
Age and Sex-Related Changes in Retinal Function in the Vervet Monkey

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
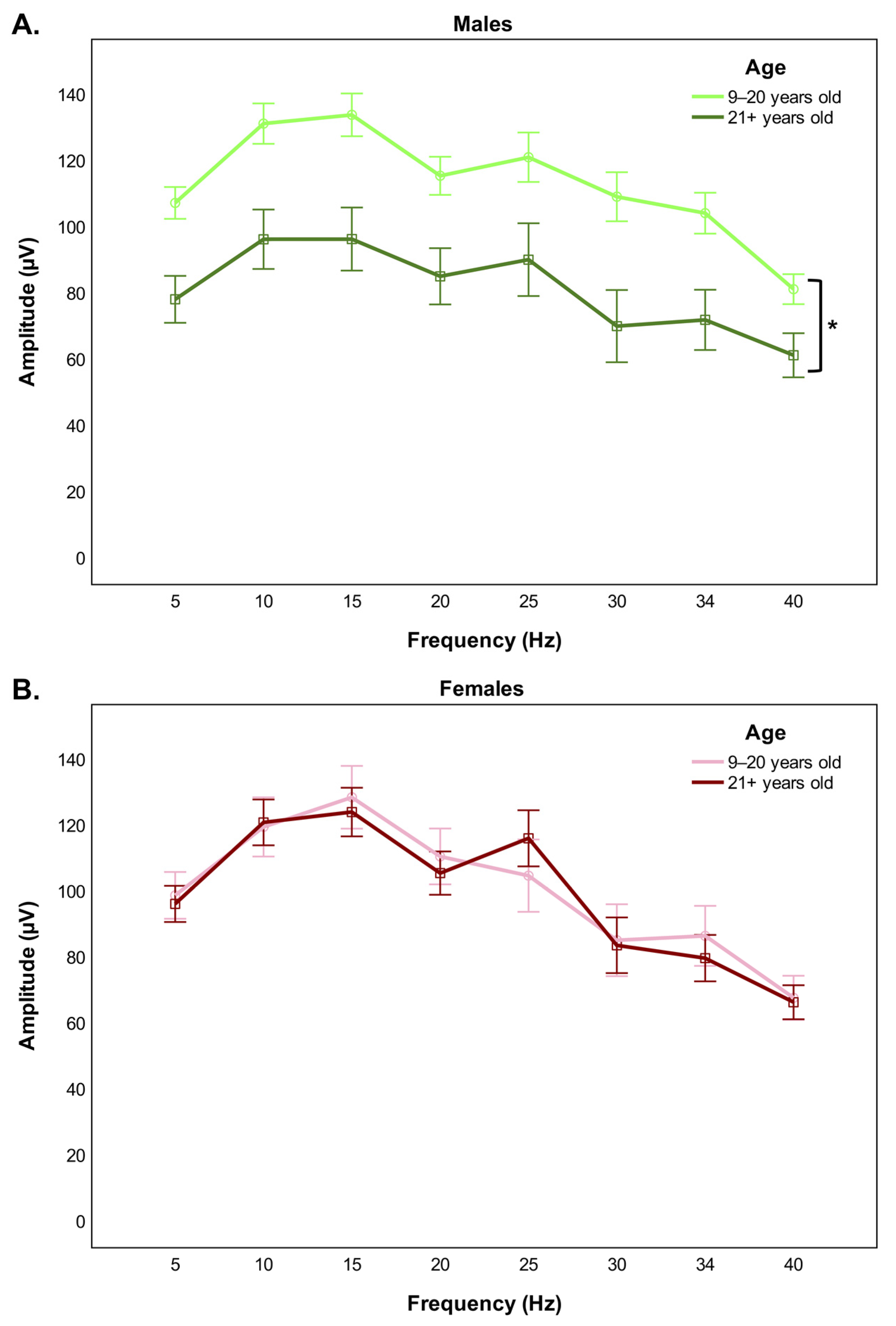
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Animal Preparation
2.3. Intraocular Pressure (IOP) Measurement
2.4. Photopic Flicker Electroretinogram (ERG)
2.5. Statistical Analysis
3. Results
3.1. Intraocular Pressure
3.2. Photopic Flicker ERG
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Panda-Jonas, S.; Jonas, J.B.; Jakobczyk-Zmija, M. Retinal photoreceptor density decreases with age. Ophthalmology 1995, 102, 1853–1859. [Google Scholar] [CrossRef]
- Cavallotti, C.; Artico, M.; Pescosolido, N.; Feher, J. Age-related changes in rat retina. Jpn. J. Ophthalmol. 2001, 45, 68–75. [Google Scholar] [CrossRef]
- Cavallotti, C.; Artico, M.; Pescosolido, N.; Leali, F.M.T.; Feher, J. Age-related changes in the human retina. Can. J. Ophthalmol. 2004, 39, 61–68. [Google Scholar] [CrossRef]
- Karpe, G.; Rickenbach, K.; Thomasson, S. The clinical electroretinogram: I. The Normal electroretinogram above fifty years of age. Acta Ophthalmol. 1950, 28, 301–305. [Google Scholar] [CrossRef]
- Zeidler, I. The clinical electroretinogram: IX. The Normal Electroretinogram.: Value of the b-Potential in Different Age Groups and Its Differences in Men and Women. Acta Ophthalmol. 1959, 37, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Weleber, R.G. The effect of age on human cone and rod ganzfeld electroretinograms. Investig. Ophthalmol. Vis. Sci. 1981, 20, 392–399. [Google Scholar]
- Wright, C.; Williams, D.; Drasdo, N.; Harding, G. The influence of age on the electroretinogram and visual evoked potential. Doc. Ophthalmol. 1985, 59, 365–384. [Google Scholar] [CrossRef]
- Birch, D.G.; Anderson, J.L. Standardized full-field electroretinography: Normal values and their variation with age. Arch. Ophthalmol. 1992, 110, 1571–1576. [Google Scholar] [CrossRef]
- Kergoat, H.; Kergoat, M.J.; Justino, L. Age-Related Changes in the Flash Electroretinogram and Oscillatory Potentials in Individuals Age 75 and Older. J. Am. Geriatr. Soc. 2001, 49, 1212–1217. [Google Scholar] [CrossRef]
- Neveu, M.M.; Dangour, A.; Allen, E.; Robson, A.G.; Bird, A.C.; Uauy, R.; Holder, G.E. Electroretinogram measures in a septuagenarian population. Doc. Ophthalmol. 2011, 123, 75–81. [Google Scholar] [CrossRef]
- Trick, G.L.; Nesher, R.; Cooper, D.G.; Shields, S.M. The human pattern ERG: Alteration of response properties with aging. Optom. Vis. Sci. 1992, 69, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Brûlé, J.; Lavoie, M.-P.; Casanova, C.; Lachapelle, P.; Hébert, M. Evidence of a possible impact of the menstrual cycle on the reproducibility of scotopic ERGs in women. Doc. Ophthalmol. 2007, 114, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Nadal-Nicolás, F.M.; Vidal-Sanz, M.; Agudo-Barriuso, M. The aging rat retina: From function to anatomy. Neurobiol. Aging 2018, 61, 146–168. [Google Scholar] [CrossRef] [PubMed]
- Chaychi, S.; Polosa, A.; Lachapelle, P. Differences in retinal structure and function between aging male and female Sprague-Dawley rats are strongly influenced by the estrus cycle. PLoS ONE 2015, 10, e0136056. [Google Scholar] [CrossRef]
- Rodrigo, M.J.; Martinez-Rincon, T.; Subias, M.; Mendez-Martinez, S.; Luna, C.; Pablo, L.E.; Polo, V.; Garcia-Martin, E. Effect of age and sex on neurodevelopment and neurodegeneration in the healthy eye: Longitudinal functional and structural study in the Long–Evans rat. Exp. Eye Res. 2020, 200, 108208. [Google Scholar] [CrossRef]
- Bouskila, J.; Javadi, P.; Palmour, R.M.; Bouchard, J.-F.; Ptito, M. Standardized full-field electroretinography in the Green Monkey (Chlorocebus sabaeus). PLoS ONE 2014, 9, e111569. [Google Scholar] [CrossRef]
- Robson, A.G.; Frishman, L.J.; Grigg, J.; Hamilton, R.; Jeffrey, B.G.; Kondo, M.; Li, S.; McCulloch, D.L. ISCEV Standard for full-field clinical electroretinography (2022 update). Doc. Ophthalmol. 2022, 144, 165–177. [Google Scholar] [CrossRef]
- Bouskila, J.; Palmour, R.M.; Bouchard, J.-F.; Ptito, M. Retinal structure and function in monkeys with fetal alcohol exposure. Exp. Eye Res. 2018, 177, 55–64. [Google Scholar] [CrossRef]
- Han, Y.; Bearse, M.A.; Schneck, M.E.; Barez, S.; Jacobsen, C.H.; Adams, A.J. Multifocal electroretinogram delays predict sites of subsequent diabetic retinopathy. Investig. Ophthalmol. Vis. Sci. 2004, 45, 948–954. [Google Scholar] [CrossRef]
- Bearse Jr, M.A.; Adams, A.J.; Han, Y.; Schneck, M.E.; Ng, J.; Bronson-Castain, K.; Barez, S. A multifocal electroretinogram model predicting the development of diabetic retinopathy. Prog. Retin. Eye Res. 2006, 25, 425–448. [Google Scholar] [CrossRef]
- Wenner, M.M.; Stachenfeld, N.S. Blood pressure and water regulation: Understanding sex hormone effects within and between men and women. J. Physiol. 2012, 590, 5949–5961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cascio, C.; Deidda, I.; Russo, D.; Guarneri, P. The estrogenic retina: The potential contribution to healthy aging and age-related neurodegenerative diseases of the retina. Steroids 2015, 103, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Cascio, C.; Russo, D.; Drago, G.; Galizzi, G.; Passantino, R.; Guarneri, R.; Guarneri, P. 17β-Estradiol synthesis in the adult male rat retina. Exp. Eye Res. 2007, 85, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Kobayashi, H.; Ueda, M.; Honda, Y. Estrogen receptor expression in bovine and rat retinas. Investig. Ophthalmol. Vis. Sci. 1998, 39, 2105–2110. [Google Scholar]
- Ogueta, S.B.; Schwartz, S.D.; Yamashita, C.K.; Farber, D.B. Estrogen receptor in the human eye: Influence of gender and age on gene expression. Investig. Ophthalmol. Vis. Sci. 1999, 40, 1906–1911. [Google Scholar]
- Munaut, C.; Lambert, V.; Noël, A.; Frankenne, F.; Deprez, M.; Foidart, J.-M.; Rakic, J.-M. Presence of oestrogen receptor type β in human retina. Br. J. Ophthalmol. 2001, 85, 877–882. [Google Scholar] [CrossRef]
- Kumar, D.M.; Perez, E.; Aoun, P.; Brun-Zinkernagel, A.-M.; Covey, D.F.; Simpkins, J.W.; Agarwal, N. Role of nonfeminizing estrogen analogues in neuroprotection of rat retinal ganglion cells against glutamate-induced cytotoxicity. Free Radic. Biol. Med. 2005, 38, 1152–1163. [Google Scholar] [CrossRef]
- Mo, M.-S.; Li, H.-B.; Wang, B.-Y.; Wang, S.-L.; Zhu, Z.-L.; Yu, X.-R. PI3K/Akt and NF-κB activation following intravitreal administration of 17β-estradiol: Neuroprotection of the rat retina from light-induced apoptosis. Neuroscience 2013, 228, 1–12. [Google Scholar] [CrossRef]
- Nixon, E.; Simpkins, J.W. Neuroprotective effects of nonfeminizing estrogens in retinal photoreceptor neurons. Investig. Ophthalmol. Vis. Sci. 2012, 53, 4739–4747. [Google Scholar] [CrossRef]
- Wang, S.; Wang, B.; Feng, Y.; Mo, M.; Du, F.; Li, H.; Yu, X. 17β-Estradiol ameliorates light-induced retinal damage in Sprague–Dawley rats by reducing oxidative stress. J. Mol. Neurosci. 2015, 55, 141–151. [Google Scholar] [CrossRef]
- Smith, W.; Mitchell, P.; Wang, J. Gender, oestrogen, hormone replacement and age-related macular degeneration: Results from the Blue Mountains Eye Study. Aust. New Zealand J. Ophthalmol. 1997, 25, 13–15. [Google Scholar] [CrossRef] [PubMed]
- Snow, K.K.; Cote, J.; Yang, W.; Davis, N.J.; Seddon, J.M. Association between reproductive and hormonal factors and age-related maculopathy in postmenopausal women. Am. J. Ophthalmol. 2002, 134, 842–848. [Google Scholar] [CrossRef]
- Feskanich, D.; Cho, E.; Schaumberg, D.A.; Colditz, G.A.; Hankinson, S.E. Menopausal and reproductive factors and risk of age-related macular degeneration. Arch. Ophthalmol. 2008, 126, 519–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patnaik, J.L.; Lynch, A.M.; Wagner, B.D.; Echalier, E.L.; Kohrt, W.M.; Mathias, M.T.; Siringo, F.S.; Palestine, A.G.; Mandava, N. Hormone therapy as a protective factor for age-related macular degeneration. Ophthalmic Epidemiol. 2020, 27, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Atkins, H.M.; Willson, C.J.; Silverstein, M.; Jorgensen, M.; Floyd, E.; Kaplan, J.R.; Appt, S.E. Characterization of ovarian aging and reproductive senescence in vervet monkeys (Chlorocebus aethiops sabaeus). Comp. Med. 2014, 64, 55–62. [Google Scholar] [PubMed]
- El-Mofty, A.A.; Eisner, G.; Balazs, E.A.; Denlinger, J.L.; Gouras, P. Retinal degeneration in rhesus monkeys, Macaca mulatta. Survey of three seminatural free-breeding colonies. Exp. Eye Res. 1980, 31, 147–166. [Google Scholar] [CrossRef]
- Kim, C.B.; Ver Hoeve, J.N.; Kaufman, P.L.; Nork, M.T. Interspecies and gender differences in multifocal electroretinograms of cynomolgus and rhesus macaques. Doc. Ophthalmol. 2004, 109, 73–86. [Google Scholar] [CrossRef]
- Choi, K.-E.; Anh, V.T.Q.; Yun, C.; Kim, Y.-J.; Jung, H.; Eom, H.; Shin, D.; Kim, S.-W. Normative Data of Ocular Biometry, Optical Coherence Tomography, and Electrophysiology Conducted for Cynomolgus Macaque Monkeys. Transl. Vis. Sci. Technol. 2021, 10, 14. [Google Scholar] [CrossRef]
- Sample, P.; Esterson, F.; Weinreb, R.; Boynton, R. The aging lens: In vivo assessment of light absorption in 84 human eyes. Investig. Ophthalmol. Vis. Sci. 1988, 29, 1306–1311. [Google Scholar]
- Rotenstreich, Y.; Fishman, G.; Anderson, R.; Birch, D. Interocular amplitude differences of the full field electroretinogram in normal subjects. Br. J. Ophthalmol. 2003, 87, 1268–1271. [Google Scholar] [CrossRef]
- Ordy, J.M.; Brizee, K.R.; Hansche, J. Visual acuity and foveal cone density in the retina of the aged rhesus monkey. Neurobiol. Aging 1980, 1, 133–140. [Google Scholar] [CrossRef]
- Keunen, J.; Van Norren, D.; Van Meel, G. Density of foveal cone pigments at older age. Investig. Ophthalmol. Vis. Sci. 1987, 28, 985–991. [Google Scholar]
- Gartner, S.; Henkind, P. Aging and degeneration of the human macula. 1. Outer nuclear layer and photoreceptors. Br. J. Ophthalmol. 1981, 65, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Hollyfield, J. Aging of the human retina. Differential loss of neurons and retinal pigment epithelial cells. Investig. Ophthalmol. Vis. Sci. 1992, 33, 1–17. [Google Scholar]
- Curcio, C.; Millican, C.L.; Allen, K.; Kalina, R. Aging of the human photoreceptor mosaic: Evidence for selective vulnerability of rods in central retina. Investig. Ophthalmol. Vis. Sci. 1993, 34, 3278–3296. [Google Scholar]
- CIDECIYAN, A.V.; JACOBSON, S.G. An alternative phototransduction model for human rod and cone ERG a-waves: Normal parameters and variation with age. Vis. Res. 1996, 36, 2609–2621. [Google Scholar] [CrossRef]
- Freund, P.R.; Watson, J.; Gilmour, G.S.; Gaillard, F.; Sauvé, Y. Differential changes in retina function with normal aging in humans. Doc. Ophthalmol. 2011, 122, 177–190. [Google Scholar] [CrossRef]
- Chirco, K.; Sohn, E.; Stone, E.; Tucker, B.; Mullins, R. Structural and molecular changes in the aging choroid: Implications for age-related macular degeneration. Eye 2017, 31, 10–25. [Google Scholar] [CrossRef]
- Bonilha, V.L. Age and disease-related structural changes in the retinal pigment epithelium. Clin. Ophthalmol. 2008, 2, 413. [Google Scholar] [CrossRef]
- Marin-Castano, M.E.; Elliot, S.J.; Potier, M.; Karl, M.; Striker, L.J.; Striker, G.E.; Csaky, K.G.; Cousins, S.W. Regulation of estrogen receptors and MMP-2 expression by estrogens in human retinal pigment epithelium. Investig. Ophthalmol. Vis. Sci. 2003, 44, 50–59. [Google Scholar] [CrossRef]
- Liang, F.-Q.; Godley, B.F. Oxidative stress-induced mitochondrial DNA damage in human retinal pigment epithelial cells: A possible mechanism for RPE aging and age-related macular degeneration. Exp. Eye Res. 2003, 76, 397–403. [Google Scholar] [CrossRef]
- Wang, A.L.; Lukas, T.J.; Yuan, M.; Neufeld, A.H. Increased mitochondrial DNA damage and down-regulation of DNA repair enzymes in aged rodent retinal pigment epithelium and choroid. Mol. Vis. 2008, 14, 644. [Google Scholar] [PubMed]
- Wang, A.L.; Lukas, T.J.; Yuan, M.; Neufeld, A.H. Age-related increase in mitochondrial DNA damage and loss of DNA repair capacity in the neural retina. Neurobiol. Aging 2010, 31, 2002–2010. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Chen, M.; Forrester, J.V. Para-inflammation in the aging retina. Prog. Retin. Eye Res. 2009, 28, 348–368. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Micaelo-Fernandes, C.; Bouskila, J.; Palmour, R.M.; Bouchard, J.-F.; Ptito, M. Age and Sex-Related Changes in Retinal Function in the Vervet Monkey. Cells 2022, 11, 2751. https://doi.org/10.3390/cells11172751
Micaelo-Fernandes C, Bouskila J, Palmour RM, Bouchard J-F, Ptito M. Age and Sex-Related Changes in Retinal Function in the Vervet Monkey. Cells. 2022; 11(17):2751. https://doi.org/10.3390/cells11172751
Chicago/Turabian StyleMicaelo-Fernandes, Catarina, Joseph Bouskila, Roberta M. Palmour, Jean-François Bouchard, and Maurice Ptito. 2022. "Age and Sex-Related Changes in Retinal Function in the Vervet Monkey" Cells 11, no. 17: 2751. https://doi.org/10.3390/cells11172751