2. Materials and Methods
2.1. Isolation of γδ T Cells and Culture of Fibroblasts
This study was approved by the Institutional Review Board of the Nagasaki University Hospital. Peripheral blood samples were obtained from healthy adult volunteers and patients with IPF. Written informed consent was obtained from all the participants. Heparin sodium (Mochida Pharmaceutical, Co., Ltd., Shinjuku-ku, Tokyo, Japan; 1/100 volume) was added to blood samples diluted with equal volumes of Dulbecco’s phosphate-buffered saline (PBS) (-) (Nissui Pharmaceutical Co., Ltd., Minato-ku, Tokyo, Japan). Diluted blood samples (20 mL) were loaded onto 20 mL Ficoll-PaqueTM PLUS (GE Healthcare BioSciences AB, Chicago, IL, USA) in 50 mL conical tubes (Corning Inc., Corning, NY, USA) and centrifuged at 600× g at 25 °C for 30 min. The fluffy layer was then transferred into a new 50 mL conical tube and diluted with 2.5 volumes of PBS (-). The peripheral blood mononuclear cell (PBMC) suspension was centrifuged at 900× g at 4 °C for 10 min. After the supernatant was aspirated, cell pellets were dispersed by tapping and then resuspended in 13 mL PBS (-). The cell suspension was centrifuged at 600× g at 4 °C for 5 min, and the supernatant was aspirated. Subsequently, the cell pellets were dispersed by tapping and resuspended in 7 mL of Yssel’s medium consisting of Iscove’s modified Dulbecco’s medium (Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% human AB serum (Cosmo Bio Co., Ltd., Koto-ku, Tokyo, Japan), 3.6 × 10−2 M NaHCO3 (Nacalai Tesque Inc., Nakagyo-ku, Kyoto, Japan), 3.3 × 10−5 M 2-aminoethanol (Nacalai Tesque Inc., Kyoto, Japan), 40 mg/L transferin apo form (Nacalai Tesque Inc., Kyoto, Japan), 5 mg/L human recombinant insulin (Merck & Co., Inc., Darmstadt, Hesse, Germany), 2 mg/L linoleic acid (Merck & Co., Inc., Darmstadt, Germany), 2 mg/L oleic acid (Merck & Co., Inc., Darmstadt, Germany), 2 mg/mL palmitic acid (Merck & Co., Inc., Darmstadt, Germany), 100 μg/mL streptomycin, and 100 U/mL penicillin. The PBMC suspension (1.5 mL each, 1–2.5 × 106 cells/mL) was dispensed into the wells of a 24-well plate (Corning Inc., Corning, NY, USA). A PTA stock solution (1.5 μL, 1 mM) in dimethyl sulfoxide at a final concentration of 1 μM was added to each well. The plate was incubated at 37 °C and 5% CO2 for 24 h. Interleukin-2 (IL-2, Shionogi Pharmaceutical Co., Ltd., Chuo-ku, Osaka, Japan) was added to each well every day from Day 1 to Day 5 to obtain a final concentration of 100 U/mL. On Day 6, 1.5 mL of Yssel’s medium was added to the wells, and the diluted cell suspension was mixed by pipetting. Half of the suspension was transferred into a new well, and 100 U/mL of IL-2 was added. On Day 7, 1.5 mL of complete RPMI 1640 medium (Merck & Co., Inc., Darmstadt, Germany) supplemented with 10% fetal calf serum (FCS, Merck & Co., Inc., Darmstadt, Germany), 10−5 M of 2-mercaptoethanol (Wako Pure Chemical Industries, Ltd., Osaka, Japan), 100 μg/mL streptomycin (Meiji Seika Pharma Co., Ltd., Tokyo, Japan), and 100 U/mL penicillin (Meiji Seika Pharma Co., Ltd.) was added to each well. Each cell suspension was split into two wells, and IL-2 was added at a final concentration of 100 U/mL. Vγ9Vδ2-positive γδ T cells were expanded using complete RPMI 1640 medium plus 100 U/mL of IL-2 by Day 11, harvested by centrifuging at 600× g at 4 °C for 5 min, resuspended in cryopreservation media, dispensed into cryovials, incubated overnight at −80 °C, and then stored in liquid nitrogen until use. Cells collected on Day 0 and Day 11 were stained with 3 μL of phycoerythrin (PE)-conjugated anti-CD3 mAb (BD Biosciences, San Diego, CA), NKG2D, DNAM-1, FasL, or tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) mAb (BioLegend, San Diego, CA), and FITC-conjugated anti-TCR Vδ2 mAb (BD Biosciences) on ice for 15 min in a 96-well round-bottom plate. After three washes using 200 μL PBS (-) supplemented with 2% FCS, the cells were resuspended in 200 μL of PBS (-)/2% FCS and analyzed using a FACSLyric flow cytometer (BD Biosciences). The cell population was visualized using FlowJo software ver. 10 (FlowJo LLC, Ashland, OR). Diseased human lung fibroblasts derived from a patient with IPF (DHLF, Lonza Walkersville, Inc., Basel, Switzerland) were cultured in complete RPMI 1640 medium in 75 cm2 flasks (Corning Inc., Corning, NY, USA) at 37 °C and 5% CO2. Assays were conducted using cells with low passage numbers (passages 3–7) to avoid altered morphologies, growth rates, and responses to stimuli.
2.2. Co-Culture of Lung Fibroblasts with γδ T Cells
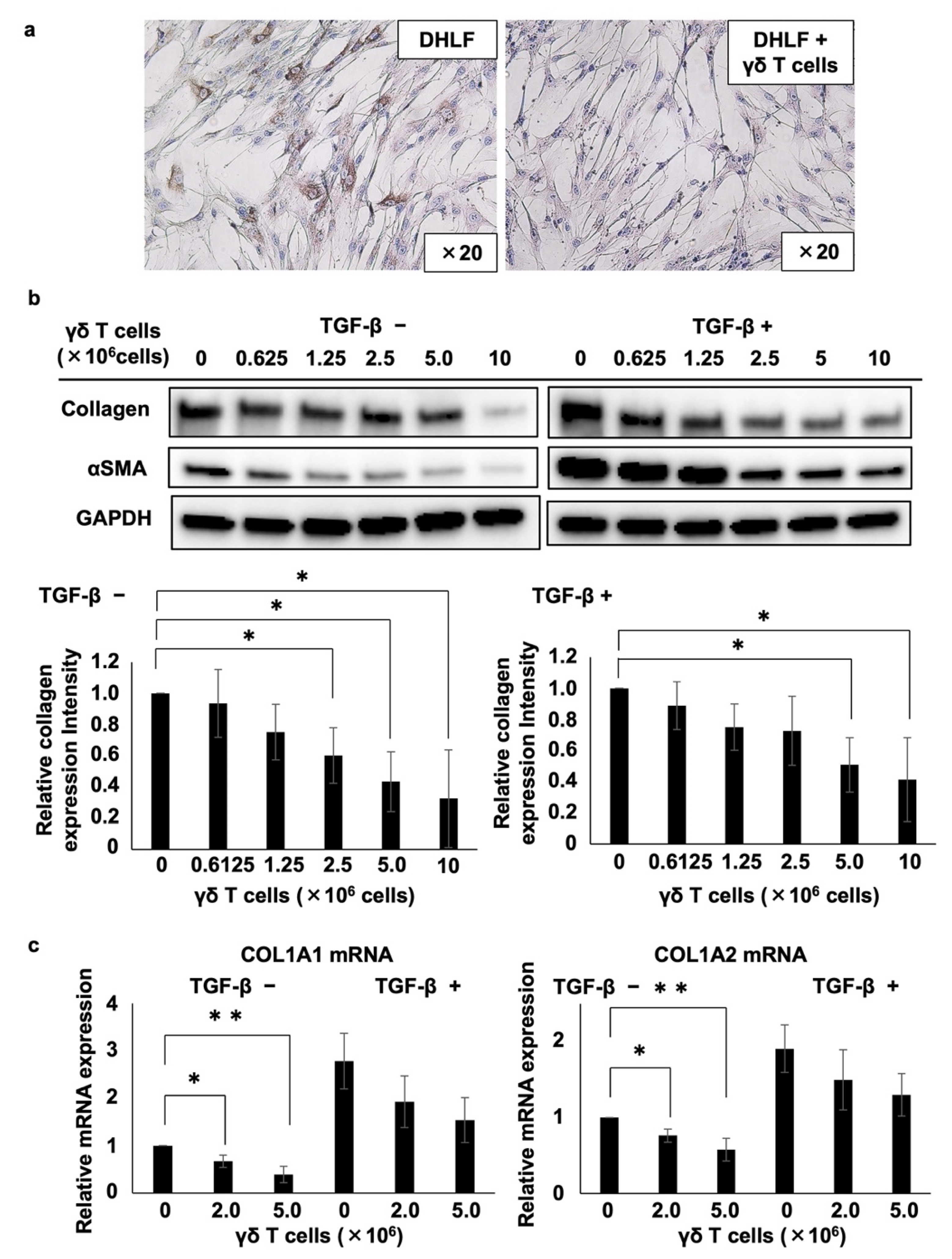
DHLF cells (2.5 × 104 cells in 1 mL of complete RPMI 1640 medium) were cultured overnight in a Lab-Tek chamber slide (Thermo Fisher Scientific, Waltham, MA, USA). After the supernatant was aspirated, 1 mL of thawed γδ T cells (2 × 106) or complete RPMI 1640 medium was added to the chamber slide, and the mixture was co-cultured at 37 °C and 5% CO2 for 2 days. Collagen type I in DHLF cells was analyzed via immunocytochemistry. Western blotting and real-time reverse transcription polymerase chain reaction (real-time RT-PCR) assays were performed to confirm the collagen expression levels. Briefly, the DHLF cells (1.5 × 105 cells in 2 mL of complete RPMI 1640 medium) were seeded in a 6-well plate (Corning Inc., Corning, NY, USA). Either 1 μL of complete RPMI 1640 medium or 1 μL of transforming growth factor-β1 (TGF-β) (final concentration: 5 ng/mL) was then added. The plate was incubated overnight at 37 °C and 5% CO2, the supernatant was aspirated, and 2 mL of PBS (-) was added to the well. After aspirating the supernatant, 2 mL of serially diluted PTA-expanded γδ T cells (6.25 × 105, 1.25 × 106, 2.5 × 106, 5 × 106, or 1 × 107 cells for Western blot assay; 2 × 106 or 5 × 106 cells for RT-PCR) or complete RPMI 1640 medium was added, and the plate was incubated for 2 days at 37 °C and 5% CO2. The supernatant was aspirated, and the cells were washed twice with 2 mL of PBS (-) to remove γδ T cells. For Western blot analysis, 100 μL of RIPA buffer containing 1% protease/phosphatase inhibitor (Thermo Fisher Scientific, Waltham, MA, USA) was added to each well, and the plate was then incubated on ice for 5 min. Cell lysates were transferred into tubes using a Coster cell scraper (Corning Inc., Corning, NY, USA) and centrifuged at 14,000× g for 15 min at 4 °C. The supernatants were then transferred into new tubes and stored at −80 ℃ until use. Collagen type I in DHLF cells was analyzed via Western blotting. To conduct real-time RT-PCR assays, cells were dissolved in 350 μL of lysis buffer (RLT lysis buffer containing 1% β-mercaptoethanol) using a Coster cell scraper (Corning Inc., Corning, NY, USA), transferred into safe-lock tubes (Eppendorf, Hamburg, Germany), and stored at −80 ℃ until use. Total RNA was prepared using an RNeasy Plus mini kit (QIAgen, Valencia, CA), and COL1A1 and COL1A2 mRNA expression was quantified using the standard protocol. To analyze γδ T cell cytotoxicity, DHLF cells (2 × 104) treated with TGF-β (5 ng/mL) or complete RPMI 1640 medium were cultured overnight in a 96-well flat-bottom plate (Corning Inc., Corning, NY, USA) at 37 ℃ and 5% CO2. The supernatants were then aspirated and the cells were washed with 200 μL of PBS (-). After removing the supernatant, 200 μL of γδ T cells at effector-to-target cell ratios (E/T ratios) of 0, 6.25:1, 12.5:1, 25:1, 50:1, and 100:1 were added into the wells, and the plate was incubated for 24 h at 37 ℃ and 5% CO2. The supernatants were then aspirated, and the cells were washed three times with 200 μL of complete RPMI 1640 medium. A total of 100 μL of CellTiter-Glo® reagent (PerkinElmer Inc., Ealtham, MA, USA) was subsequently added into each well, and cell lysates were transferred into a 96-well OptiPlate (PerkinElmer Inc.). Luminescence was measured using an ARVO multiplate reader (PerkinElmer Inc.).
2.3. Co-Culture of Lung Fibroblasts and γδ T Cells through a Membrane Insert
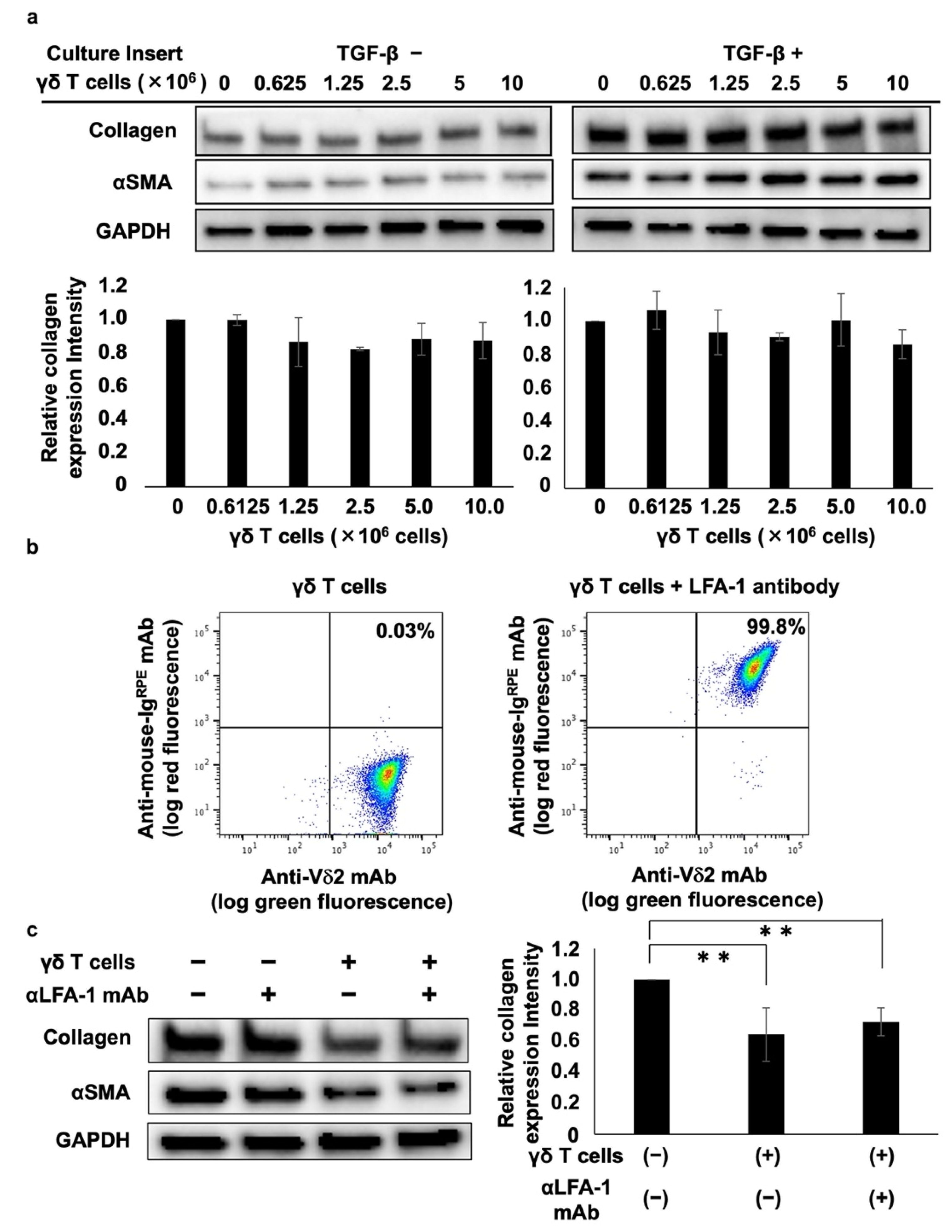
DHLF cells (1.5 × 105 cells in 2 mL complete RPMI 1640 medium) were placed in a 6-well plate, combined with either 1 μL of TGF-β (final concentration 5 ng/mL) or 1 μL of complete RPMI 1640 medium, and incubated overnight at 37 °C and 5% CO2. The supernatant was aspirated, and 2 mL of PBS (-) was added. Supernatants were then removed, and 1.5 mL of complete RPMI 1640 medium was added into the wells. Next, cell culture inserts (0.4 μm pore size; polyethylene terephthalate membrane, Corning Inc., Corning, NY, USA) were placed into the 6-well plate, and 1 mL of γδ T cells (6.25 × 105, 1.25 × 106, 2.5 × 106, 5.0 × 106, or 1.0 × 107 cells) or complete RPMI 1640 medium was added. The plate was incubated for 2 days at 37 °C and 5% CO2. After this, the cell culture inserts were removed, the supernatants were aspirated, and the cells were washed with PBS (-). Cell lysates were dissolved in RIPA buffer as described earlier, and collagen type I was analyzed using Western blot assay.
2.4. Flow Cytometric Analysis of LFA-1 Expression on γδ T Cells
γδ T cell suspensions were placed in a 96-well round-bottom plate and centrifuged at 600× g and 4 °C for 2 min. The supernatants were then aspirated, cell pellets were resuspended in 50 μL of PBS/2% FCS, and 2 μg/mL of mouse anti-human lymphocyte function-associated antigen-1 (LFA-1, or CD11a/CD18) mAb (BioXcell, West Lebanon, NH, USA) was added. The plate was placed on ice for 30 min, and the cells were washed three times with 200 μL of PBS/2% FCS and resuspended in 50 μL of PBS/2% FCS. The cells were subsequently placed on ice for 15 min and stained with fluorescein isothiocyanate (FITC)-conjugated anti-TCR Vδ2 mAb and R-phycoerythrin (RPE)-conjugated anti-mouse Ig (Agilent Technologies, Inc., Santa Clara, CA). The cells were then washed three times with 200 μL of PBS (-) and resuspended in 200 μL of PBS (-). Stained cells were analyzed using a FACSLyric flow cytometer, and the cell population was visualized using FlowJo software ver. 10.
2.5. Effect of Anti-LFA-1 mAb on Collagen Type I in Lung Fibroblasts Co-Cultured with γδ T Cells
DHLF cells (1.5 × 105) in 2 mL of complete RPMI 1640 medium were placed in a 6-well plate and incubated overnight at 37 °C and 5% CO2. The supernatant was aspirated, and 2 mL of γδ T cell suspension (5 × 106 cells) or complete RPMI 1640 medium was added. Anti-LFA1 antibody was added to obtain a final concentration of 2 μg/mL, and the plate was incubated for 2 days at 37 °C and 5% CO2. The cells were washed twice with 2 mL of PBS (-) and then dissolved in RIPA buffer, as described earlier. Collagen type I and αSMA protein expression levels in DHLF cells were analyzed via Western blot assays.
2.6. Co-Culturing Lung Fibroblasts and γδ T Cells Stimulated with HMBPP
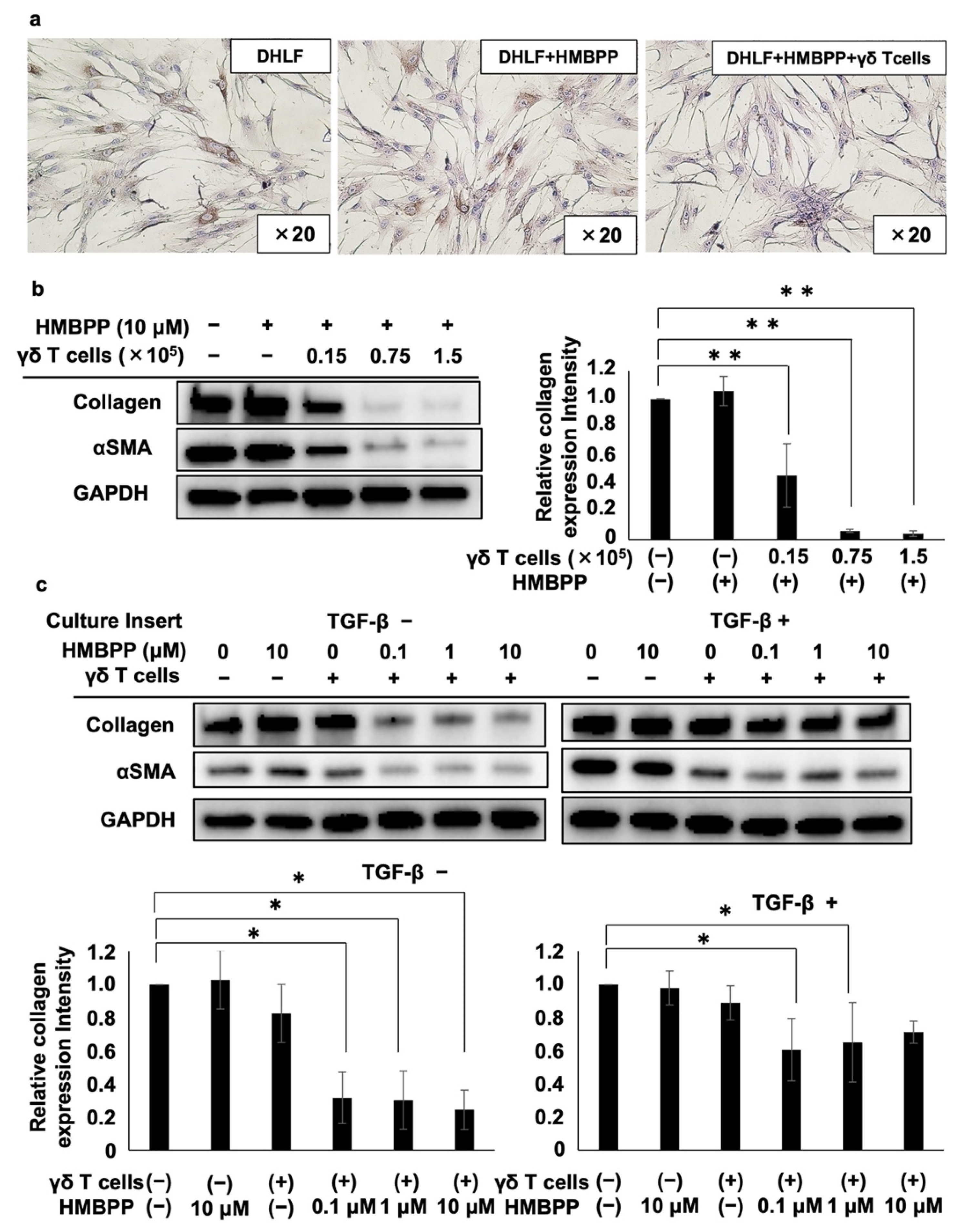
DHLF cells (2.5 × 104) were cultured in 1 mL of complete RPMI 1640 medium overnight in a Lab-Tek chamber slide at 37 ℃ and 5% CO2. The supernatant was removed, and 1 mL of γδ T cell suspension (5 × 103 cells) or complete RPMI 1640 medium was added to the chamber slide. Then, HMBPP, which was synthesized in our laboratory, was added to the chamber to obtain a final concentration of 10 μM. The plate was incubated for 2 days at 37 ℃ and 5% CO2, and collagen type I levels in DHLF cells were examined via immunocytochemistry. For Western blot analysis, DHLF cells (1.5 × 105) were placed in a 6-well plate and incubated overnight at 37 ℃ and 5% CO2. After the supernatant was removed, γδ T cell suspensions (0, 0.15 × 105, 0.75 × 105, or 1.5 × 105 cells) and HMBPP (final concentration of 10 μM) were added, and the plate was incubated at 37 ℃ and 5% CO2 for 2 days. DHLF cells were then washed twice with 2 mL of PBS (-), dissolved in RIPA buffer (as described earlier), and collagen type I and αSMA protein expression levels were examined using Western blot analysis.
To examine the effect of HMBPP on the γδ T cell-mediated modulation of collagen levels in fibroblasts, DHLF cells (1.5 × 105) treated with TGF-β (5 ng/mL) or RPMI 1640 (negative control) were incubated overnight in a 6-well plate at 37 °C and 5% CO2. The cells were washed with PBS (-), and then 1.5 mL of complete RPMI 1640 was added and culture membrane inserts were placed in the wells. γδ T cells (5.0 × 106) or 1 mL of complete RPMI 1640 medium was added to the culture membrane inserts, and HMBPP (final concentrations of 0, 0.1, 1, and 10 μM) was added. After incubation for 2 days at 37 °C and 5% CO2, the culture inserts were removed and DHLF cells were dissolved in lysis buffer, as described earlier. Then, the collagen levels were determined via Western blot analysis.
2.7. Effect of γδ T Cell Supernatants on Lung Fibroblasts
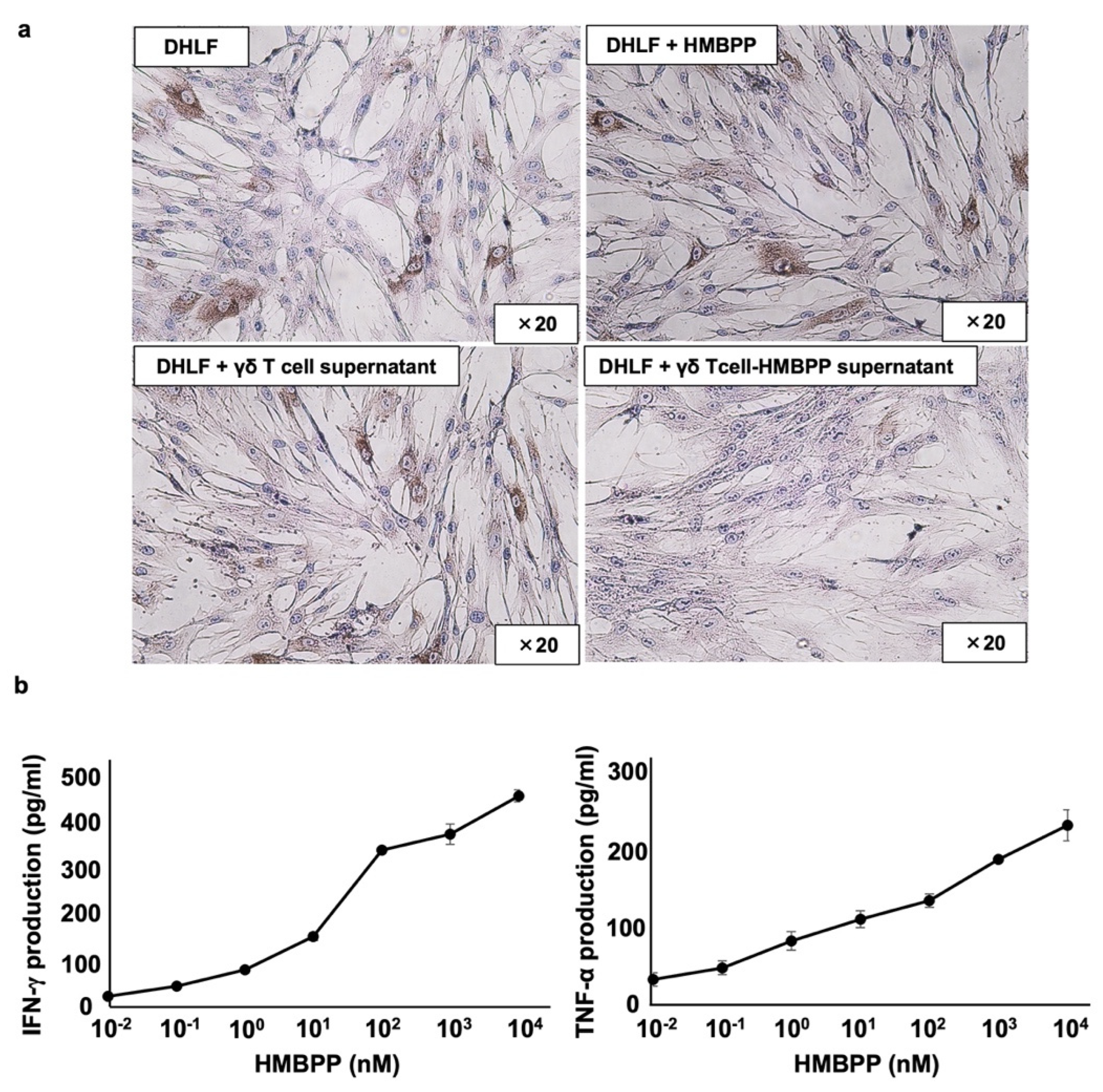
γδ T cells (5 × 106) in 1.5 mL of complete RPMI 1640 medium were cultured overnight at 37 °C and 5% CO2 in a 24-well plate (Corning Inc., Corning, NY, USA) in the presence or absence of HMBPP (final concentration of 10 μM). The supernatant was then transferred into conical tubes and centrifuged at 600× g and 4 °C for 5 min; subsequently, the supernatant was transferred into new conical tubes and stored at −80 °C until use. DHLF cells (2.5 × 104) were cultured overnight in a Lab-Tek chamber slide, and the supernatant was then aspirated. The γδ T cell culture supernatant (500 μL) was thawed and combined with 500 μL of complete RPMI 1640 medium on the chamber slide. Complete RPMI 1640 medium (1 mL) was added to the chamber slide as a control. The slide was incubated for 2 days at 37 °C and 5% CO2, and collagen type I expression was then determined via immunocytochemistry.
To examine the effect of cytokines in γδ T cell culture supernatants on the modulation of collagen levels, DHLF cells (1.5 × 105) were cultured overnight in a 6-well plate at 37 °C and 5% CO2. γδ T cell culture supernatants were thawed and transferred into conical tubes. Anti-IFN-γ antibodies (Biolegend) or anti-tumor necrosis factor-α (TNF-α) antibodies (Biolegend) were added to obtain concentrations of 0, 0.1, 1, or 10 μg/mL. The tubes were placed at room temperature for 15 min. DHLF culture supernatants were aspirated, and 1 mL of complete RPMI 1640 medium was added to the chamber slide. A mixture of γδ T cell culture supernatant and serially diluted antibody was then added to the slide. The complete RPMI 1640 medium (2 mL) was used as a control, and HMBPP (final concentration of 10 μM) was added. The plate was incubated for 2 days at 37 °C and 5% CO2, and the DHLF cells were dissolved in lysis buffer as described earlier and analyzed via Western blotting.
To examine the effect of IFN-γ on the modulation of collagen levels, γδ T cells (5 × 106) were cultured overnight in the presence or absence of HMBPP (final concentration of 0.1 nM) and interleukin-18 (IL-18; final concentration of 100 ng/mL) in a 24-well plate at 37 °C and 5% CO2. The cells were centrifuged at 600× g and 4 °C for 5 min, and the supernatants were transferred into conical tubes and stored at −80 ℃ until use. The culture supernatants were removed after incubating DHLF cells (1.5 × 105) overnight in a 6-well plate at 37 °C and 5% CO2. One milliliter of thawed γδ T cell culture supernatant and 1 mL of complete RPMI 1640 medium were added to the wells. The complete RPMI 1640 medium (2 mL) in the presence or absence of HMBPP (final concentration of 0.1 nM) and IL-18 (final concentration of 100 ng/mL) was added to the wells as a control. The plate was then incubated for 2 days at 37 ℃ and 5% CO2, after which the DHLF cells were dissolved in lysis buffer, and collagen expression was determined via Western blot assay.
To further examine the effect of IFN-γ on the modulation of collagen levels, thawed γδ T cell culture supernatants were treated with anti-IFN-γ antibodies (0, 1, or 10 μg/mL) at room temperature for 15 min. DHLF cells were then cultured in a 6-well plate, as described earlier, and the supernatants were aspirated. Then, 1 mL of γδ T cell culture supernatant in the presence or absence of anti-IFN-γ antibody and 1 mL of complete RPMI 1640 were added to the wells. Complete RPMI 1640 (2 mL) in the presence or absence of HMBPP (0.1 nM) and IL-18 (100 ng/mL) was added to the wells as a control. The plate was incubated for 2 days, and DHLF cells were analyzed for collagen expression levels via Western blot analysis.
2.8. IFN-γ and TNF-α Enzyme-Linked Immunosorbent Assay
To determine IFN-γ and TNF-α secretion from γδ T cells in response to HMBPP, γδ T cells (4 × 105 cells in 198 μL of complete RPMI 1640 medium) were added to a 96-well flat-bottom plate, and 2 μL of HMBPP was added to obtain final concentrations of 0.01 nM, 0.1 nM, 1 nM, 10 nM, 100 nM, 1 μM, or 10 μM. The plate was incubated for 24 h at 37 °C and 5% CO2. The cell suspension was mixed well, and the plate was centrifuged at 600× g and 4 °C for 2 min. The supernatant was then transferred into a 96-well round-bottom plate, and the plate was stored at −80 °C for 24 h. The samples were then thawed, and IFN-γ and TNF-α levels were analyzed via enzyme-linked immunosorbent assay (ELISA, Peprotech, Rocky Hill, NJ), according to the manufacturer’s instructions. To measure IFN-γ secretion from γδ T cells in response to HMBPP and IL-18, γδ T cells (4 × 105) were resuspended in 196 μL of complete RPMI 1640 medium in a 96-well flat-bottom plate, and 2 μL of HMBPP at final concentrations of 1, 10, 100, 1, or 10 nM and 2 μL of IL-18 (final concentration of 100 ng/mL) or complete RPMI 1640 medium were added. After incubation for 24 h at 37 °C and 5% CO2, the supernatants were transferred into a 96-well round-bottom plate and stored at −80 °C.
2.9. Effect of Human γδ T Cells on Mouse Lung Fibroblasts
Mouse lung fibroblasts (Mlg) (1.5 × 105 cells) were cultured overnight in a 6-well plate at 37 °C and 5% CO2. Culture supernatants were removed and combined with human γδ T cells (5 × 106 cells) directly or through a culture membrane insert. The effect of HMBPP (10 μM) was examined by adding HMBPP through the culture membrane insert. The plate was incubated for 2 days, and collagen levels in Mlg were measured via Western blot analysis.
2.10. Immunocytochemistry
To stain cells, culture supernatants were removed from chamber slides, and the cells were washed three times with 0.5 mL of PBS (-). Phosphate buffer containing 4% paraformaldehyde (0.5 mL, Fujifilm Wako Pure Chemical Corp. Chuo-ku, Osaka, Japan) was added to the chamber slide, and the slide was placed at room temperature for 10 min. The solution was then aspirated, and the chamber slide was washed three times with 0.5 mL of PBS (-). Hydrogen peroxide solution (0.5 mL, 0.3% in methanol, Fujifilm Wako Pure Chemical Corp., Osaka, Japan) was added to the chamber slide, and the slide was incubated at room temperature for 20 min. The solution was then aspirated, and the cells were washed three times with 0.5 mL PBS (-). Anti-collagen type I polyclonal antibody (Thermo Fisher Scientific, Waltham, MA, USA) was diluted at a ratio of 1:500 with 0.1% bovine serum albumin solution in PBS (-), and 200 μL of this diluted solution was added to the chamber slide, which was incubated overnight at 4 °C. Normal rabbit IgG (200 μL, Santa Cruz Biotechnology Inc., Santa Cruz, CA) was used as a negative control. The antibody solution was aspirated, and 0.5 mL of PBS (-) was added to the chamber slide. The chamber slide was then incubated for 3 min, and the obtained solution was discarded. This step was repeated two more times, following which two drops of Histofine Simple Stain MAX-PO(R) (Nichirei Co., Tokyo, Japan) were added to the chamber slide, and the slide was incubated for 30 min at room temperature. The solution was aspirated, and the chamber slide was washed four times with 0.5 mL of PBS (-) every 3 min. The coloring reagent (200 μL) containing 5 mg of 3,3′-diaminobenzidine tetrahydrochloride (Fujifilm Wako Pure Chemical Corp., Osaka, Japan), 10 mL of 50 mM Tris buffer (Sigma-Aldrich, St Louis, MO, USA), and 10 μL of 30% hydrogen peroxide solution (Fujifilm Wako Pure Chemical Corp., Osaka, Japan) was added to the slide, which was then incubated at room temperature for 10 min. The solution was subsequently aspirated, and the chamber slide was immersed in a tray of distilled water for 2 min and then in another tray of distilled water for 2 s. The water was aspirated, and 300 μL of Mayer’s hematoxylin solution (Fujifilm Wako Pure Chemical Corp., Osaka, Japan) was added to the chamber slide. After incubation for 10 min, the solution was removed, and the slide was immersed in a tray of distilled water for 2 min and then in another tray of distilled water for 2 s. Finally, the chamber slide was air-dried and observed under a microscope (ECLIPSE Ci, Nikon Instruments Inc., Tokyo, Japan).
2.11. Western Blot Assay
Protein concentrations in cell lysates were determined using a BCA protein assay kit (Thermo Fisher Scientific, Waltham, MA, USA), according to the manufacturer’s instructions, and optical density was measured at 540 nm using a multiplate reader (Thermo Fisher Scientific, Waltham, MA, USA). Cell lysates (30 μg in 30 μL of RIPA buffer) were diluted with equal volumes of Laemmli sample buffer (Bio-Rad, Hercules, CA, USA) containing 5% 2-mercaptoethanol (Fujifilm Wako Pure Chemical Corp., Osaka, Japan) to obtain a final protein concentration of 5 μg/10 μL. The samples were then vortexed thoroughly, placed on a heating block (AGC Techno Glass Co., Ltd., Haibara, Shizuoka, Japan) set at 95 °C for 5 min, vortexed once more, and placed on ice for 10 min. Samples (10 μL) were loaded into lanes on a 4–15% Mini-PROTEAN® TGX™ Gel (Bio-Rad, Hercules, CA, USA) and run at 200 V and 40 mA for 30 min. The protein bands were transferred onto a membrane for 7 min using a Trans-Blot® TurboTM Transfer System (MW-1.3A, Bio-Rad, Hercules, CA, USA). The membrane was then immersed in a plastic container with 20 mL blocking buffer containing 5% BSA in Tris-buffered saline with Tween 20 (TBST, 1 tablet in 500 mL of distilled water, Takara Bio Inc., Kusatsu, Shiga, Japan) and incubated for 1 h. After the blocking buffer was removed, the membrane was washed with TBST, cut into pieces, and incubated overnight at 4 °C in a Hybri-Bag (Cosmo Bio Co., Ltd., Koto-ku, Tokyo, Japan) containing rabbit anti-collagen I polyclonal antibody (1/5000 dilution, Thermo Fisher Scientific, Waltham, MA, USA) or mouse anti-GAPDH polyclonal antibody (1/1000 dilution, Thermo Fisher Scientific, Waltham, MA, USA). The membrane was subsequently washed three times with 20 mL of TBST for 10 min and then treated with horseradish peroxidase (HRP)-labeled secondary antibody, anti-rabbit IgG antibody (1/5000 dilution, Abcam, Cambridge, UK), or anti-mouse IgG antibody (1/1000 dilution, Biotechne, Minneapolis, MN, USA) at room temperature for 1 h. The membranes (for collagen type I) were washed three times with 20 mL of TBST for 10 min and treated with ClarityTM Western ECL Substrate (Bio-Rad, Hercules, CA, USA). Membrane luminescence was monitored using a ChemiDoc™ MP Imaging System (Bio-Rad, Hercules, CA, USA). The membranes (for GAPDH) were washed with TBST for 15 min and treated with RestoreTM PLUS Western blot stripping buffer (Thermo Fisher Scientific, Waltham, MA, USA) for 15 min and then with TBST for 15 min before the treatment with blocking buffer for 1 h. The membranes were then placed in a Hybri-Bag containing anti-αSMA antibody (1/1000 dilution, Abcam) and incubated overnight at 4 °C. Subsequently, the membranes were washed three times with TBST for 10 min and incubated at room temperature for 1 h with the secondary antibody (anti-rabbit IgG antibody). The membranes were subsequently washed three times with TBST for 10 min, and luminescence was measured as described earlier. Relative collagen and αSMA expression intensities were calculated by normalization to GAPDH levels using Image Lab™ software ver. 5.2.1 (Bio-Rad, Hercules, CA, USA).
2.12. Real-Time RT-PCR
Cells were dissolved in lysis buffer and transferred to a microcentrifuge tube. The tube was vortexed three times for 10 s and spun down. Total RNA was extracted from the supernatant using a QIAcube kit (QIAGEN, Hilden, Dusseldorf, Germany), according to the manufacturer’s instructions, and quantified using a Nano Drop spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). Purified RNA was diluted with diethylpyrocarbonate-treated water (part of the SuperScript®Ⅲ First-Strand Synthesis System (SS III kit), Thermo Fisher Scientific, Waltham, MA, USA) to obtain a concentration of 2 μg/8 μL, and transferred into PCR tubes (NIPPON Genetics Co., Ltd., Tokyo, Japan) on ice. Two microliters of oligo(dT)20 (50 μM, Thermo Fisher Scientific, Waltham, MA, USA) and deoxyribonucleotides (10 mM, part of the SS III kit) were added to the diluted RNA (8 μL), and this mixture was placed on ice. The samples were then centrifuged at 14,000 rpm and 4 °C for 7 s and incubated at 65 ℃ for 5 min on a T100TM thermal cycler (Bio-Rad, Hercules, CA, USA). The samples were incubated on ice for 1 min, followed by the addition of 10 μL of a reagent consisting of 10× reverse transcription buffer (2.25 μL), 25 mM MgCl (4.5 μL), 0.1 M dithiothreitol (2.25 μL), RNaseOUTTM (40 U/μL, 1.125 μL), and SuperScript® III RT (200 U/μL, 1.125 μL; part of the SS III kit). The samples were incubated at 50 °C for 50 min and then at 85 °C for 5 min on a thermal cycler (Bio-Rad, Hercules, CA, USA). Subsequently, the samples were centrifuged at 14,000 rpm for 7 s at 4 °C. RNase H (1 μL, part of the SS III kit) was added, and the tubes were incubated at 37 ℃ for 20 min on the thermal cycler. The synthesized cDNA was diluted 2.5-fold with nuclease-free water. Then, the diluted cDNA (2.2 μL) was transferred into a 96-well plate (Thermo Fisher Scientific, Waltham, MA, USA), and 20.0 μL of a reagent consisting of TaqManTM Universal PCR Master Mix, no AmpEraseTM UNG (11.25 μL, Thermo Fisher Scientific, Waltham, MA, USA), TaqMan® Gene Expression Assay (1.125 μL, COL1A1, COL1A2, or GAPDH mRNA, Thermo Fisher Scientific, Waltham, MA, USA), and nuclease-free water (7.65 μL) was added. The plate was sealed with MicroAmp® Optical Adhesive Film (Thermo Fisher Scientific, Waltham, MA, USA) and centrifuged at 170 g and 4 °C for 15 s, and the samples were analyzed using a QuantStudioTM 12K Flex real-time PCR system (Thermo Fisher Scientific, Waltham, MA, USA). Data were processed using QuantStudioTM 12K Flex software V1.x (Thermo Fisher Scientific, Waltham, MA, USA). Relative COL1A1 and COL1A2 mRNA expression levels were calculated after normalization to GAPDH levels.
2.13. Statistical Analyses
Statistical significance was determined using Dunnett’s test, implemented in JMP Pro software ver. 14.0. p < 0.05 was considered statistically significant.
4. Discussion
Several studies have reported the effect of human γδ T cells on the regulation of collagen type I in fibroblasts, although the results are contradictory [
17,
19,
20,
22], partially because the purity, quality, and number of γδ T cells were insufficient to appropriately evaluate these effects. In this study, we prepared a large number of purified human γδ T cells using PTA, a nitrogen-containing bisphosphonate prodrug that efficiently stimulates γδ T cells [
24,
25]. The purity of the PTA/IL-2-expanded γδ T cells used in this study was at least 98%, and the cells expanded for 11 days before γδ T cell effector functions deteriorated.
PTA is a highly hydrophobic compound that readily penetrates cell membranes. Once the compound is internalized by cells, it is hydrolyzed by intracellular esterases to give TA, a biologically active bisphosphonate that inhibits FDPS [
23]. When FDPS is inhibited, the immediate upstream metabolites, IPP and DMAPP, accumulate in the cytoplasm and bind to the B30.2 intracellular domain of butyrophilin 3A1 (BTN3A1) [
25,
26]. Although the precise mechanism has not been elucidated, the B30.2/IPP/DMAPP complex activates γδ T cells in a BTN3A1/BTN2A1-dependent manner [
27]. This protocol for expanding γδ T cells facilitated the examination of the role of γδ T cells in the suppression of collagen type I in lung fibroblasts.
PTA/IL-2-induced γδ T cells effectively reduced the quantities of collagen type I in lung fibroblasts, in a cell–cell contact-dependent manner, based on the results from a trans-well plate assay system. Therefore, we hypothesized that certain adhesion molecules may be involved in the γδ T cell-mediated suppression of collagen type I [
28,
29]. Several adhesion molecules that play a pivotal role in the adhesion of γδ T cells to fibroblasts are expressed on γδ T cell surfaces, including LFA-1, very late antigen-4 (VLA-4), and VLA-5 [
30,
31,
32]. LFA-1 is expressed on IPP-stimulated γδ T cells and is activated following its ligation to its ligands, including ICAM-1 [
31,
32].
In this study, we demonstrated that PTA/IL-2-expanded γδ T cells expressed high LFA-1 levels. Treating γδ T cells with anti-LFA-1 neutralizing mAb, however, did not reverse the suppression of collagen type I expression, suggesting that the modulation of collagen type I in lung fibroblasts by γδ T cells is not mediated by LFA-1.
Since human γδ T cells exhibited TCR-independent cytotoxicity against human lung cancer cells [
25], it is possible that the suppression of collagen type I expression by γδ T cells is caused by γδ T cell-mediated cytotoxic effects against lung fibroblasts. Based on the cytotoxicity assay results, however, γδ T cells showed no cytotoxicity against human lung fibroblasts under the conditions in which this study was conducted. Although γδ T cells did not alter the viability of lung fibroblasts, they significantly suppressed the expression of COL1A1 and COL1A2 mRNAs in lung fibroblasts, suggesting that these cells transduce signals in fibroblasts, inhibiting the transcription and translation of collagen type I genes, wherein the interaction between γδ T cells and fibroblasts plays an essential role in signal transduction. Cell–cell contact-dependent suppression of collagen type I was shown to be species-specific, suggesting the involvement of human-specific ligand-receptor combination in signal transduction.
The effect of activated γδ T cells on the regulation of collagen type I in lung fibroblasts was examined because activated γδ T cells express a variety of effector molecules that may mediate the suppression of collagen type I expression. We used HMBPP, a phosphoantigen, to activate the γδ T cells. This phosphoantigen is biosynthesized through the 2-C-methyl-D-erythritol 4-phosphate (MEP) pathway and is converted to IPP in bacteria, protozoa, and plants [
33]. HMBPP binds B30.2 of BTN3A1-like IPP and induces effector functions in γδ T cells, including cytotoxicity against tumor cells and the secretion of proinflammatory cytokines and growth factors [
33,
34].
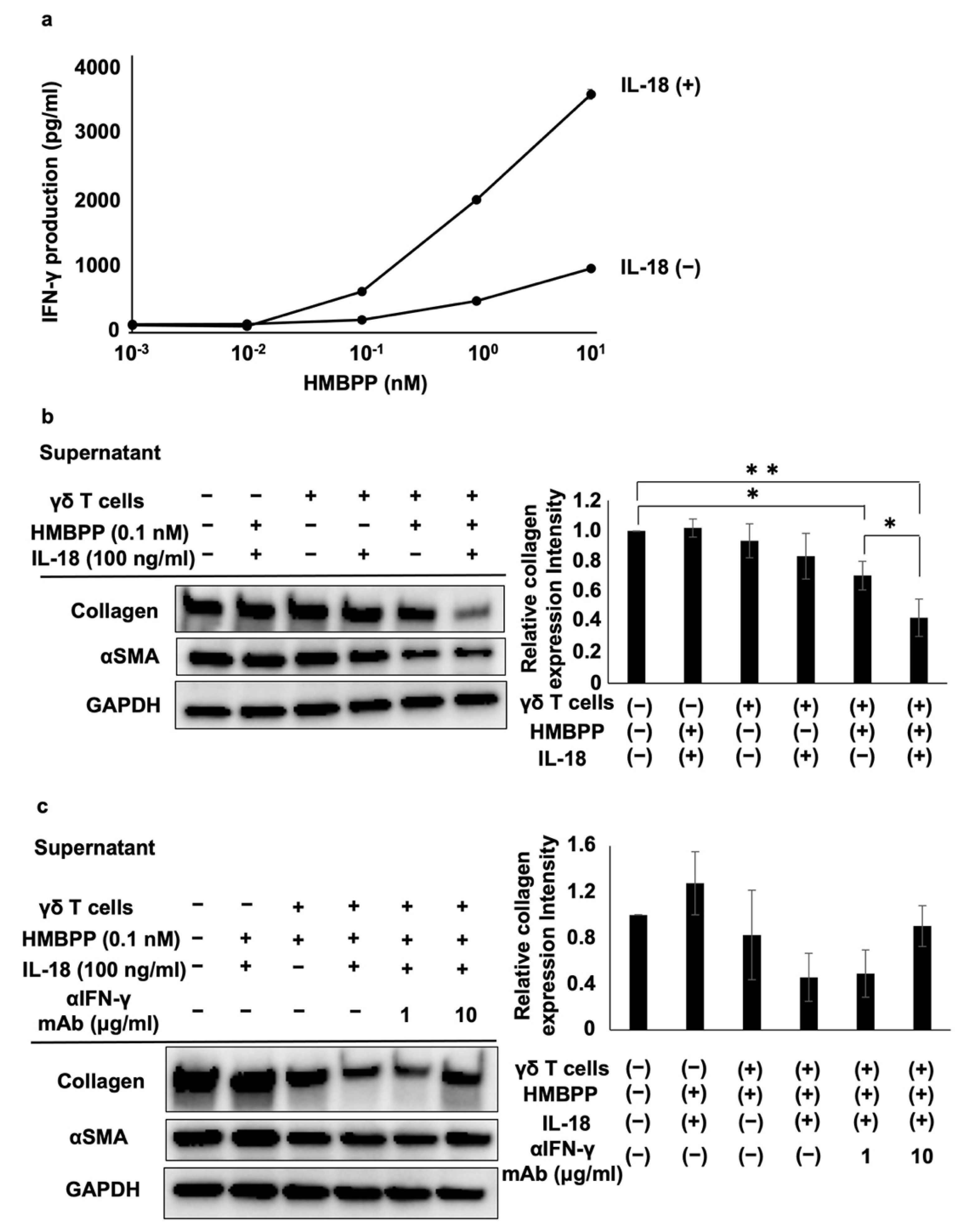
HMBPP-activated γδ T cells suppressed collagen type I levels in human lung fibroblasts more efficiently than unstimulated γδ T cells. Thus, we examined whether HMBPP-activated γδ T cells secrete soluble factors that might be involved in the suppression of collagen type I. Supernatants of HMBPP-activated γδ T cells suppressed collagen type I in lung fibroblasts, unlike unstimulated γδ T cells, suggesting that certain humoral factors secreted by HMBPP-activated γδ T cells are involved in collagen suppression. Since human γδ T cells can produce antifibrotic cytokines such as IFN-γ and TNF-α [
35,
36,
37], we examined the effect of these cytokines secreted from HMBPP-activated γδ T cells. ELISA-based analysis using neutralizing mAbs showed that IFN-γ, but not TNF-α, was involved in the suppression of collagen type I in lung fibroblasts, although both cytokines previously demonstrated the ability to inhibit collagen synthesis [
38,
39,
40], suggesting that IFN-γ plays a major role in the γδ T cell-mediated suppression of collagen type I in lung fibroblasts. Some in vitro studies have shown that IFN-γ inhibits collagen synthesis and type I procollagen mRNA levels in fibroblasts and suppresses fibroblast proliferation [
40,
41]. Furthermore, an in vivo study indicated that IFN-γ administration decreases fibroblast numbers and hydroxyproline content in a bleomycin-induced pulmonary fibrosis mouse model [
42]. Therefore, IFN-γ is closely associated with lung fibroblasts, and our findings are consistent with the findings of these previous reports.
Since IL-18 is a member of the IL-1 cytokine superfamily, enhances IFN-γ production, and stimulates effector T cells together with IL-12, IL-2, or antigens, we examined the effect of IL-18 on the suppression of collagen type I in lung fibroblasts [
43,
44]. As expected, collagen type I expression was significantly suppressed by supernatants of HMBPP/IL-18-stimulated γδ T cells, even at low HMBPP concentrations. Adding anti-IFN-γ neutralizing mAb reversed the suppressive effect of the supernatants, suggesting that IFN-γ is a pivotal factor in the supernatants of HMBPP/IL-18-stimulated γδ T cells that suppresses collagen type I in lung fibroblasts.
Collagen type I levels in mouse fibroblasts were not altered by treatment with the supernatants of HMBPP/IL-18-stimulated human γδ T cells because human IFN-γ is not recognized by murine IFN-γ receptors. Although murine γδ T cells have antifibrotic properties [
11,
12,
13,
14,
15,
16], it is difficult to establish murine models for studying the suppression of collagen type I by HMBPP/IL-18-stimulated γδ T cells because mice lack an HMBPP-reactive γδ T cell equivalent [
45]. Currently, efforts are being devoted to developing protocols for the adoptive transfer of human γδ T cells in the field of cancer immunotherapy, which is well tolerated [
46].
Since antifibrotic therapies based on small-molecule drugs do not currently have satisfactory outcomes, it is imperative to develop alternative therapeutic modalities such as γδ T cell-based immunotherapies for lung fibrosis. Since our γδ T cell culture system based on PTA allows us to prepare a large number of highly purified human γδ T cells with no restriction to major histocompatibility complex class I or II, it may be possible to develop γδ T cell-based off-the-shelf drugs for lung fibrotic diseases.
There are some limitations to this study. First, the detailed mechanism underlying the direct inhibition of collagen in lung fibroblasts by γδ T cells is unknown. Although cell–cell contact is important for collagen suppression, additional studies are needed to examine cell-surface markers and pathways involved in collagen inhibition by γδ T cells. Second, there might be other humoral factors in the supernatant of γδ T cells that are associated with collagen inhibition. Although IFN-γ plays an important role in collagen inhibition by the supernatant of γδ T cells, the collagen-suppressive effect of the supernatant was not completely reversed by anti-IFN-γ mAb alone. This suggests that other humoral factors secreted by γδ T cells might also be related to collagen inhibition. Despite these limitations, this study provides considerable insight into the development of γδ T cell-based immunotherapies for lung fibrosis.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





