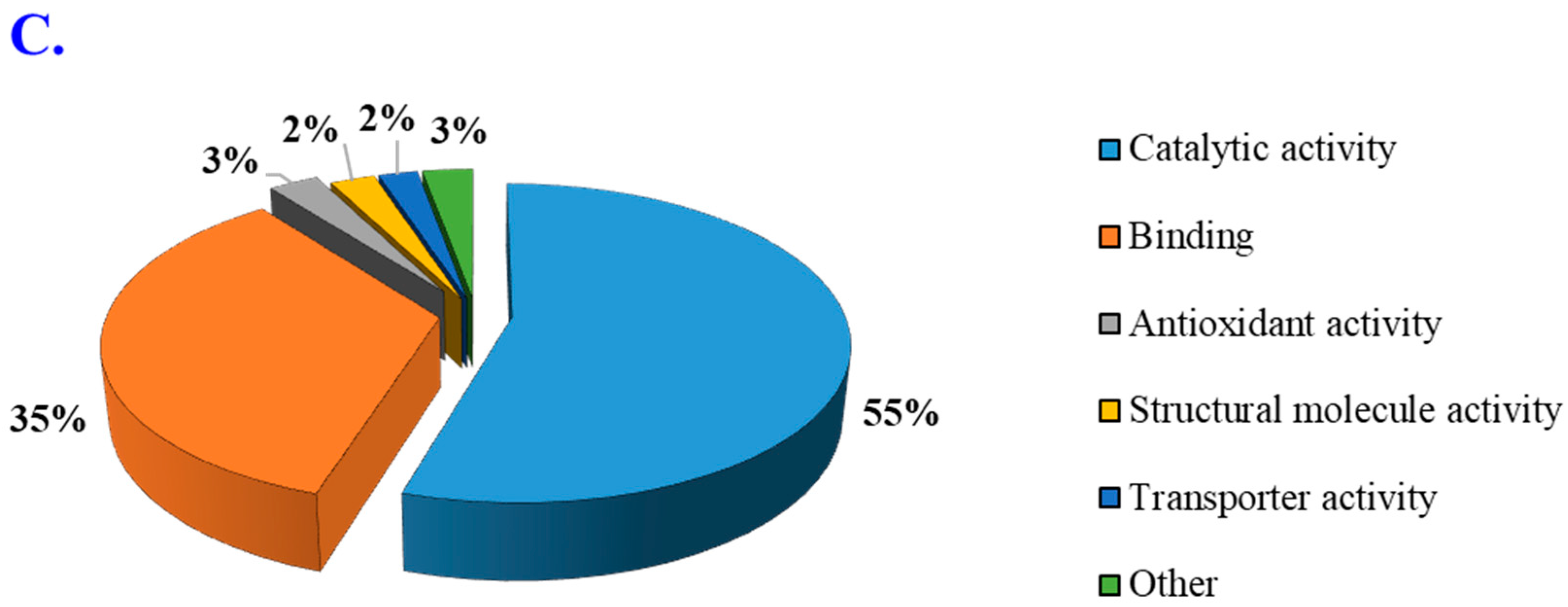
3.4. Gene Ontology Enrichment Analysis
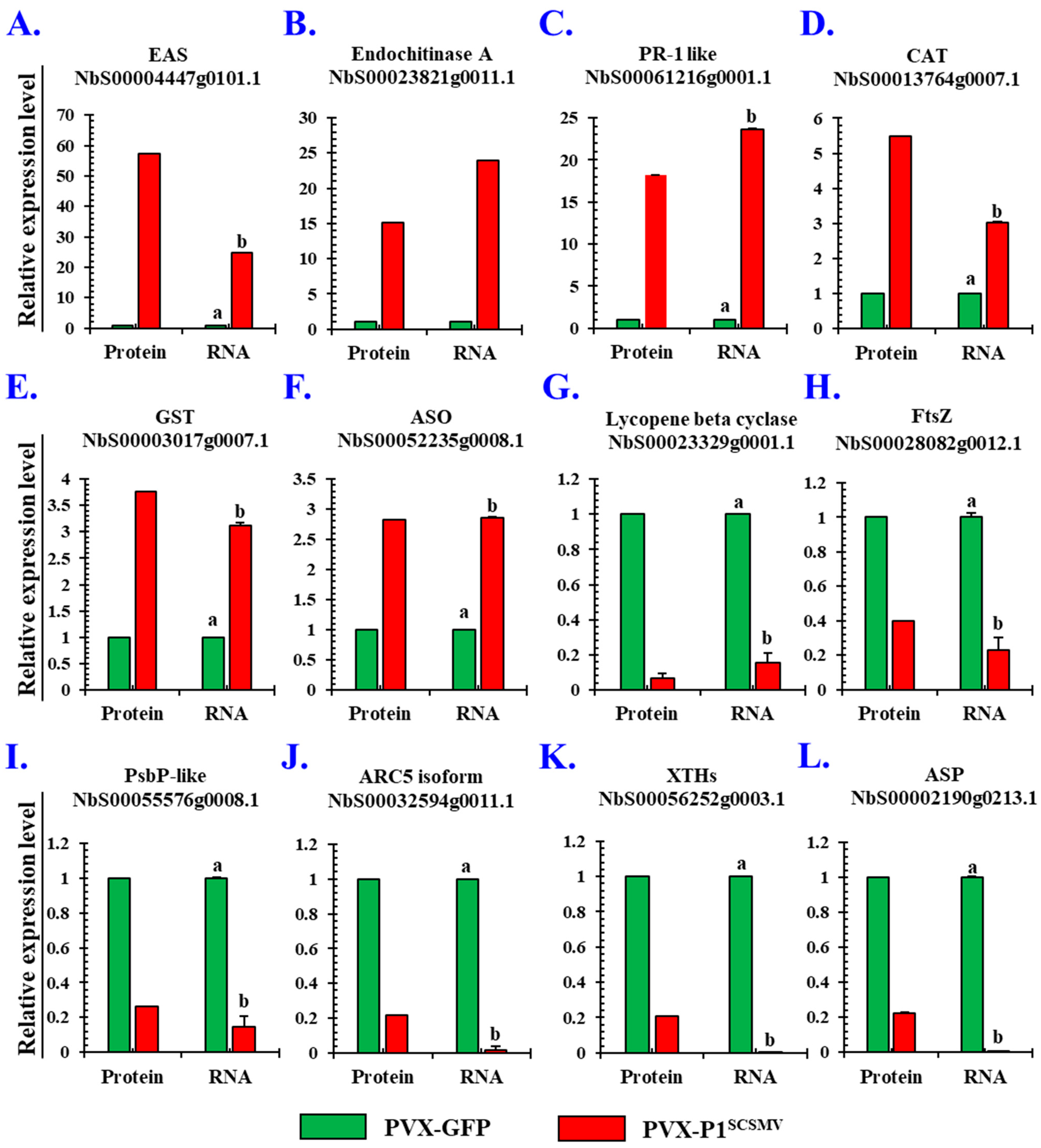
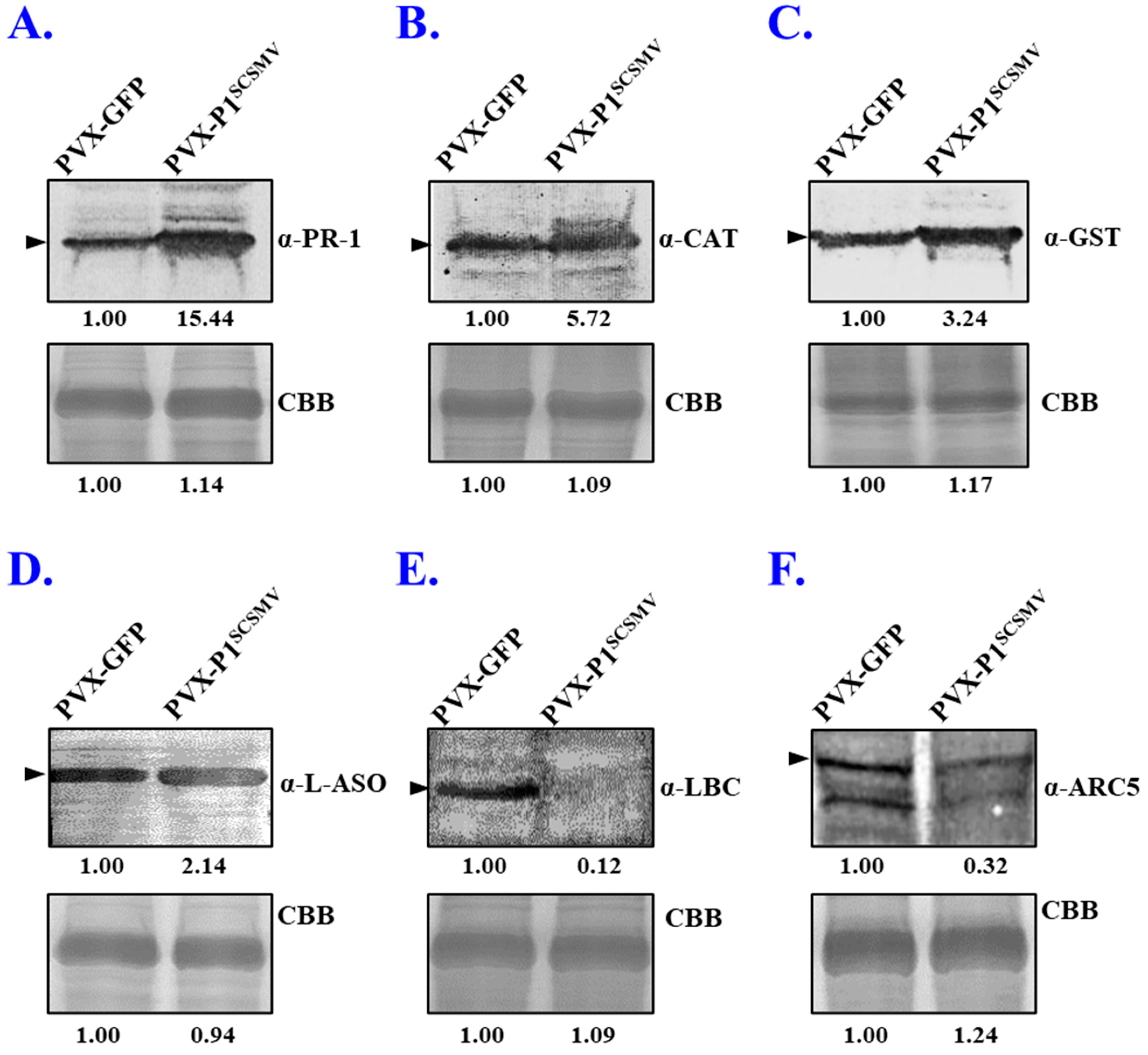
3.5. Validation of the Selected DAPs by qRT-PCR and Western Blotting
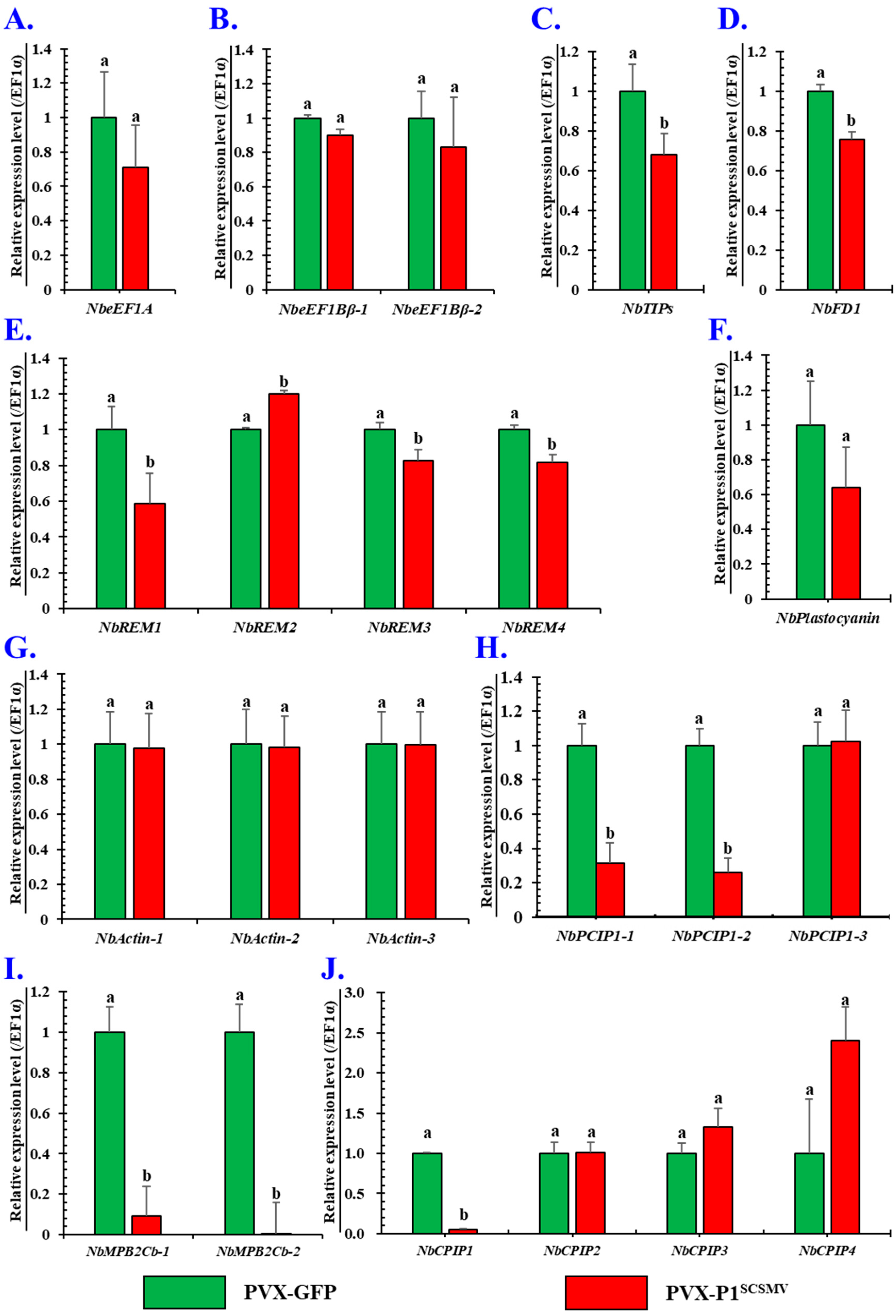
3.6. Effects of Is-Heterogeneous Expression of P1SCSMV on Intercellular Spread of PVX
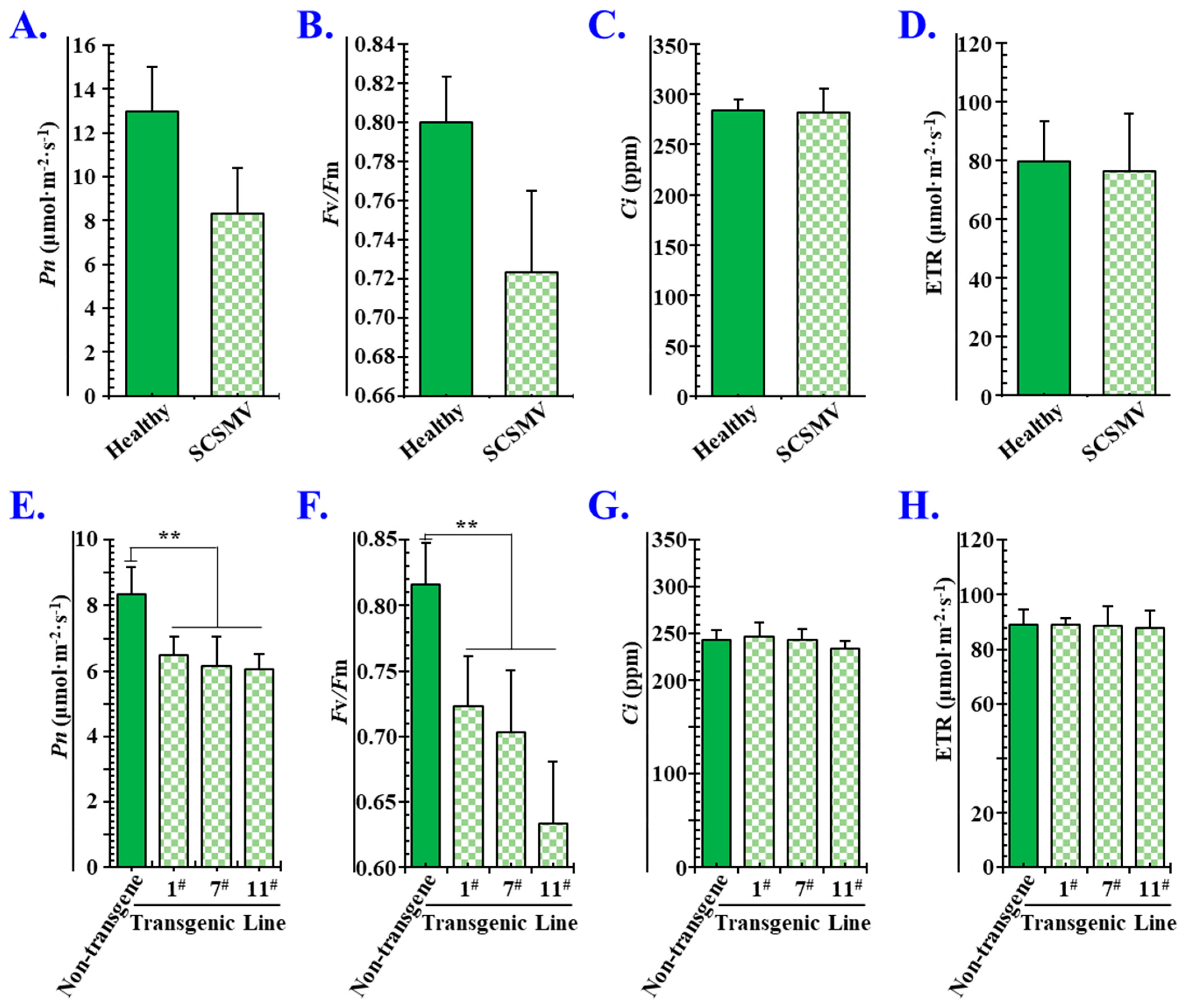
3.7. P1SCSMV Attenuates Photosynthesis by Damaging the PSII System
4. Discussion
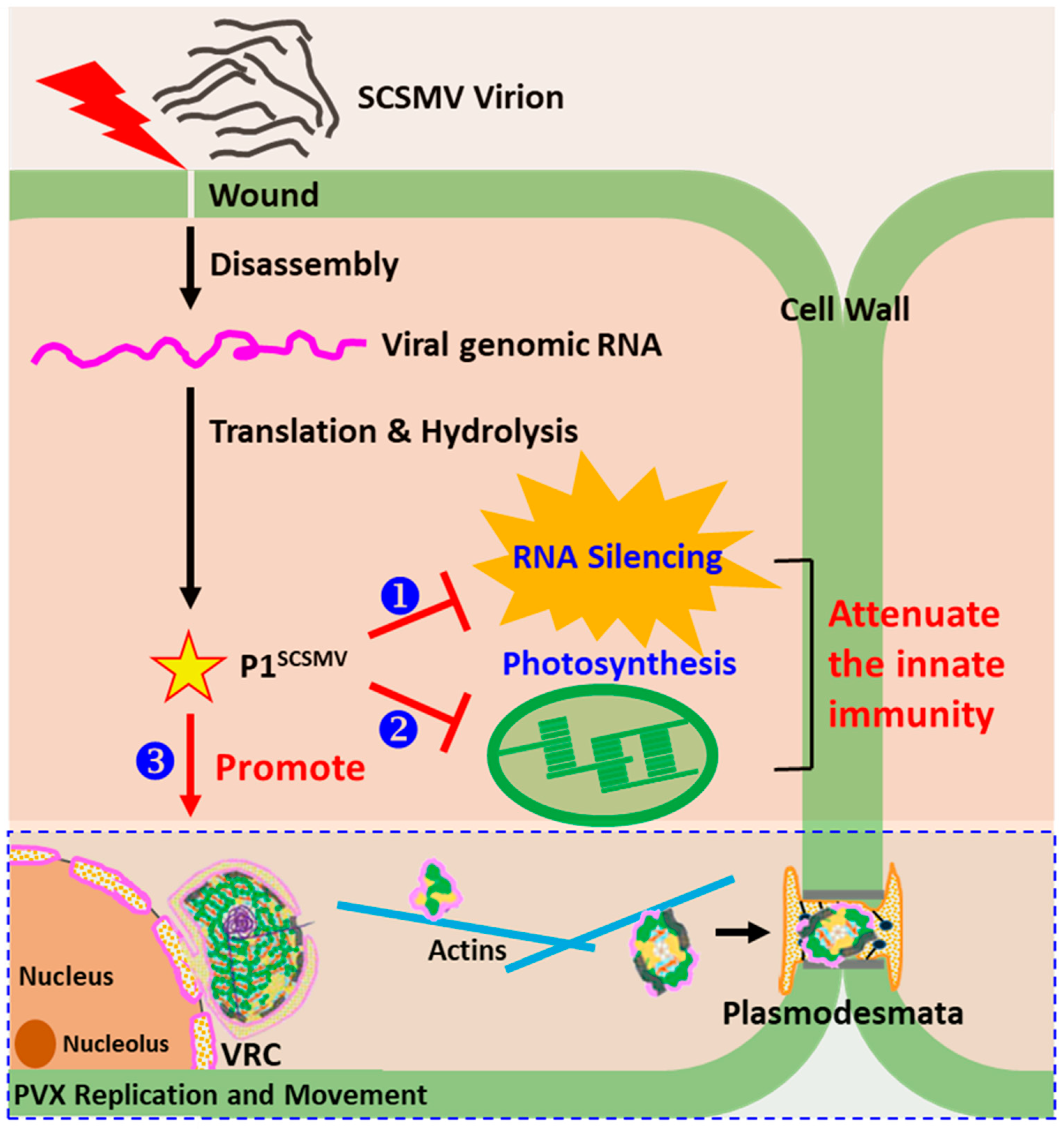
4.1. General Outcomes
4.2. P1SCSMV Protects the Viral Genomic RNA from Degradation by Host RNA Silencing
4.3. P1SCSMV Provides Better Cellular Conditions for PVX Accumulation
4.4. Overexpressing P1SCSMV Decreases Photosynthesis by Damaging the PSII System
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SCSMV | sugarcane streak mosaic virus |
| P1SCSMV | SCSMV-encoded protein 1 |
| DAPs | differential accumulated proteins |
| PVX | potato virus X |
| SMD | sugarcane mosaic disease |
| SBVD | sugarcane bacilliform virus disease |
| SCMV | sugarcane mosaic virus |
| SrMV | Sorghum mosaic virus |
| Knt | kilo-nucleotides |
| RSS | RNA silencing suppressor |
| TriMV | Triticum mosaic virus |
| VSR | viral encoded RNA silencing suppressor |
| 4D-proteomics | four-dimensional proteomics |
| CBB | coomassie brilliant blue |
| FC | fold change |
| NbEAS | 5-epi-aristolochene synthase-like isoform X1 |
| NbPR-1 | pathogenesis-related protein |
| NbXTH | xyloglucan endotransglucosylase/hydrolase |
| NbHSP | heat shock protein |
| NbARC5 | dynamin-like protein ARC5 |
| COG | clusters of orthologous groups |
| KEGG | Kyoto Encyclopedia of Genes and Genomes Pathways |
| GO | gene ontology |
| BP | biological process |
| CC | cellular components |
| MF | molecular functions |
| NbLBC | lycopene beta cyclase |
| NbFtsZ | cell division protein FtsZ homolog 1 |
| NbPsbP like | psbP domain-containing protein 5 |
| NbARC5i | dynamin-like protein ARC5 isoform |
| NbASP | aspartic proteinase |
| NbXTH | Xyloglucan endotransglucosylase |
| NbeEF1A | eukaryotic translation elongation factor 1A |
| NbeEF1Bβ1 | plant translation elongation factor |
| NbCPIPs | CP interacting proteins |
| MPB2C | TMV-MP30 binding protein 2C |
| NbFD1 | ferredoxin 1 |
| NbREMs | remorin |
| LHC | light-harvesting chlorophyll protein complex |
References
- Wang, K.L.; Deng, Q.Q.; Chen, J.W.; Shen, W.K. Development of a Reverse Transcription Loop-Mediated Isothermal Amplification Assay for Rapid and Visual Detection of Sugarcane Streak Mosaic Virus in Sugarcane. Crop Prot. 2019, 119, 38–45. [Google Scholar] [CrossRef]
- Vega, J.; Scagliusi, S.M.M.; Ulian, E.C. Sugarcane Yellow Leaf Disease in Brazil: Evidence of Association with a Luteovirus. Plant Dis. 1997, 81, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.L.; Park, J.W.; Mirkov, T.E.; Zhou, G.H. Viruses Causing Mosaic Disease in Sugarcane and Their Genetic Diversity in Southern China. Arch. Virol. 2008, 153, 1031–1039. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, K.; Sun, S.R.; Chen, J.L.; Huang, M.T.; Fu, H.Y.; Gao, S.J. Presence of Diverse Sugarcane Bacilliform Viruses Infecting Sugarcane in China Revealed by Pairwise Sequence Comparisons and Phylogenetic Analysis. Plant Pathol. J. 2019, 35, 41–50. [Google Scholar] [CrossRef]
- Viswanathan, R.; Rao, G.P. Disease Scenario and Management of Major Sugarcane Diseases in India. Sugar Tech 2011, 13, 336–353. [Google Scholar] [CrossRef]
- Li, W.; He, Z.; Li, S.; Huang, Y.; Zhang, Z.; Jiang, D.; Wang, X.; Luo, Z. Molecular Characterization of a New Strain of Sugarcane Streak Mosaic Virus (SCSMV). Arch. Virol. 2011, 156, 2101–2104. [Google Scholar] [CrossRef]
- King, A.M.Q.; Adams, M.J.; Eric, B.; Lefkowitz, E.J. Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier: San Diego, CA, USA, 2011; p. 1338. [Google Scholar]
- Putra, L.K.; Kristini, A.; Achadian, E.M.; Damayanti, T.A. Sugarcane streak mosaic virus in Indonesia: Distribution, Characterisation, Yield Losses and Management Approaches. Sugar Tech 2014, 16, 392–399. [Google Scholar] [CrossRef]
- Tatineni, S.; Qu, F.; Li, R.; Morris, T.J.; French, R. Triticum mosaic poacevirus enlists P1 rather than HC-Pro to suppress RNA silencing-mediated host defense. Virology 2012, 433, 104–115. [Google Scholar] [CrossRef]
- Chen, J.R.; Liang, S.S.; Sun, S.R.; Damaj, M.B.; Fu, H.Y.; Gao, S.J. Diverse Conserved Domains and A Positively Selected Site in the Sugarcane Streak Mosaic Virus P1 Protein Are Essential for RNA Silencing Suppression and Protein Stability. Plant Pathog. 2020, 69, 1390–1400. [Google Scholar] [CrossRef]
- Gupta, A.K.; Tatineni, S. RNA silencing suppression mechanisms of Triticum mosaic virus P1: dsRNA binding property and mapping functional motifs. Virus Res. 2019, 269, 197640. [Google Scholar] [CrossRef]
- Wu, X.; Valli, A.; García, J.A.; Zhou, X.; Cheng, X. The Tug-of-War between Plants and Viruses: Great Progress and Many Remaining Questions. Viruses 2019, 11, 203. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Dong, K.; Xu, K.; Zhang, K.; Jin, X.; Yang, M.; Zhang, Y.; Wang, X.; Han, C.; Yu, J.; et al. Barley Stripe Mosaic Virus Infection Requires PKA-Mediated Phosphorylation of γb for suppression of both RNA silencing and the host cell death response. New Phytol. 2018, 218, 1570–1585. [Google Scholar] [CrossRef] [PubMed]
- Pertermann, R.; Tamilarasan, S.; Gursinsky, T.; Gambino, G.; Schuck, J.; Weinholdt, C.; Lilie, H.; Grosse, I.; Golbik, R.P.; Pantaleo, V.; et al. A Viral Suppressor Modulates the Plant Immune Response Early in Infection by Regulating MicroRNA Activity. mBio 2018, 9, e00419-18. [Google Scholar] [CrossRef]
- Prianichnikov, N.; Koch, H.; Koch, S.; Lubeck, M.; Heilig, R.; Brehmer, S.; Fischer, R.; Cox, J. MaxQuant Software for Ion Mobility Enhanced Shotgun Proteomics. Mol. Cell. Proteom. 2020, 19, 1058–1069. [Google Scholar] [CrossRef]
- Meier, F.; Brunner, A.-D.; Koch, S.; Koch, H.; Lubeck, M.; Krause, M.; Goedecke, N.; Decker, J.; Kosinski, T.; Park, M.A.; et al. Online Parallel Accumulation–Serial Fragmentation (PASEF) with a Novel Trapped Ion Mobility Mass Spectrometer. Mol. Cell. Proteom. 2018, 17, 2534–2545. [Google Scholar] [CrossRef] [PubMed]
- Pang, H.; Li, W.; Zhang, W.; Zhou, S.; Hoare, R.; Monaghan, S.J.; Jian, J.; Lin, X. Acetylome profiling of Vibrio alginolyticus reveals its role in bacterial virulence. J. Proteom. 2020, 211, 103543. [Google Scholar] [CrossRef]
- Yu, H.; Bu, C.; Liu, Y.; Gong, T.; Liu, X.; Liu, S.; Peng, X.; Zhang, W.; Peng, Y.; Yang, J.; et al. Global crotonylome reveals CDYL-regulated RPA1 crotonylation in homologous recombination–mediated DNA repair. Sci. Adv. 2020, 6, eaay4697. [Google Scholar] [CrossRef]
- Yin, D.; Jiang, N.; Zhang, Y.; Wang, D.; Sang, X.; Feng, Y.; Chen, R.; Wang, X.; Yang, N.; Chen, Q. Global Lysine Crotonylation and 2-Hydroxyisobutyrylation in Phenotypically Different Toxoplasma gondii Parasites. Mol. Cell. Proteom. 2019, 18, 2207–2224. [Google Scholar] [CrossRef]
- Li, J.; Sun, Y.B.Y.; Chen, W.; Fan, J.; Li, S.; Qu, X.; Chen, Q.; Chen, R.; Zhu, D.; Zhang, J.; et al. Smad4 promotes diabetic nephropathy by modulating glycolysis and OXPHOS. EMBO Rep. 2020, 21, e48781. [Google Scholar] [CrossRef]
- Varela, A.L.N.; Oliveira, J.T.A.; Komatsu, S.; Silva, R.G.G.; Martins, T.F.; de Souza, P.F.N.; Lobo, A.K.M.; Vasconcelos, I.M.; Carvalho, F.E.L.; Silveira, J.A.G. A resistant cowpea (Vigna unguiculata [L.] Walp.) genotype became susceptible to cowpea severe mosaic virus (CPSMV) after exposure to salt stress. J. Proteom. 2019, 194, 200–217. [Google Scholar] [CrossRef]
- Dang, M.; Cheng, Q.; Hu, Y.; Wu, J.; Zhou, X.; Qian, Y. Proteomic Changes during MCMV Infection Revealed by iTRAQ Quantitative Proteomic Analysis in Maize. Int. J. Mol. Sci. 2019, 21, 35. [Google Scholar] [CrossRef]
- Rommens, C.M.; Salmeron, J.M.; Oldroyd, G.E.; Staskawicz, B.J. Use of a Gene Expression System Based on Potato Virus X to Rapidly Identify and Characterize a Tomato Pto Homolog That Controls Fenthion Sensitivity. Plant Cell 1995, 7, 249–257. [Google Scholar] [PubMed]
- Chapman, S.; Kavanagh, T.; Baulcombe, D. Potato Virus X as a Vector for Gene Expression in Plants. Plant J. 1992, 2, 549–557. [Google Scholar] [PubMed]
- Zhang, L.; Wang, Z.; Wang, X.; Li, D.; Han, C.; Zhai, Y.; Yu, J. Two Virus-Encoded RNA Silencing Suppressors, P14 OfBeet Necrotic Yellow Vein Virus and S6 of Rice Black Streak Dwarf Virus. Chin. Sci. Bull. 2005, 50, 305–310. [Google Scholar]
- Chen, H.; Cao, Y.; Li, Y.; Xia, Z.; Xie, J.; Carr, J.P.; Wu, B.; Fan, Z.; Zhou, T. Identification of differentially regulated maize proteins conditioning Sugarcane mosaic virus systemic infection. New Phytol. 2017, 215, 1156–1172. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Pan, X.; Wang, S.; Zhang, D. Cu2+ Inhibits Photosystem II Activities but Enhances Photosystem I Quantum Yield of Microcystis aeruginosa. Biol. Trace Elem. Res. 2014, 160, 268–275. [Google Scholar] [CrossRef]
- Hofgen, R.; Willmitzer, L. Storage of Competent Cells for Agrobacterium Transformation. Nucleic Acids Res. 1988, 16, 9877. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Yang, M.; Liu, S.; Li, Z.; Wang, X.; Han, C.; Yu, J.; Li, D. The Barley stripe mosaic virus γb protein promotes chloroplast-targeted replication by enhancing unwinding of RNA duplexes. PLoS Pathog. 2017, 13, e1006319. [Google Scholar] [CrossRef]
- Zhang, K.; Niu, S.; Di, D.; Shi, L.; Liu, D.; Cao, X.; Miao, H.; Wang, X.; Han, C.; Yu, J.; et al. Selection of reference genes for gene expression studies in virus-infected monocots using quantitative real-time PCR. J. Biotechnol. 2013, 168, 7–14. [Google Scholar] [CrossRef]
- Horsch, R.B.; Fry, J.; Hoffmann, N.; Neidermeyer, J.; Rogers, S.G.; Fraley, R.T. Leaf Disc Transformation. In Plant Molecular Biology Manual; Springer: Dordrecht, The Netherlands, 1989; pp. 63–71. [Google Scholar]
- Li, Y.; Sun, Q.; Zhao, T.; Xiang, H.; Zhang, X.; Wu, Z.; Zhou, C.; Zhang, X.; Wang, Y.; Zhang, Y.; et al. Interaction between Brassica yellows virus silencing suppressor P0 and plant SKP1 facilitates stability of P0 in vivo against degradation by proteasome and autophagy pathways. New Phytol. 2019, 222, 1458–1473. [Google Scholar] [CrossRef]
- Krenz, B.; Deuschle, K.; Deigner, T.; Unseld, S.; Kepp, G.; Wege, C.; Kleinow, T.; Jeske, H. Early Function of the Abutilon Mosaic Virus AC2 Gene as a Replication Brake. J. Virol. 2015, 89, 3683–3699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Pogany, J.; Panavas, T.; Xu, K.; Esposito, A.M.; Kinzy, T.G.; Nagy, P.D. Translation elongation factor 1A is a component of the tombusvirus replicase complex and affects the stability of the p33 replication co-factor. Virology 2009, 385, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.; Lee, S.; Lee, J.-H.; Kang, W.-H.; Kang, J.-H.; Kang, M.-Y.; Oh, C.-S.; Kang, B.-C. Plant Translation Elongation Factor 1Bβ Facilitates Potato Virus X (PVX) Infection and Interacts with PVX Triple Gene Block Protein 1. PLoS ONE 2015, 10, e0128014. [Google Scholar] [CrossRef]
- Fridborg, I.; Grainger, J.; Page, A.; Coleman, M.; Findlay, K.; Angell, S. TIP, A Novel Host Factor Linking Callose Degradation with the Cell-to-Cell Movement of Potato Virus X. Mol. Plant-Microbe Interact. 2003, 16, 132–140. [Google Scholar] [CrossRef]
- Yang, X.; Lu, Y.; Wang, F.; Chen, Y.; Tian, Y.; Jiang, L.; Peng, J.; Zheng, H.; Lin, L.; Yan, C.; et al. Involvement of the chloroplast gene ferredoxin 1 in multiple responses of Nicotiana benthamiana to Potato virus X infection. J. Exp. Bot. 2020, 71, 2142–2156. [Google Scholar] [CrossRef] [PubMed]
- Raffaele, S.; Bayer, E.; Lafarge, D.; Cluzet, S.; Retana, S.G.; Boubekeur, T.; Leborgne-Castel, N.; Carde, J.-P.; Lherminier, J.; Noirot, E.; et al. Remorin, a Solanaceae Protein Resident in Membrane Rafts and Plasmodesmata, Impairs Potato virus X Movement. Plant Cell 2009, 21, 1541–1555. [Google Scholar] [CrossRef]
- Qiao, Y.; Li, H.F.; Wong, S.M.; Fan, Z.F. Plastocyanin Transit Peptide Interacts with Potato virus X Coat Protein, While Silencing of Plastocyanin Reduces Coat Protein Accumulation in Chloroplasts and Symptom Severity in Host Plants. Mol. Plant-Microbe Interact. 2009, 22, 1523–1534. [Google Scholar] [CrossRef]
- Tilsner, J.; Linnik, O.; Wright, K.M.; Bell, K.; Roberts, A.G.; Lacomme, C.; Santa Cruz, S.; Oparka, K.J. The TGB1 Movement Protein of Potato Virus X Reorganizes Actin and Endomembranes into the X-Body, a Viral Replication Factory. Plant Physiol. 2012, 158, 1359–1370. [Google Scholar] [CrossRef] [PubMed]
- Park, M.-R.; Park, S.-H.; Cho, S.-Y.; Kim, K.-H. Nicotiana benthamiana protein, NbPCIP1, interacting with Potato virus X coat protein plays a role as susceptible factor for viral infection. Virology 2009, 386, 257–269. [Google Scholar] [CrossRef]
- Cho, S.-Y.; Cho, W.K.; Choi, H.-S.; Kim, K.-H. Cis-Acting Element (SL1) of Potato Virus X Controls Viral Movement by Interacting with the NbMPB2Cb and Viral Proteins. Virology 2012, 427, 166–176. [Google Scholar] [CrossRef]
- Choi, H.; Cho, W.K.; Kim, K.-H. Two homologous host proteins interact with potato virus X RNAs and CPs and affect viral replication and movement. Sci. Rep. 2016, 6, 28743. [Google Scholar] [CrossRef] [PubMed]
- Sasvari, Z.; Izotova, L.; Kinzy, T.G.; Nagy, P.D. Synergistic Roles of Eukaryotic Translation Elongation Factors 1Bγ and 1A in Stimulation of Tombusvirus Minus-Strand Synthesis. PLoS Pathog. 2011, 7, e1002438. [Google Scholar] [CrossRef]
- Hall, J.S.; Adams, B.; Parsons, T.J.; French, R.; Lane, L.C.; Jensen, S.G. Molecular Cloning, Sequencing, and Phylogenetic Relationships of a New Potyvirus: Sugarcane Streak Mosaic Virus, and a Reevaluation of the Classification of the Potyviridae. Mol. Phylogenet. Evol. 1998, 10, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Zhang, X.; Zhang, Q.; Ji, X.; Jia, Y.; Wang, H.; Niu, L.; Zhang, Y. Comparative transcriptome profiling uncovers a Lilium regale NAC transcription factor, LrNAC35, contributing to defence response against cucumber mosaic virus and tobacco mosaic virus. Mol. Plant Pathol. 2019, 20, 1662–1681. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, G.; Zhang, Y.; Song, Q.; Chen, Z.; Wang, J.; Guo, J.; Niu, N.; Wang, J.; Ma, S. Comparative Studies of Mitochondrial Proteomics Reveal an Intimate Protein Network of Male Sterility in Wheat (Triticum aestivum L.). J. Exp. Bot. 2015, 66, 6191–6203. [Google Scholar] [CrossRef]
- Valentim-Neto, P.A.; Rossi, G.B.; Anacleto, K.B.; de Mello, C.S.; Balsamo, G.M.; Arisi, A.C.M. Leaf proteome comparison of two GM common bean varieties and their non-GM counterparts by principal component analysis. J. Sci. Food Agric. 2016, 96, 927–932. [Google Scholar] [CrossRef]
- Stare, T.; Stare, K.; Weckwerth, W.; Wienkoop, S.; Gruden, K. Comparison between Proteome and Transcriptome Response in Potato (Solanum tuberosum L.) Leaves Following Potato Virus Y (PVY) Infection. Proteomes 2017, 5, 14. [Google Scholar] [CrossRef]
- Di Carli, M.; Villani, M.E.; Bianco, L.; Lombardi, R.; Perrotta, G.; Benvenuto, E.; Donini, M. Proteomic Analysis of the Plant−Virus Interaction in Cucumber Mosaic Virus (CMV) Resistant Transgenic Tomato. J. Proteome Res. 2010, 9, 5684–5697. [Google Scholar] [CrossRef]
- Varela, A.L.N.; Komatsu, S.; Wang, X.; Silva, R.G.G.; Souza, P.F.N.; Lobo, A.K.M.; Vasconcelos, I.M.; Silveira, J.A.G.; Oliveira, J.T.A. Gel-Free/Label-Free Proteomic, Photosynthetic, and Biochemical Analysis of Cowpea (Vigna unguiculata [L.] Walp.) Resistance against Cowpea Severe Mosaic Virus (CPSMV). J. Proteom. 2017, 163, 76–91. [Google Scholar] [CrossRef]
- Zhong, X.; Wang, Z.Q.; Xiao, R.; Wang, Y.; Xie, Y.; Zhou, X. iTRAQ analysis of the tobacco leaf proteome reveals that RNA-directed DNA methylation (RdDM) has important roles in defense against geminivirus-betasatellite infection. J. Proteom. 2017, 152, 88–101. [Google Scholar] [CrossRef]
- Meier, F.; Brunner, A.-D.; Frank, M.; Ha, A.; Bludau, I.; Voytik, E.; Kaspar-Schoenefeld, S.; Lubeck, M.; Raether, O.; Bache, N.; et al. diaPASEF: Parallel accumulation–serial fragmentation combined with data-independent acquisition. Nat. Methods 2020, 17, 1229–1236. [Google Scholar] [CrossRef] [PubMed]
- Merai, Z.; Kerényi, Z.; Kertész, S.; Magna, M.; Lakatos, L.; Silhavy, D. Double-Stranded RNA Binding May Be a General Plant RNA Viral Strategy to Suppress RNA Silencing. J. Virol. 2006, 80, 5747–5756. [Google Scholar] [CrossRef] [PubMed]
- Burgyán, J.; Havelda, Z. Viral suppressors of RNA silencing. Trends Plant Sci. 2011, 16, 265–272. [Google Scholar] [CrossRef]
- Ruiz-Ferrer, V.; Voinnet, O. Roles of Plant Small RNAs in Biotic Stress Responses. Annu. Rev. Plant Biol. 2009, 60, 485–510. [Google Scholar] [CrossRef]
- Gupta, A.K.; Tatineni, S. Wheat streak mosaic virus P1 Binds to dsRNAs without Size and Sequence Specificity and a GW Motif Is Crucial for Suppression of RNA Silencing. Viruses 2019, 11, 472. [Google Scholar] [CrossRef] [PubMed]
- Young, B.A.; Stenger, D.C.; Qu, F.; Morris, T.J.; Tatineni, S.; French, R. Tritimovirus P1 functions as a suppressor of RNA silencing and an enhancer of disease symptoms. Virus Res. 2012, 163, 672–677. [Google Scholar] [CrossRef]
- Vance, V.B.; Berger, P.H.; Carrington, J.C.; Hunt, A.G.; Shi, X.M. 5′ Proximal potyviral sequences mediate potato virus X/potyviral synergistic disease in transgenic tobacco. Virology 1995, 206, 583–590. [Google Scholar] [CrossRef]
- Gonzalez-Jara, P.; Tenllado, F.; Martinez-Garcia, B.; Atencio, F.A.; Barajas, D.; Vargas, M.; Díaz-Ruiz, J.R. Host-dependent differences during synergistic infection by Potyviruses with potato virus X. Mol. Plant Pathol. 2004, 5, 29–35. [Google Scholar] [CrossRef]
- Aguilar, E.; Almendral, D.; Allende, L.; Pacheco, R.; Chung, B.N.; Canto, T.; Tenllado, F. The P25 Protein of Potato Virus X (PVX) Is the Main Pathogenicity Determinant Responsible for Systemic Necrosis in PVX-Associated Synergisms. J. Virol. 2015, 89, 2090–2103. [Google Scholar] [CrossRef]
- De, S.; Chavez-Calvillo, G.; Wahlsten, M.; Mäkinen, K. Disruption of the methionine cycle and reduced cellular gluthathione levels underlie potex-potyvirus synergism in Nicotiana benthamiana. Mol. Plant Pathol. 2018, 19, 1820–1835. [Google Scholar] [CrossRef]
- Hofius, D.; Maier, A.T.; Dietrich, C.; Jungkunz, I.; Bornke, F.; Maiss, E.; Sonnewald, U. Capsid Protein-Mediated Recruitment of Host DnaJ-Like Proteins Is Required for Potato Virus Y Infection in Tobacco Plants. J. Virol. 2007, 81, 11870–11880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano, I.; Audran, C.; Rivas, S. Chloroplasts at work during plant innate immunity. J. Exp. Bot. 2016, 67, 3845–3854. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Lee, K.P.; Dogra, V.; Zhang, S.; Liu, K.; Caceres-Moreno, C.; Lv, S.; Xing, W.; Kato, Y.; Sakamoto, W.; et al. Impaired PSII Proteostasis Promotes Retrograde Signaling via Salicylic Acid. Plant Physiol. 2019, 180, 2182–2197. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.-D.; Cho, Y.-H.; Yoo, S.-D. Phytohormone ethylene-responsive Arabidopsis organ growth under light is in the fine regulation of Photosystem II deficiency-inducible AKIN10 expression. Sci. Rep. 2017, 7, 2767. [Google Scholar] [CrossRef] [Green Version]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, K.; Xu, X.; Guo, X.; Ding, S.; Gu, T.; Qin, L.; He, Z. Sugarcane Streak Mosaic Virus P1 Attenuates Plant Antiviral Immunity and Enhances Potato Virus X Infection in Nicotiana benthamiana. Cells 2022, 11, 2870. https://doi.org/10.3390/cells11182870
Zhang K, Xu X, Guo X, Ding S, Gu T, Qin L, He Z. Sugarcane Streak Mosaic Virus P1 Attenuates Plant Antiviral Immunity and Enhances Potato Virus X Infection in Nicotiana benthamiana. Cells. 2022; 11(18):2870. https://doi.org/10.3390/cells11182870
Chicago/Turabian StyleZhang, Kun, Xiaowei Xu, Xiao Guo, Shiwen Ding, Tianxiao Gu, Lang Qin, and Zhen He. 2022. "Sugarcane Streak Mosaic Virus P1 Attenuates Plant Antiviral Immunity and Enhances Potato Virus X Infection in Nicotiana benthamiana" Cells 11, no. 18: 2870. https://doi.org/10.3390/cells11182870