
Influence of Tamoxifen on Different Biological Pathways in Tumorigenesis and Transformation in Adipose-Derived Stem Cells, Mammary Cells and Mammary Carcinoma Cell Lines—An In Vitro Study
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Cell Culture of ASC
2.2. Cell Culture of MCF-10A Mammary Cell Line
2.3. Cell Culture of MCF-7 Mammary Carcinoma Cell Line
2.4. Cell Culture of BT-474 Mammary Carcinoma Cell Line
2.5. Co-Culture of Cells
2.6. Tamoxifen Treatment of Cells
2.7. RNA Extraction and Quantitative Real-Time-PCR Array
3. Results
3.1. PCR Array of MCF-10A Showed Increased Gene Expression after Tamoxifen Treatment and Co-Cultures with ASC Showed Decreased Gene Expression
3.2. Co-Cultures of MCF-7 Mammary Carcinoma Cell Line and ASC Showed Increased Gene Expression with Increasing Tamoxifen Concentrations after 48 h
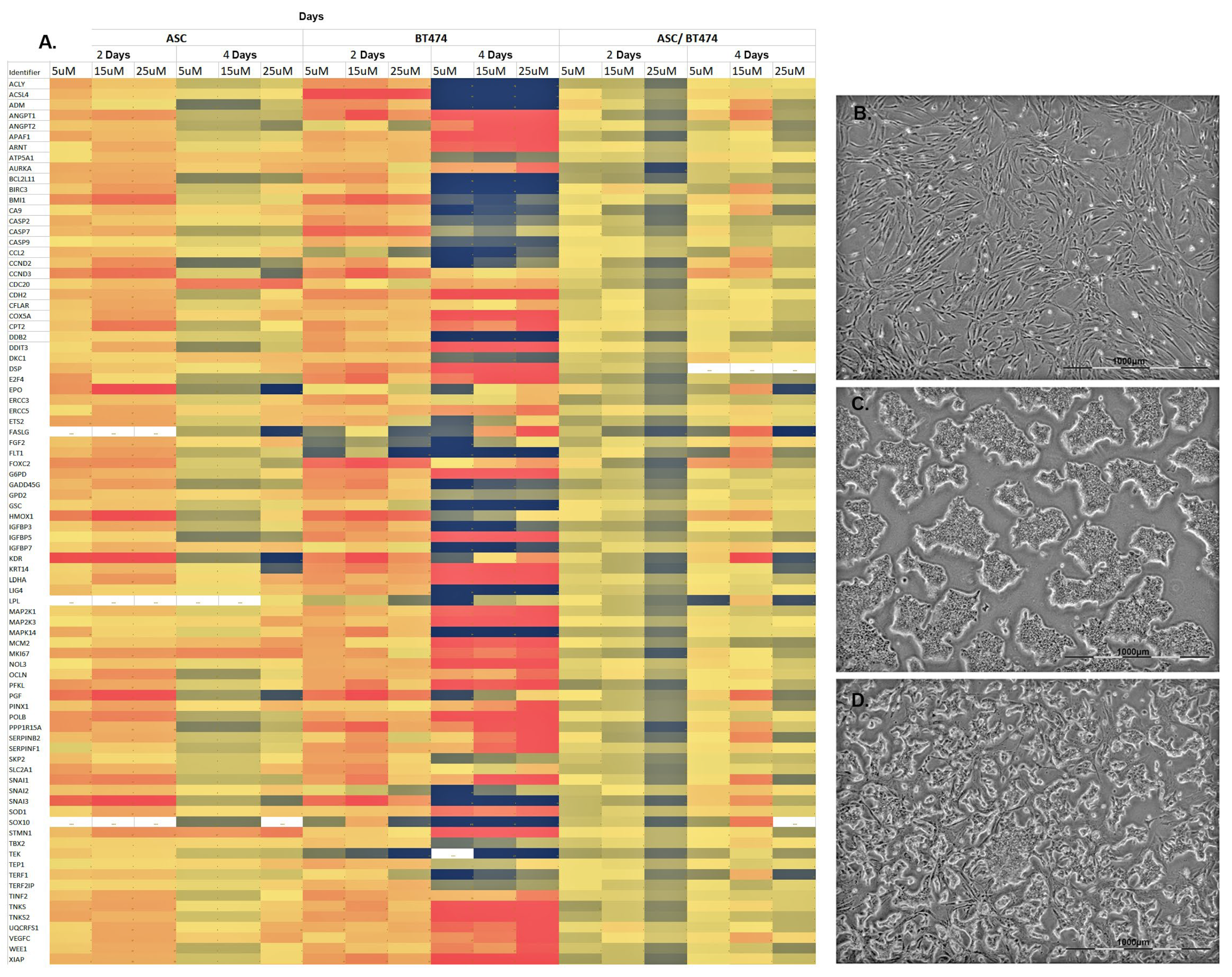
3.3. Co-Culture of BT-474 Mammary Carcinoma Cell Line and ASC Showed a Moderate Increase in Gene Expression of Pathways Involved in Tumorigenesis and Transformation
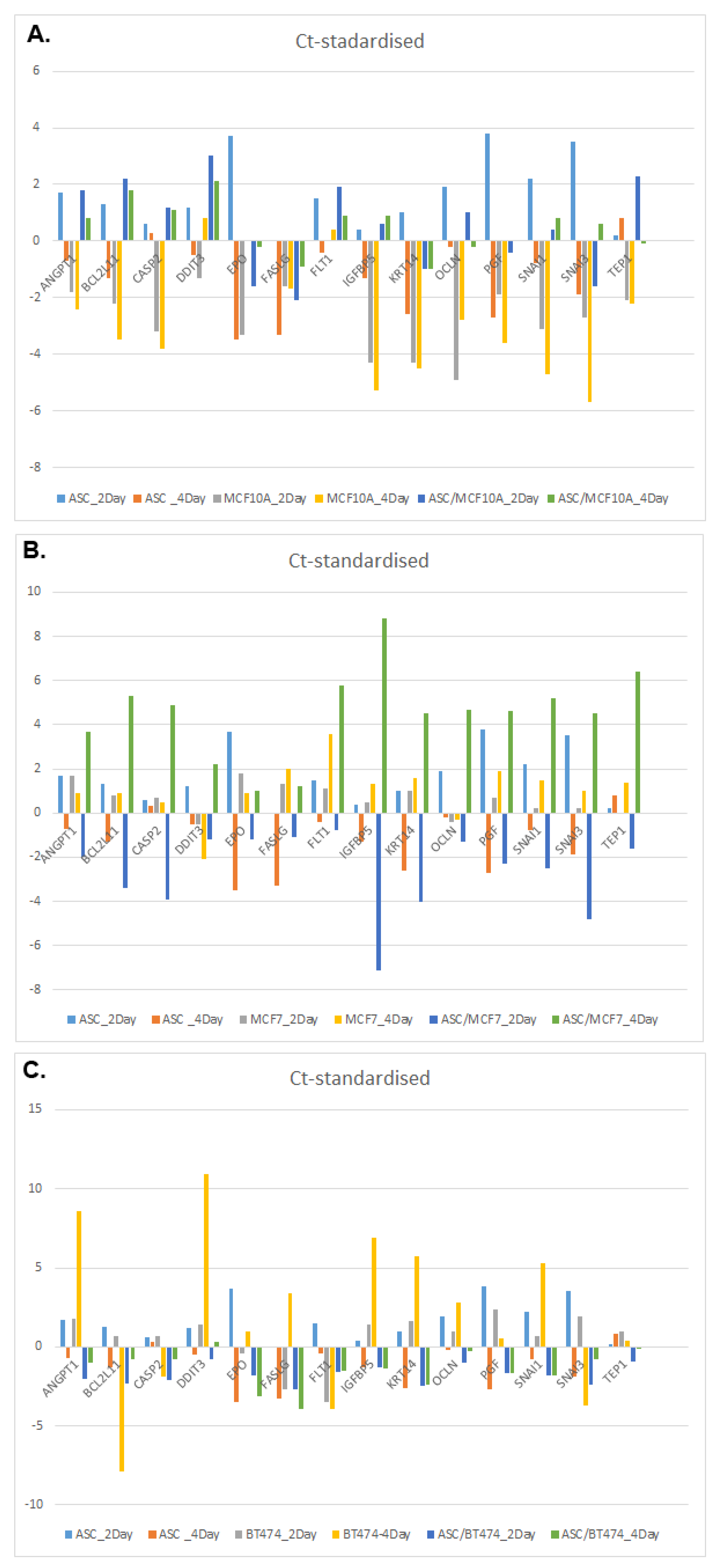
3.4. Comparison of 14 Genes with the Strongest Expression Variation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA. Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Hortobagyi, G.N. Treatment of Breast Cancer. N. Engl. J. Med. 1998, 339, 974–984. [Google Scholar] [CrossRef]
- Trocchi, P.; Kuss, O.; Kääb-Sanyal, V.; Heidinger, O.; Stang, A. Trends in surgical treatment for breast cancer in Germany after the implementation of the mammography screening program. Eur. J. Epidemiol. 2019, 34, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Amato, B.; Rispoli, C.; Iannone, L.; Testa, S.; Compagna, R.; Rocco, N. Surgical margins of resection for breast cancer: Current evidence. Minerva Chir. 2012, 67, 445–452. [Google Scholar] [PubMed]
- Waldmann, A.; Hübner, J.; Katalinic, A. Trends over Time in Breast-Cancer-Specific Mortality in Germany. Dtsch. Arztebl. Int. 2021, 118, 538. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Barone, C.; Steigelman, M.; Kahlenberg, M.; Rousseau, D.; Berger, J.; Daum, A.; Ortegon, D. Aesthetic outcomes in breast conservation therapy. Aesthetic Surg. J. 2008, 28, 165–170. [Google Scholar] [CrossRef]
- Serletti, J.M.; Fosnot, J.; Nelson, J.A.; Disa, J.J.; Bucky, L.P. Breast Reconstruction after Breast Cancer. Plast. Reconstr. Surg. 2011, 127, 124e–135e. [Google Scholar] [CrossRef] [PubMed]
- Nahabedian, M.Y. Large-Volume Autologous Fat Grafting to the Breast. Aesthetic Surg. J. 2021, 41, S16–S24. [Google Scholar] [CrossRef]
- Li, X.; Meng, M.; Zhao, J.; Zhang, X.; Yang, D.; Fang, J.; Wang, J.; Han, L.; Hao, Y. Shared Decision-Making in Breast Reconstruction for Breast Cancer Patients: A Scoping Review. Patient Prefer. Adherence 2021, 15, 2763–2781. [Google Scholar] [CrossRef]
- Castaneda, S.A.; Strasser, J. Updates in the Treatment of Breast Cancer with Radiotherapy. Surg. Oncol. Clin. N. Am. 2017, 26, 371–382. [Google Scholar] [CrossRef]
- Wei, N.-N.; Li, F.; Cai, P.; Yin, H.-M.; Zhu, C.-M.; Zhang, Q.; Li, D.-J. Progress of clinical study on hypofractionated radiotherapy after breast-conserving surgery. Ann. Palliat. Med. 2020, 9, 463–471. [Google Scholar] [CrossRef]
- Early Breast Cancer Trialists’ Collaborative Group (EBCTCG). Effects of chemotherapy and hormonal therapy for early breast cancer on recurrence and 15-year survival: An overview of the randomised trials. Lancet 2005, 365, 1687–1717. [Google Scholar] [CrossRef]
- Goldhirsch, A.; Wood, W.C.; Coates, A.S.; Gelber, R.D.; Thürlimann, B.; Senn, H.-J. Panel members Strategies for subtypes—dealing with the diversity of breast cancer: Highlights of the St. Gallen International Expert Consensus on the Primary Therapy of Early Breast Cancer 2011. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2011, 22, 1736–1747. [Google Scholar] [CrossRef]
- Fisusi, F.A.; Akala, E.O. Drug Combinations in Breast Cancer Therapy. Pharm. Nanotechnol. 2019, 7, 3–23. [Google Scholar] [CrossRef]
- Bradley, R.; Braybrooke, J.; Gray, R.; Hills, R.; Liu, Z.; Peto, R.; Davies, L.; Dodwell, D.; McGale, P.; Pan, H.; et al. Trastuzumab for early-stage, HER2-positive breast cancer: A meta-analysis of 13,864 women in seven randomised trials. Lancet Oncol. 2021, 22, 1139–1150. [Google Scholar] [CrossRef]
- Osborne, C.K. Tamoxifen in the Treatment of Breast Cancer. N. Engl. J. Med. 1998, 339, 1609–1618. [Google Scholar] [CrossRef]
- Jang, H.; Baek, J.; Nam, K.-S.; Kim, S. Determination of the optimal time for tamoxifen treatment in combination with radiotherapy. Int. J. Oncol. 2016, 49, 2147–2154. [Google Scholar] [CrossRef]
- Horwitz, K.B.; Jackson, T.A.; Bain, D.L.; Richer, J.K.; Takimoto, G.S.; Tung, L. Nuclear receptor coactivators and corepressors. Mol. Endocrinol. 1996, 10, 1167–1177. [Google Scholar]
- Ellis, P.A.; Saccani-Jotti, G.; Clarke, R.; Johnston, S.R.D.; Anderson, E.; Howell, A.; A’hern, R.; Salter, J.; Detre, S.; Nicholson, R.; et al. Induction of apoptosis by tamoxifen and ICI 182780 in primary breast cancer. Int. J. Cancer 1997, 72, 608–613. [Google Scholar] [CrossRef]
- DeCensi, A.; Puntoni, M.; Guerrieri-Gonzaga, A.; Caviglia, S.; Avino, F.; Cortesi, L.; Taverniti, C.; Pacquola, M.G.; Falcini, F.; Gulisano, M.; et al. Randomized Placebo Controlled Trial of Low-Dose Tamoxifen to Prevent Local and Contralateral Recurrence in Breast Intraepithelial Neoplasia. J. Clin. Oncol. 2019, 37, 1629–1637. [Google Scholar] [CrossRef]
- Eisen, A.; Fletcher, G.G.; Gandhi, S.; Mates, M.; Freedman, O.C.; Dent, S.F.; Trudeau, M.E. Optimal Systemic Therapy for Early Breast Cancer in Women: A Clinical Practice Guideline. Curr. Oncol. 2015, 22, 67–81. [Google Scholar] [CrossRef] [Green Version]
- Early Breast Cancer Trialists’ Collaborative Group (EBCTCG); Davies, C.; Godwin, J.; Gray, R.; Clarke, M.; Cutter, D.; Darby, S.; McGale, P.; Pan, H.C.; Taylor, C.; et al. Relevance of breast cancer hormone receptors and other factors to the efficacy of adjuvant tamoxifen: Patient-level meta-analysis of randomised trials. Lancet 2011, 378, 771–784. [Google Scholar]
- Todorova, V.K.; Kaufmann, Y.; Luo, S.; Klimberg, V.S. Tamoxifen and raloxifene suppress the proliferation of estrogen receptor-negative cells through inhibition of glutamine uptake. Cancer Chemother. Pharmacol. 2011, 67, 285–291. [Google Scholar] [CrossRef]
- Barnea, Y.; Bracha, G.; Arad, E.; Gur, E.; Inbal, A. Breast Reduction and Mastopexy for Repair of Asymmetry After Breast Conservation Therapy: Lessons Learned. Aesthetic Plast. Surg. 2019, 43, 600–607. [Google Scholar] [CrossRef]
- Brown, A.W.W.; Kabir, M.; Sherman, K.A.; Meybodi, F.; French, J.R.; Elder, E.B. Patient reported outcomes of autologous fat grafting after breast cancer surgery. Breast 2017, 35, 14–20. [Google Scholar] [CrossRef]
- Caviggioli, F.; Maione, L.; Klinger, F.; Lisa, A.; Klinger, M. Autologous Fat Grafting Reduces Pain in Irradiated Breast: A Review of Our Experience. Stem Cells Int. 2016, 2016, 1–5. [Google Scholar] [CrossRef]
- Delay, E.; Garson, S.; Tousson, G.; Sinna, R. Fat Injection to the Breast: Technique, Results, and Indications Based on 880 Procedures Over 10 Years. Aesthetic Surg. J. 2009, 29, 360–376. [Google Scholar] [CrossRef] [PubMed]
- Petit, J.Y.; Botteri, E.; Lohsiriwat, V.; Rietjens, M.; De Lorenzi, F.; Garusi, C.; Rossetto, F.; Martella, S.; Manconi, A.; Bertolini, F.; et al. Locoregional recurrence risk after lipofilling in breast cancer patients. Ann. Oncol. 2012, 23, 582–588. [Google Scholar] [CrossRef]
- Bielli, A.; Scioli, M.G.; Gentile, P.; Agostinelli, S.; Tarquini, C.; Cervelli, V.; Orlandi, A. Adult adipose-derived stem cells and breast cancer: A controversial relationship. Springerplus 2014, 3, 345. [Google Scholar] [CrossRef] [PubMed]
- Naderi, N.; Combellack, E.J.; Griffin, M.; Sedaghati, T.; Javed, M.; Findlay, M.W.; Wallace, C.G.; Mosahebi, A.; Butler, P.E.; Seifalian, A.M.; et al. The regenerative role of adipose-derived stem cells (ADSC) in plastic and reconstructive surgery. Int. Wound J. 2017, 14, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Kuhbier, J.W.; Weyand, B.; Radtke, C.; Vogt, P.M.; Kasper, C.; Reimers, K. Isolation, Characterization, Differentiation, and Application of Adipose-Derived Stem Cells. In Bioreactor Systems for Tissue Engineering II; Springer: Berlin/Heidelberg, Germany, 2010; pp. 55–105. [Google Scholar]
- Schlottmann, F.; Strauss, S.; Plaass, C.; Welke, B.; Vogt, P.M.; Kuhbier, J.W. Spider Silk-Augmented Scaffolds and Adipose-Derived Stromal Cells Loaded with Uniaxial Cyclic Strain: First Investigations of a Novel Approach for Tendon-Like Constructs. Appl. Sci. 2021, 11, 1218. [Google Scholar] [CrossRef]
- Strawford, A.; Antelo, F.; Christiansen, M.; Hellerstein, M.K. Adipose tissue triglyceride turnover, de novo lipogenesis, and cell proliferation in humans measured with 2H2O. Am. J. Physiol. Metab. 2004, 286, E577–E588. [Google Scholar]
- Lauvrud, A.T.; Kelk, P.; Wiberg, M.; Kingham, P.J. Characterization of human adipose tissue-derived stem cells with enhanced angiogenic and adipogenic properties. J. Tissue Eng. Regen. Med. 2017, 11, 2490–2502. [Google Scholar] [CrossRef]
- Traktuev, D.O.; Merfeld-Clauss, S.; Li, J.; Kolonin, M.; Arap, W.; Pasqualini, R.; Johnstone, B.H.; March, K.L. A Population of Multipotent CD34-Positive Adipose Stromal Cells Share Pericyte and Mesenchymal Surface Markers, Reside in a Periendothelial Location, and Stabilize Endothelial Networks. Circ. Res. 2008, 102, 77–85. [Google Scholar] [CrossRef]
- Kølle, S.-F.T.; Fischer-Nielsen, A.; Mathiasen, A.B.; Elberg, J.J.; Oliveri, R.S.; Glovinski, P.V.; Kastrup, J.; Kirchhoff, M.; Rasmussen, B.S.; Talman, M.-L.M.; et al. Enrichment of autologous fat grafts with ex-vivo expanded adipose tissue-derived stem cells for graft survival: A randomised placebo-controlled trial. Lancet 2013, 382, 1113–1120. [Google Scholar] [CrossRef]
- Miller, W.R.; Mullen, P.; Telford, J.; Dixon, J.M. Clinical importance of intratumoral aromatase. Breast Cancer Res. Treat. 1998, 49, S27–S32. [Google Scholar] [CrossRef]
- Sasaki, Y.; Miki, Y.; Hirakawa, H.; Onodera, Y.; Takagi, K.; Akahira, J.; Honma, S.; Ishida, T.; Watanabe, M.; Sasano, H.; et al. Immunolocalization of estrogen-producing and metabolizing enzymes in benign breast disease: Comparison with normal breast and breast carcinoma. Cancer Sci. 2010, 101, 2286–2292. [Google Scholar] [CrossRef] [PubMed]
- Walter, M.; Liang, S.; Ghosh, S.; Hornsby, P.J.; Li, R. Interleukin 6 secreted from adipose stromal cells promotes migration and invasion of breast cancer cells. Oncogene 2009, 28, 2745–2755. [Google Scholar] [CrossRef]
- Mandlekar, S.; Kong, A.N. Mechanisms of tamoxifen-induced apoptosis. Apoptosis 2001, 6, 469–477. [Google Scholar] [CrossRef]
- Pike, S.; Zhang, P.; Wei, Z.; Wu, N.; Klinger, A.; Chang, S.; Jones, R.; Carpenter, J.; Brown, S.A.; DiMuzio, P.; et al. In vitro effects of tamoxifen on adipose-derived stem cells. Wound Repair Regen. 2015, 23, 728–736. [Google Scholar] [CrossRef]
- Boemi, I.; Lisa, A.V.E.; Vitali, E.; Liman, N.; Battistini, A.; Barbera, F.; Maione, L.; Vinci, V.; Klinger, M.E.A.; Lania, A.G.A. Evaluation of the ex vivo Effects of Tamoxifen on Adipose-Derived Stem Cells: A Pilot Study. Front. Cell Dev. Biol. 2021, 9, 555248. [Google Scholar] [CrossRef] [PubMed]
- Lasfargues, E.Y.; Coutinho, W.G.; Redfield, E.S. Isolation of two human tumor epithelial cell lines from solid breast carcinomas. J. Natl. Cancer Inst. 1978, 61, 967–978. [Google Scholar] [PubMed]
- Cuzick, J. The ATAC trial: The vanguard trial for use of aromatase inhibitors in early breast cancer. Expert Rev. Anticancer Ther. 2007, 7, 1089–1094. [Google Scholar] [CrossRef] [PubMed]
- Buckley, M.M.; Goa, K.L. Tamoxifen. A reappraisal of its pharmacodynamic and pharmacokinetic properties, and therapeutic use. Drugs 1989, 37, 451–490. [Google Scholar] [CrossRef]
- Yang, G.; Nowsheen, S.; Aziz, K.; Georgakilas, A.G. Toxicity and adverse effects of Tamoxifen and other anti-estrogen drugs. Pharmacol. Ther. 2013, 139, 392–404. [Google Scholar] [CrossRef]
- Lien, E.A.; Solheim, E.; Ueland, P.M. Distribution of tamoxifen and its metabolites in rat and human tissues during steady-state treatment. Cancer Res. 1991, 51, 4837–4844. [Google Scholar]
- Michalets, E.L. Update: Clinically significant cytochrome P-450 drug interactions. Pharmacotherapy 1998, 18, 84–112. [Google Scholar] [CrossRef]
- Guillaume, V.G.J.; Ruhl, T.; Boos, A.M.; Beier, J.P. The Crosstalk Between Adipose-Derived Stem or Stromal Cells (ASC) and Cancer Cells and ASC-Mediated Effects on Cancer Formation and Progression—ASCs: Safety Hazard or Harmless Source of Tropism? Stem Cells Transl. Med. 2022, 11, 394–406. [Google Scholar] [CrossRef]
- Varghese, J.; Griffin, M.; Mosahebi, A.; Butler, P. Systematic review of patient factors affecting adipose stem cell viability and function: Implications for regenerative therapy. Stem Cell Res. Ther. 2017, 8, 45. [Google Scholar] [CrossRef]
- Kim, J.; Choi, Y.S.; Lim, S.; Yea, K.; Yoon, J.H.; Jun, D.-J.; Ha, S.H.; Kim, J.-W.; Kim, J.H.; Suh, P.-G.; et al. Comparative analysis of the secretory proteome of human adipose stromal vascular fraction cells during adipogenesis. Proteomics 2010, 10, 394–405. [Google Scholar] [CrossRef]
- Robinson, J.L.; Feizi, A.; Uhlén, M.; Nielsen, J. A Systematic Investigation of the Malignant Functions and Diagnostic Potential of the Cancer Secretome. Cell Rep. 2019, 26, 2622–2635.e5. [Google Scholar] [CrossRef]
- Kengelbach-Weigand, A.; Tasbihi, K.; Strissel, P.L.; Schmid, R.; Marques, J.M.; Beier, J.P.; Beckmann, M.W.; Strick, R.; Horch, R.E.; Boos, A.M. Plasticity of patient-matched normal mammary epithelial cells is dependent on autologous adipose-derived stem cells. Sci. Rep. 2019, 9, 10722. [Google Scholar] [CrossRef]
- Koellensperger, E.; Bonnert, L.-C.; Zoernig, I.; Marmé, F.; Sandmann, S.; Germann, G.; Gramley, F.; Leimer, U. The impact of human adipose tissue-derived stem cells on breast cancer cells: Implications for cell-assisted lipotransfers in breast reconstruction. Stem Cell Res. Ther. 2017, 8, 121. [Google Scholar] [CrossRef]
- Trivanović, D.; Nikolić, S.; Krstić, J.; Jauković, A.; Mojsilović, S.; Ilić, V.; Okić-Djordjević, I.; Santibanez, J.F.; Jovčić, G.; Bugarski, D. Characteristics of human adipose mesenchymal stem cells isolated from healthy and cancer affected people and their interactions with human breast cancer cell line MCF-7 in vitro. Cell Biol. Int. 2014, 38, 254–265. [Google Scholar] [CrossRef]
- Sun, B.; Roh, K.-H.; Park, J.-R.; Lee, S.-R.; Park, S.-B.; Jung, J.-W.; Kang, S.-K.; Lee, Y.-S.; Kang, K.-S. Therapeutic potential of mesenchymal stromal cells in a mouse breast cancer metastasis model. Cytotherapy 2009, 11, 289–298. [Google Scholar] [CrossRef]
- Hass, R.; Otte, A. Mesenchymal stem cells as all-round supporters in a normal and neoplastic microenvironment. Cell Commun. Signal. 2012, 10, 26. [Google Scholar] [CrossRef]
- Donahue, H.J.; Saunders, M.M.; Li, Z.; Mastro, A.M.; Gay, C.V.; Welch, D.R. A potential role for gap junctions in breast cancer metastasis to bone. J. Musculoskelet. Neuronal Interact. 2003, 3, 156–161. [Google Scholar]
- Kuhbier, J.W.; Bucan, V.; Reimers, K.; Strauss, S.; Lazaridis, A.; Jahn, S.; Radtke, C.; Vogt, P.M. Observed Changes in the Morphology and Phenotype of Breast Cancer Cells in Direct Co-Culture with Adipose-Derived Stem Cells. Plast. Reconstr. Surg. 2014, 134, 414–423. [Google Scholar] [CrossRef]
- Mandel, K.; Yang, Y.; Schambach, A.; Glage, S.; Otte, A.; Hass, R. Mesenchymal Stem Cells Directly Interact with Breast Cancer Cells and Promote Tumor Cell Growth In Vitro and In Vivo. Stem Cells Dev. 2013, 22, 3114–3127. [Google Scholar] [CrossRef]
- Soule, H.D.; Maloney, T.M.; Wolman, S.R.; Peterson, W.D.; Brenz, R.; McGrath, C.M.; Russo, J.; Pauley, R.J.; Jones, R.F.; Brooks, S.C. Isolation and characterization of a spontaneously immortalized human breast epithelial cell line, MCF-10. Cancer Res. 1990, 50, 6075–6086. [Google Scholar]
- Hodges, V.M.; Rainey, S.; Lappin, T.R.; Maxwell, A.P. Pathophysiology of anemia and erythrocytosis. Crit. Rev. Oncol. Hematol. 2007, 64, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Leyland-Jones, B.; Semiglazov, V.; Pawlicki, M.; Pienkowski, T.; Tjulandin, S.; Manikhas, G.; Makhson, A.; Roth, A.; Dodwell, D.; Baselga, J.; et al. Maintaining Normal Hemoglobin Levels With Epoetin Alfa in Mainly Nonanemic Patients With Metastatic Breast Cancer Receiving First-Line Chemotherapy: A Survival Study. J. Clin. Oncol. 2005, 23, 5960–5972. [Google Scholar] [CrossRef] [PubMed]
- Ordoñez-Moreno, A.; Rodriguez-Monterrosas, C.; Cortes-Reynosa, P.; Perez-Carreon, J.I.; Perez Salazar, E. Erythropoietin Induces an Epithelial to Mesenchymal Transition-Like Process in Mammary Epithelial Cells MCF10A. J. Cell. Biochem. 2017, 118, 2983–2992. [Google Scholar] [CrossRef] [PubMed]
- Akwii, R.G.; Sajib, M.S.; Zahra, F.T.; Mikelis, C.M. Role of Angiopoietin-2 in Vascular Physiology and Pathophysiology. Cells 2019, 8, 471. [Google Scholar] [CrossRef]
- Silva, A.B.D.; Haupenthal, F.; Morais, A.D.; Ascenço, A.S.K.; Sebastião, A.P.M.; Cavalcanti, M.A.R.; Freitas, R.S. Relationship between Tamoxifen and the Absorption of Subfascial Autologous Fat Grafts. Plast. Reconstr. Surg. 2018, 141, 1408–1415. [Google Scholar] [CrossRef]
- Green, D.R.; Droin, N.; Pinkoski, M. Activation-induced cell death in T cells. Immunol. Rev. 2003, 193, 70–81. [Google Scholar] [CrossRef]
- Brooks, S.C.; Locke, E.R.; Soule, H.D. Estrogen receptor in a human cell line (MCF-7) from breast carcinoma. J. Biol. Chem. 1973, 248, 6251–6253. [Google Scholar] [CrossRef]
- Soule, H.D.; Vazquez, J.; Long, A.; Albert, S.; Brennan, M. A Human Cell Line From a Pleural Effusion Derived From a Breast Carcinoma 2. JNCI J. Natl. Cancer Inst. 1973, 51, 1409–1416. [Google Scholar] [CrossRef]
- Chen, L.; Willis, S.N.; Wei, A.; Smith, B.J.; Fletcher, J.I.; Hinds, M.G.; Colman, P.M.; Day, C.L.; Adams, J.M.; Huang, D.C.S. Differential Targeting of Prosurvival Bcl-2 Proteins by Their BH3-Only Ligands Allows Complementary Apoptotic Function. Mol. Cell 2005, 17, 393–403. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Trost, N.; Juvan, P.; Sersa, G.; Debeljak, N. Contrasting effect of recombinant human erythropoietin on breast cancer cell response to cisplatin induced cytotoxicity. Radiol. Oncol. 2012, 46, 213. [Google Scholar] [CrossRef] [PubMed]
- Sureshbabu, A.; Okajima, H.; Yamanaka, D.; Tonner, E.; Shastri, S.; Maycock, J.; Szymanowska, M.; Shand, J.; Takahashi, S.-I.; Beattie, J.; et al. IGFBP-5 induces cell adhesion, increases cell survival and inhibits cell migration in MCF-7 human breast cancer cells. J. Cell Sci. 2012, 125, 1693–1705. [Google Scholar] [CrossRef] [PubMed]
- Slamon, D.J.; Clark, G.M.; Wong, S.G.; Levin, W.J.; Ullrich, A.; McGuire, W.L. Human Breast Cancer: Correlation of Relapse and Survival with Amplification of the HER-2/neu Oncogene. Science 1987, 235, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Colomer, R.; Montero, S.; Lluch, A.; Ojeda, B.; Barnadas, A.; Casado, A.; Massutí, B.; Cortés-Funes, H.; Lloveras, B. Circulating HER2 extracellular domain and resistance to chemotherapy in advanced breast cancer. Clin. Cancer Res. 2000, 6, 2356–2362. [Google Scholar]
- Newby, J.C.; Johnston, S.R.; Smith, I.E.; Dowsett, M. Expression of epidermal growth factor receptor and c-erbB2 during the development of tamoxifen resistance in human breast cancer. Clin. Cancer Res. 1997, 3, 1643–1651. [Google Scholar]
- Ropero, S.; Abel Menéndez, J.; Vázquez-Martín, A.; Montero, S.; Cortés-Funes, H.; Colomer, R. Trastuzumab Plus Tamoxifen: Anti-Proliferative and Molecular Interactions in Breast Carcinoma. Breast Cancer Res. Treat. 2004, 86, 125–137. [Google Scholar] [CrossRef]
- Li, T.; Su, L.; Lei, Y.; Liu, X.; Zhang, Y.; Liu, X. DDIT3 and KAT2A Proteins Regulate TNFRSF10A and TNFRSF10B Expression in Endoplasmic Reticulum Stress-mediated Apoptosis in Human Lung Cancer Cells. J. Biol. Chem. 2015, 290, 11108–11118. [Google Scholar] [CrossRef]
- Zhang, C.; Duan, X.; Xu, L.; Ye, J.; Zhao, J.; Liu, Y. Erythropoietin receptor expression and its relationship with trastuzumab response and resistance in HER2-positive breast cancer cells. Breast Cancer Res. Treat. 2012, 136, 739–748. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function | Gene |
|---|---|
| Angiogenesis | Angiopoetin 1 (ANGPT1) Angiopoetin 2 (ANGPT2) Chemokine (C-C motif) ligand 2 (CCL2) Basic fibroblastic growth factor 2 (FGF2) Kinase insert domain receptor (KDR) Vascular endothelial growth factor receptor 1 (VEGFR1) Vascular endothelial growth factor 3 (VEGFR3) Placental growth factor (PGF) Serpin peptidase inhibitor, clade F (alpha-2 antiplasmin, pigment epithelium derived factor), member 1 (SERPINF1) TEK tyrosine kinase, endothelial (TEK) Vascular endothelial growth factor C (VEGFC) |
| Apoptosis | Apoptotic peptidase activating factor 1 (APAF1) B-cell lymphoma 2 like 11 (apoptosis facilitator) (BCL2L11) Baculoviral inhibitor of apoptosis repeat-containing protein 3 (BIRC3) Caspase 2, apoptosis-related cysteine peptidase (CASP2) Caspase 7, apoptosis-related cysteine peptidase (CASP7) Caspase 9, apoptosis-related cysteine peptidase (CASP9) Caspase 8 und fas-associating death domain-containing protein-like apoptosis regulator (CFLAR) Fas- ligand (TNF superfamily, member 6) (FASLG) Nucleolar protein 3 (apoptosis repressor with CARD domain) (NOL3) X-linked inhibitor of apoptosis (XIAP) |
| Cell cycle | Aurora kinase A (AURKA) Cyclin D2 (CCND2) Cyclin D3 (CCND3) Cell division cycle 20 homolog (S. cerevisiae) (CDC20) E2F transcription factor 4, p107/p130-binding (E2F4) FMS related tyrosine kinase 1 (FLT1) Minichromosome maintenance complex component 2 (MCM2) Antigen identified by monoclonal antibody Ki-67 (MKI67) S-phase kinase-associated protein 2 (p45) (SKP2) Stathmin 1 (STMN1) WEE homolog (S.pombe) (WEE1) |
| Cellular Senescence | BMI1 polycomb ring finger oncogene (BMI1) V-Ets erythroblastosis virus E26 oncogene homolog 2 (avian) (ETS2) Insulin-like growth factor binding protein 3 (IGFBP3) Insulin-like growth factor binding protein 5 (IGFBP5) Insulin-like growth factor binding protein 7 (IGFBP7) Mitogen-activated protein kinase 1 (MAP2K1) Mitogen-activated protein kinase 3 (MAP2K3) Mitogen-activated protein kinase 14 (MAPK14) Serpin peptidase inhibitor, clade B (ovalbumin), member 2 (SERPINB2) Superoxidase dismutase 1, soluble (SOD1) T-box 2 (TBX2) |
| DNA Damage and Repair | Damage-specific DNA binding protein 2, 48kDa (DDB2) DNA-damage-inducible transcript 3 (DDIT3) Excision repair cross-complementing rodent repair deficiency, complementation group 3 (ERCC3) Excision repair cross-complementing rodent repair deficiency, complementation group 5 (ERCC5) Growth arrest and DNA-damage-inducible, gamma (GADD45G) Ligase IV, DNA, ATP-dependent (LIG4) Polymerase (DNA directed), beta (POLB) Protein phosphatase 1, regulatory (inhibitor) subunit 15A (PPP1R15A) |
| Epithelial-to-Mesenchymal Transition | Cadherin 2, type 1, N-cadherin (neuronal) (CDH2) Desmoplakin (DSP) Forkhead box C2 (MFH-1, mesenchyme forkhead 1) (FOXC2) Goosecoid homeobox (GSC) Keratin 14 (KRT14) Occludin (OCLN) Snail homolog 1 (Drosophila) (SNAI1) Snail homolog 2 (Drosophila) (SNAI2) Snail homolog 3 (Drosophila) (SNAI3) SRY (sex determining region Y)-box 10 (SOX10) |
| Hypoxia Signaling | Adrenomedullin (ADM) Aryl hydrocarbon receptor nuclear translocator (ARNT) Carbonic anhydrase IX (CA9) Erythropoietin (EPO) Heme oxygenase (decycling) 1 (HMOX1) Lactate dehydrogenase A (LDHA) Solute carrier family 2 (facilitated glucose transporter), member 1 (SLC2A1) |
| Metabolism | ATP citrate lyase (ACLY)Acyl-CoA synthetase long-chain family member 4 (ACSL4) ATP synthase, H+ transporting, mitochondrial F1 complex, alpha subunit 1, cardiac muscle (ATP5A1) Cytochrome c oxidase subunit Va (COX5A) Carnitine palmitoyltransferase 2 (CPT2) Glucose-6-phosphate dehydrogenase (G6PD) Glycerol-3-phosphate dehydrogenase 2 (mitochondrial) (GPD2) Lipoprotein lipase (LPL) Phosphofructokinase, liver (PFKL) Ubiquinol-cytochrome c reductase, Rieske iron-sulfur polypeptide 1 (UQCRFS1) |
| Telomeres and Telomerase | Dyskeratosis congenita 1, dyskerin (DKC1) PIN2/TERF1 interacting, telomerase inhibitor 1 (PINX1) Telomerase-associated protein 1 (TEP1) Telomeric repeat binding factor (NIMA-interacting 1) (TERF1) Telomeric repeat binding factor 2, interacting protein (TERF2IP) TERF1 (TRF1)-interacting nuclear factor 2 (TINF2) Tankyrase, TRF1-interacting ankyrin-related ADP-ribose polymerase (TNKS) Tankyrase, TRF1-interacting ankyrin-related ADP-ribose polymerase 2 (TNKS2) |
| Function | Gene |
|---|---|
| Angiogenesis | ANGPT1 |
| Apoptosis | BCL2L11 CASP2 FASLG |
| Cell Cycle | FLT1 |
| Cellular Senescence | IGFBP5 |
| DNA Damage and Repair | DDIT3 |
| Epithelial-to-Mesenchymal Transition | KRT14 OCLN SNAI1 SNAI3 |
| Hypoxia Signaling | EPO |
| Metabolism | −/− |
| Telomeres and Telomerase | TEP1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schlottmann, F.; Bucan, V.; Strauß, S.; Koop, F.; Vogt, P.M.; Mett, T.R. Influence of Tamoxifen on Different Biological Pathways in Tumorigenesis and Transformation in Adipose-Derived Stem Cells, Mammary Cells and Mammary Carcinoma Cell Lines—An In Vitro Study. Cells 2022, 11, 2733. https://doi.org/10.3390/cells11172733
Schlottmann F, Bucan V, Strauß S, Koop F, Vogt PM, Mett TR. Influence of Tamoxifen on Different Biological Pathways in Tumorigenesis and Transformation in Adipose-Derived Stem Cells, Mammary Cells and Mammary Carcinoma Cell Lines—An In Vitro Study. Cells. 2022; 11(17):2733. https://doi.org/10.3390/cells11172733
Chicago/Turabian StyleSchlottmann, Frederik, Vesna Bucan, Sarah Strauß, Felix Koop, Peter M. Vogt, and Tobias R. Mett. 2022. "Influence of Tamoxifen on Different Biological Pathways in Tumorigenesis and Transformation in Adipose-Derived Stem Cells, Mammary Cells and Mammary Carcinoma Cell Lines—An In Vitro Study" Cells 11, no. 17: 2733. https://doi.org/10.3390/cells11172733