
Recent Advances in the Modulation of Pain by the Metabotropic Glutamate Receptors

Abstract
:1. Introduction
2. mGluRs and Clinical Trials in The Context of Pain
3. Group I mGluRs in Preclinical Pain Models
3.1. Preclinical Behavioral Studies
3.2. Preclinical Electrophysiological Studies
4. Group II mGluRs in Preclinical Pain Models
4.1. Preclinical Behavioral Studies
4.2. Preclinical Electrophysiological Studies
5. Group III mGluRs in Preclinical Pain Models
5.1. Preclinical Behavioral Studies
5.2. Preclinical Electrophysiological Studies
6. Role of mGluRs in Neuroimmune Signaling
7. Sex Differences and mGluRs
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mills, S.E.E.; Nicolson, K.P.; Smith, B.H. Chronic pain: A review of its epidemiology and associated factors in population-based studies. Br. J. Anaesth. 2019, 123, e273–e283. [Google Scholar] [CrossRef] [PubMed]
- Colloca, L.; Ludman, T.; Bouhassira, D.; Baron, R.; Dickenson, A.H.; Yarnitsky, D.; Freeman, R.; Truini, A.; Attal, N.; Finnerup, N.B.; et al. Neuropathic pain. Nat. Rev. Dis. Primers 2017, 3, 17002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spooren, W.; Ballard, T.; Gasparini, F.; Amalric, M.; Mutel, V.; Schreiber, R. Insight into the function of Group I and Group II metabotropic glutamate (mGlu) receptors: Behavioural characterization and implications for the treatment of CNS disorders. Behav. Pharmacol. 2003, 14, 257–277. [Google Scholar] [CrossRef] [PubMed]
- Neugebauer, V. Glutamate receptor ligands. Handb. Exp. Pharmacol. 2007, 217–249. [Google Scholar] [CrossRef]
- Chiechio, S. Modulation of Chronic Pain by Metabotropic Glutamate Receptors. Adv. Pharmacol. 2016, 75, 63–89. [Google Scholar] [CrossRef]
- Pereira, V.; Goudet, C. Emerging Trends in Pain Modulation by Metabotropic Glutamate Receptors. Front. Mol. Neurosci. 2018, 11, 464. [Google Scholar] [CrossRef]
- Niswender, C.M.; Conn, P.J. Metabotropic glutamate receptors: Physiology, pharmacology, and disease. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 295–322. [Google Scholar] [CrossRef] [Green Version]
- Celli, R.; Santolini, I.; Van Luijtelaar, G.; Ngomba, R.T.; Bruno, V.; Nicoletti, F. Targeting metabotropic glutamate receptors in the treatment of epilepsy: Rationale and current status. Expert Opin. Ther. Targets 2019, 23, 341–351. [Google Scholar] [CrossRef]
- Gass, J.T.; Olive, M.F. Glutamatergic substrates of drug addiction and alcoholism. Biochem. Pharmacol. 2008, 75, 218–265. [Google Scholar] [CrossRef] [Green Version]
- Crupi, R.; Impellizzeri, D.; Cuzzocrea, S. Role of Metabotropic Glutamate Receptors in Neurological Disorders. Front. Mol. Neurosci. 2019, 12, 20. [Google Scholar] [CrossRef] [Green Version]
- Nicoletti, F.; Bockaert, J.; Collingridge, G.L.; Conn, P.J.; Ferraguti, F.; Schoepp, D.D.; Wroblewski, J.T.; Pin, J.P. Metabotropic glutamate receptors: From the workbench to the bedside. Neuropharmacology 2011, 60, 1017–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zussy, C.; Gomez-Santacana, X.; Rovira, X.; De Bundel, D.; Ferrazzo, S.; Bosch, D.; Asede, D.; Malhaire, F.; Acher, F.; Giraldo, J.; et al. Dynamic modulation of inflammatory pain-related affective and sensory symptoms by optical control of amygdala metabotropic glutamate receptor 4. Mol. Psychiatry 2018, 23, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Mony, L.; Paoletti, P. Photocontrol of Metabotropic Glutamate Receptors: When One Agonist Is Not Enough, Make It Two. Neuron 2020, 105, 395–397. [Google Scholar] [CrossRef] [PubMed]
- Loche, A.; Simonetti, F.; Lobina, C.; Carai, M.A.; Colombo, G.; Castelli, M.P.; Barone, D.; Cacciaglia, R. Anti-Alcohol and Anxiolytic Properties of a New Chemical Entity, GET73. Front. Psychiatry 2012, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Tomasini, M.C.; Borelli, A.C.; Beggiato, S.; Tanganelli, S.; Loche, A.; Cacciaglia, R.; Ferraro, L.; Antonelli, T. GET73 Prevents Ethanol-Induced Neurotoxicity in Primary Cultures of Rat Hippocampal Neurons. Alcohol Alcohol. 2016, 51, 128–135. [Google Scholar] [CrossRef] [Green Version]
- Ottani, A.; Leone, S.; Vergara, F.B.; Tacchi, R.; Loche, A.; Bertolini, A. Preference for palatable food is reduced by the gamma-hydroxybutyrate analogue GET73, in rats. Pharmacol. Res. 2007, 55, 271–279. [Google Scholar] [CrossRef]
- Haass-Koffler, C.L.; Goodyear, K.; Long, V.M.; Tran, H.H.; Loche, A.; Cacciaglia, R.; Swift, R.M.; Leggio, L. A Phase I randomized clinical trial testing the safety, tolerability and preliminary pharmacokinetics of the mGluR5 negative allosteric modulator GET 73 following single and repeated doses in healthy volunteers. Eur. J. Pharm. Sci. 2017, 109, 78–85. [Google Scholar] [CrossRef]
- Gibert-Rahola, J.; Villena-Rodriguez, A. Glutamatergic drugs for schizophrenia treatment. Actas. Esp. Psiquiatr. 2014, 42, 234–241. [Google Scholar]
- Stauffer, V.L.; Millen, B.A.; Andersen, S.; Kinon, B.J.; Lagrandeur, L.; Lindenmayer, J.P.; Gomez, J.C. Pomaglumetad methionil: No significant difference as an adjunctive treatment for patients with prominent negative symptoms of schizophrenia compared to placebo. Schizophr. Res. 2013, 150, 434–441. [Google Scholar] [CrossRef]
- Adams, D.H.; Kinon, B.J.; Baygani, S.; Millen, B.A.; Velona, I.; Kollack-Walker, S.; Walling, D.P. A long-term, phase 2, multicenter, randomized, open-label, comparative safety study of pomaglumetad methionil (LY2140023 monohydrate) versus atypical antipsychotic standard of care in patients with schizophrenia. BMC Psychiatry 2013, 13, 143. [Google Scholar] [CrossRef] [Green Version]
- Li, M.L.; Hu, X.Q.; Li, F.; Gao, W.J. Perspectives on the mGluR2/3 agonists as a therapeutic target for schizophrenia: Still promising or a dead end? Prog. Neuropsychopharmacol. Biol. Psychiatry 2015, 60, 66–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marek, G.J. When is a Proof-of-Concept (POC) not a POC? Pomaglumetad (LY2140023) as a Case Study for Antipsychotic Efficacy. Curr. Pharm. Des. 2015, 21, 3788–3796. [Google Scholar] [CrossRef] [PubMed]
- Quiroz, J.A.; Tamburri, P.; Deptula, D.; Banken, L.; Beyer, U.; Rabbia, M.; Parkar, N.; Fontoura, P.; Santarelli, L. Efficacy and Safety of Basimglurant as Adjunctive Therapy for Major Depression: A Randomized Clinical Trial. JAMA Psychiatry 2016, 73, 675–684. [Google Scholar] [CrossRef]
- Umbricht, D.; Niggli, M.; Sanwald-Ducray, P.; Deptula, D.; Moore, R.; Grunbauer, W.; Boak, L.; Fontoura, P. Randomized, Double-Blind, Placebo-Controlled Trial of the mGlu2/3 Negative Allosteric Modulator Decoglurant in Partially Refractory Major Depressive Disorder. J. Clin. Psychiatry 2020, 81. [Google Scholar] [CrossRef] [PubMed]
- Porter, R.H.; Jaeschke, G.; Spooren, W.; Ballard, T.M.; Buttelmann, B.; Kolczewski, S.; Peters, J.U.; Prinssen, E.; Wichmann, J.; Vieira, E.; et al. Fenobam: A clinically validated nonbenzodiazepine anxiolytic is a potent, selective, and noncompetitive mGlu5 receptor antagonist with inverse agonist activity. J. Pharmacol. Exp. Ther. 2005, 315, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Montana, M.C.; Cavallone, L.F.; Stubbert, K.K.; Stefanescu, A.D.; Kharasch, E.D.; Gereau, R.W.t. The metabotropic glutamate receptor subtype 5 antagonist fenobam is analgesic and has improved in vivo selectivity compared with the prototypical antagonist 2-methyl-6-(phenylethynyl)-pyridine. J. Pharmacol. Exp. Ther. 2009, 330, 834–843. [Google Scholar] [CrossRef] [Green Version]
- Montana, M.C.; Conrardy, B.A.; Cavallone, L.F.; Kolber, B.J.; Rao, L.K.; Greco, S.C.; Gereau, R.W.t. Metabotropic glutamate receptor 5 antagonism with fenobam: Examination of analgesic tolerance and side effect profile in mice. Anesthesiology 2011, 115, 1239–1250. [Google Scholar] [CrossRef] [Green Version]
- Jacob, W.; Gravius, A.; Pietraszek, M.; Nagel, J.; Belozertseva, I.; Shekunova, E.; Malyshkin, A.; Greco, S.; Barberi, C.; Danysz, W. The anxiolytic and analgesic properties of fenobam, a potent mGlu5 receptor antagonist, in relation to the impairment of learning. Neuropharmacology 2009, 57, 97–108. [Google Scholar] [CrossRef]
- Lax, N.C.; George, D.C.; Ignatz, C.; Kolber, B.J. The mGluR5 antagonist fenobam induces analgesic conditioned place preference in mice with spared nerve injury. PLoS ONE 2014, 9, e103524. [Google Scholar] [CrossRef] [Green Version]
- Crock, L.W.; Stemler, K.M.; Song, D.G.; Abbosh, P.; Vogt, S.K.; Qiu, C.S.; Lai, H.H.; Mysorekar, I.U.; Gereau, R.W.t. Metabotropic glutamate receptor 5 (mGluR5) regulates bladder nociception. Mol. Pain 2012, 8, 20. [Google Scholar] [CrossRef] [Green Version]
- Cavallone, L.F.; Montana, M.C.; Frey, K.; Kallogjeri, D.; Wages, J.M.; Rodebaugh, T.L.; Doshi, T.; Kharasch, E.D.; Gereau, R.W.t. The metabotropic glutamate receptor 5 negative allosteric modulator fenobam: Pharmacokinetics, side effects, and analgesic effects in healthy human subjects. Pain 2020, 161, 135–146. [Google Scholar] [CrossRef]
- Pardridge, W.M. The blood-brain barrier: Bottleneck in brain drug development. NeuroRx 2005, 2, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Brooks, T.A.; Hawkins, B.T.; Huber, J.D.; Egleton, R.D.; Davis, T.P. Chronic inflammatory pain leads to increased blood-brain barrier permeability and tight junction protein alterations. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H738–H743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, J.D.; Witt, K.A.; Hom, S.; Egleton, R.D.; Mark, K.S.; Davis, T.P. Inflammatory pain alters blood-brain barrier permeability and tight junctional protein expression. Am. J. Physiol. Heart Circ. Physiol. 2001, 280, H1241–H1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronaldson, P.T.; Davis, T.P. Targeting blood-brain barrier changes during inflammatory pain: An opportunity for optimizing CNS drug delivery. Ther. Deliv. 2011, 2, 1015–1041. [Google Scholar] [CrossRef] [Green Version]
- Viscusi, E.R.; Viscusi, A.R. Blood-brain barrier: Mechanisms governing permeability and interaction with peripherally acting mu-opioid receptor antagonists. Reg. Anesth. Pain Med. 2020, 45, 688–695. [Google Scholar] [CrossRef]
- Sevostianova, N.; Danysz, W. Analgesic effects of mGlu1 and mGlu5 receptor antagonists in the rat formalin test. Neuropharmacology 2006, 51, 623–630. [Google Scholar] [CrossRef]
- Satow, A.; Maehara, S.; Ise, S.; Hikichi, H.; Fukushima, M.; Suzuki, G.; Kimura, T.; Tanak, T.; Ito, S.; Kawamoto, H.; et al. Pharmacological effects of the metabotropic glutamate receptor 1 antagonist compared with those of the metabotropic glutamate receptor 5 antagonist and metabotropic glutamate receptor 2/3 agonist in rodents: Detailed investigations with a selective allosteric metabotropic glutamate receptor 1 antagonist, FTIDC [4-[1-(2-fluoropyridine-3-yl)-5-methyl-1H-1,2,3-triazol-4-yl]-N-isopropyl-N-methy l-3,6-dihydropyridine-1(2H)-carboxamide]. J. Pharmacol. Exp. Ther. 2008, 326, 577–586. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.Z.; Baker, S.; EI-Kouhen, O.; Lehto, S.G.; Hollingsworth, P.R.; Gauvin, D.M.; Hernandez, G.; Zheng, G.; Chang, R.; Moreland, R.B.; et al. Analgesic activity of metabotropic glutamate receptor 1 antagonists on spontaneous post-operative pain in rats. Eur. J. Pharmacol. 2008, 580, 314–321. [Google Scholar] [CrossRef]
- Varty, G.B.; Grilli, M.; Forlani, A.; Fredduzzi, S.; Grzelak, M.E.; Guthrie, D.H.; Hodgson, R.A.; Lu, S.X.; Nicolussi, E.; Pond, A.J.; et al. The antinociceptive and anxiolytic-like effects of the metabotropic glutamate receptor 5 (mGluR5) antagonists, MPEP and MTEP, and the mGluR1 antagonist, LY456236, in rodents: A comparison of efficacy and side-effect profiles. Psychopharmacology 2005, 179, 207–217. [Google Scholar] [CrossRef]
- El-Kouhen, O.; Lehto, S.G.; Pan, J.B.; Chang, R.; Baker, S.J.; Zhong, C.; Hollingsworth, P.R.; Mikusa, J.P.; Cronin, E.A.; Chu, K.L.; et al. Blockade of mGluR1 receptor results in analgesia and disruption of motor and cognitive performances: Effects of A-841720, a novel non-competitive mGluR1 receptor antagonist. Br. J. Pharmacol. 2006, 149, 761–774. [Google Scholar] [CrossRef] [PubMed]
- Walker, K.; Bowes, M.; Panesar, M.; Davis, A.; Gentry, C.; Kesingland, A.; Gasparini, F.; Spooren, W.; Stoehr, N.; Pagano, A.; et al. Metabotropic glutamate receptor subtype 5 (mGlu5) and nociceptive function. I. Selective blockade of mGlu5 receptors in models of acute, persistent and chronic pain. Neuropharmacology 2001, 40, 1–9. [Google Scholar] [CrossRef]
- Zhu, C.Z.; Wilson, S.G.; Mikusa, J.P.; Wismer, C.T.; Gauvin, D.M.; Lynch, J.J., 3rd; Wade, C.L.; Decker, M.W.; Honore, P. Assessing the role of metabotropic glutamate receptor 5 in multiple nociceptive modalities. Eur. J. Pharmacol. 2004, 506, 107–118. [Google Scholar] [CrossRef]
- Hudson, L.J.; Bevan, S.; McNair, K.; Gentry, C.; Fox, A.; Kuhn, R.; Winter, J. Metabotropic glutamate receptor 5 upregulation in A-fibers after spinal nerve injury: 2-methyl-6-(phenylethynyl)-pyridine (MPEP) reverses the induced thermal hyperalgesia. J. Neurosci. 2002, 22, 2660–2668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osikowicz, M.; Mika, J.; Makuch, W.; Przewlocka, B. Glutamate receptor ligands attenuate allodynia and hyperalgesia and potentiate morphine effects in a mouse model of neuropathic pain. Pain 2008, 139, 117–126. [Google Scholar] [CrossRef]
- Lindstrom, E.; Brusberg, M.; Hughes, P.A.; Martin, C.M.; Brierley, S.M.; Phillis, B.D.; Martinsson, R.; Abrahamsson, C.; Larsson, H.; Martinez, V.; et al. Involvement of metabotropic glutamate 5 receptor in visceral pain. Pain 2008, 137, 295–305. [Google Scholar] [CrossRef]
- Walker, K.; Reeve, A.; Bowes, M.; Winter, J.; Wotherspoon, G.; Davis, A.; Schmid, P.; Gasparini, F.; Kuhn, R.; Urban, L. mGlu5 receptors and nociceptive function II. mGlu5 receptors functionally expressed on peripheral sensory neurones mediate inflammatory hyperalgesia. Neuropharmacology 2001, 40, 10–19. [Google Scholar] [CrossRef]
- Jin, Y.H.; Yamaki, F.; Takemura, M.; Koike, Y.; Furuyama, A.; Yonehara, N. Capsaicin-induced glutamate release is implicated in nociceptive processing through activation of ionotropic glutamate receptors and group I metabotropic glutamate receptor in primary afferent fibers. J. Pharmacol. Sci. 2009, 109, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Bhave, G.; Karim, F.; Carlton, S.M.; Gereau, R.W.t. Peripheral group I metabotropic glutamate receptors modulate nociception in mice. Nat. Neurosci. 2001, 4, 417–423. [Google Scholar] [CrossRef]
- Lee, J.S.; Ro, J.Y. Peripheral metabotropic glutamate receptor 5 mediates mechanical hypersensitivity in craniofacial muscle via protein kinase C dependent mechanisms. Neuroscience 2007, 146, 375–383. [Google Scholar] [CrossRef]
- Ahn, D.K.; Kim, K.H.; Jung, C.Y.; Choi, H.S.; Lim, E.J.; Youn, D.H.; Bae, Y.C. Role of peripheral group I and II metabotropic glutamate receptors in IL-1beta-induced mechanical allodynia in the orofacial area of conscious rats. Pain 2005, 118, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.Z.; Hsieh, G.; Ei-Kouhen, O.; Wilson, S.G.; Mikusa, J.P.; Hollingsworth, P.R.; Chang, R.; Moreland, R.B.; Brioni, J.; Decker, M.W.; et al. Role of central and peripheral mGluR5 receptors in post-operative pain in rats. Pain 2005, 114, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Font, J.; Lopez-Cano, M.; Notartomaso, S.; Scarselli, P.; Di Pietro, P.; Bresoli-Obach, R.; Battaglia, G.; Malhaire, F.; Rovira, X.; Catena, J.; et al. Optical control of pain in vivo with a photoactive mGlu5 receptor negative allosteric modulator. Elife 2017, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, K.; Coderre, T.J. Comparison of nociceptive effects produced by intrathecal administration of mGluR agonists. Neuroreport 1996, 7, 2743–2747. [Google Scholar] [CrossRef] [PubMed]
- Lorrain, D.S.; Correa, L.; Anderson, J.; Varney, M. Activation of spinal group I metabotropic glutamate receptors in rats evokes local glutamate release and spontaneous nociceptive behaviors: Effects of 2-methyl-6-(phenylethynyl)-pyridine pretreatment. Neurosci. Lett. 2002, 327, 198–202. [Google Scholar] [CrossRef]
- Hama, A.T. Acute activation of the spinal cord metabotropic glutamate subtype-5 receptor leads to cold hypersensitivity in the rat. Neuropharmacology 2003, 44, 423–430. [Google Scholar] [CrossRef]
- Fisher, K.; Coderre, T.J. Hyperalgesia and allodynia induced by intrathecal (RS)-dihydroxyphenylglycine in rats. Neuroreport 1998, 9, 1169–1172. [Google Scholar] [CrossRef]
- Radwani, H.; Roca-Lapirot, O.; Aby, F.; Lopez-Gonzalez, M.J.; Benazzouz, R.; Errami, M.; Favereaux, A.; Landry, M.; Fossat, P. Group I metabotropic glutamate receptor plasticity after peripheral inflammation alters nociceptive transmission in the dorsal of the spinal cord in adult rats. Mol. Pain 2017, 13, 1744806917737934. [Google Scholar] [CrossRef] [Green Version]
- Karim, F.; Wang, C.C.; Gereau, R.W.t. Metabotropic glutamate receptor subtypes 1 and 5 are activators of extracellular signal-regulated kinase signaling required for inflammatory pain in mice. J. Neurosci. 2001, 21, 3771–3779. [Google Scholar] [CrossRef] [Green Version]
- Adwanikar, H.; Karim, F.; Gereau, R.W.t. Inflammation persistently enhances nocifensive behaviors mediated by spinal group I mGluRs through sustained ERK activation. Pain 2004, 111, 125–135. [Google Scholar] [CrossRef]
- Hu, H.J.; Alter, B.J.; Carrasquillo, Y.; Qiu, C.S.; Gereau, R.W.t. Metabotropic glutamate receptor 5 modulates nociceptive plasticity via extracellular signal-regulated kinase-Kv4.2 signaling in spinal cord dorsal horn neurons. J. Neurosci. 2007, 27, 13181–13191. [Google Scholar] [CrossRef] [Green Version]
- Dolan, S.; Nolan, A.M. Behavioural evidence supporting a differential role for group I and II metabotropic glutamate receptors in spinal nociceptive transmission. Neuropharmacology 2000, 39, 1132–1138. [Google Scholar] [CrossRef]
- Fisher, K.; Coderre, T.J. The contribution of metabotropic glutamate receptors (mGluRs) to formalin-induced nociception. Pain 1996, 68, 255–263. [Google Scholar] [CrossRef]
- Vincent, K.; Cornea, V.M.; Jong, Y.I.; Laferriere, A.; Kumar, N.; Mickeviciute, A.; Fung, J.S.T.; Bandegi, P.; Ribeiro-da-Silva, A.; O’Malley, K.L.; et al. Intracellular mGluR5 plays a critical role in neuropathic pain. Nat. Commun. 2016, 7, 10604. [Google Scholar] [CrossRef] [Green Version]
- Vincent, K.; Wang, S.F.; Laferriere, A.; Kumar, N.; Coderre, T.J. Spinal intracellular metabotropic glutamate receptor 5 (mGluR5) contributes to pain and c-fos expression in a rat model of inflammatory pain. Pain 2017, 158, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Fisher, K.; Fundytus, M.E.; Cahill, C.M.; Coderre, T.J. Intrathecal administration of the mGluR compound, (S)-4CPG, attenuates hyperalgesia and allodynia associated with sciatic nerve constriction injury in rats. Pain 1998, 77, 59–66. [Google Scholar] [CrossRef]
- Fisher, K.; Lefebvre, C.; Coderre, T.J. Antinociceptive effects following intrathecal pretreatment with selective metabotropic glutamate receptor compounds in a rat model of neuropathic pain. Pharmacol. Biochem. Behav. 2002, 73, 411–418. [Google Scholar] [CrossRef]
- Soliman, A.C.; Yu, J.S.; Coderre, T.J. mGlu and NMDA receptor contributions to capsaicin-induced thermal and mechanical hypersensitivity. Neuropharmacology 2005, 48, 325–332. [Google Scholar] [CrossRef]
- Xie, J.D.; Chen, S.R.; Pan, H.L. Presynaptic mGluR5 receptor controls glutamatergic input through protein kinase C-NMDA receptors in paclitaxel-induced neuropathic pain. J. Biol. Chem. 2017, 292, 20644–20654. [Google Scholar] [CrossRef] [Green Version]
- Li, J.Q.; Chen, S.R.; Chen, H.; Cai, Y.Q.; Pan, H.L. Regulation of increased glutamatergic input to spinal dorsal horn neurons by mGluR5 in diabetic neuropathic pain. J. Neurochem. 2010, 112, 162–172. [Google Scholar] [CrossRef] [Green Version]
- Kolber, B.J.; Montana, M.C.; Carrasquillo, Y.; Xu, J.; Heinemann, S.F.; Muglia, L.J.; Gereau, R.W.t. Activation of metabotropic glutamate receptor 5 in the amygdala modulates pain-like behavior. J. Neurosci. 2010, 30, 8203–8213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Ji, G.; Neugebauer, V. Mitochondrial reactive oxygen species are activated by mGluR5 through IP3 and activate ERK and PKA to increase excitability of amygdala neurons and pain behavior. J. Neurosci. 2011, 31, 1114–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maione, S.; Marabese, I.; Leyva, J.; Palazzo, E.; de Novellis, V.; Rossi, F. Characterisation of mGluRs which modulate nociception in the PAG of the mouse. Neuropharmacology 1998, 37, 1475–1483. [Google Scholar] [CrossRef]
- Maione, S.; Oliva, P.; Marabese, I.; Palazzo, E.; Rossi, F.; Berrino, L.; Filippelli, A. Periaqueductal gray matter metabotropic glutamate receptors modulate formalin-induced nociception. Pain 2000, 85, 183–189. [Google Scholar] [CrossRef]
- Han, J.S.; Neugebauer, V. mGluR1 and mGluR5 antagonists in the amygdala inhibit different components of audible and ultrasonic vocalizations in a model of arthritic pain. Pain 2005, 113, 211–222. [Google Scholar] [CrossRef]
- Luongo, L.; de Novellis, V.; Gatta, L.; Palazzo, E.; Vita, D.; Guida, F.; Giordano, C.; Siniscalco, D.; Marabese, I.; De Chiaro, M.; et al. Role of metabotropic glutamate receptor 1 in the basolateral amygdala-driven prefrontal cortical deactivation in inflammatory pain in the rat. Neuropharmacology 2013, 66, 317–329. [Google Scholar] [CrossRef]
- Chung, G.; Kim, C.Y.; Yun, Y.C.; Yoon, S.H.; Kim, M.H.; Kim, Y.K.; Kim, S.J. Upregulation of prefrontal metabotropic glutamate receptor 5 mediates neuropathic pain and negative mood symptoms after spinal nerve injury in rats. Sci. Rep. 2017, 7, 9743. [Google Scholar] [CrossRef]
- Gomez-Santacana, X.; Pittolo, S.; Rovira, X.; Lopez, M.; Zussy, C.; Dalton, J.A.; Faucherre, A.; Jopling, C.; Pin, J.P.; Ciruela, F.; et al. Illuminating Phenylazopyridines To Photoswitch Metabotropic Glutamate Receptors: From the Flask to the Animals. ACS Cent. Sci. 2017, 3, 81–91. [Google Scholar] [CrossRef]
- Kiritoshi, T.; Ji, G.; Neugebauer, V. Rescue of Impaired mGluR5-Driven Endocannabinoid Signaling Restores Prefrontal Cortical Output to Inhibit Pain in Arthritic Rats. J. Neurosci. 2016, 36, 837–850. [Google Scholar] [CrossRef] [Green Version]
- Kung, L.H.; Gong, K.; Adedoyin, M.; Ng, J.; Bhargava, A.; Ohara, P.T.; Jasmin, L. Evidence for glutamate as a neuroglial transmitter within sensory ganglia. PLoS ONE 2013, 8, e68312. [Google Scholar] [CrossRef] [Green Version]
- Gong, K.; Kung, L.H.; Magni, G.; Bhargava, A.; Jasmin, L. Increased response to glutamate in small diameter dorsal root ganglion neurons after sciatic nerve injury. PLoS ONE 2014, 9, e95491. [Google Scholar] [CrossRef] [Green Version]
- Stanfa, L.C.; Dickenson, A.H. Inflammation alters the effects of mGlu receptor agonists on spinal nociceptive neurones. Eur. J. Pharmacol. 1998, 347, 165–172. [Google Scholar] [CrossRef]
- Neugebauer, V.; Chen, P.S.; Willis, W.D. Role of metabotropic glutamate receptor subtype mGluR1 in brief nociception and central sensitization of primate STT cells. J. Neurophysiol. 1999, 82, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Neugebauer, V. Amygdala physiology in pain. Handb. Behav. Neurosci. 2020, 26, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.M.; Neugebauer, V. Cortico-limbic pain mechanisms. Neurosci. Lett. 2019, 702, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Koga, K.; Li, S.; Zhuo, M. Metabotropic Glutamate Receptor Dependent Cortical Plasticity in Chronic Pain. Curr. Neuropharmacol. 2016, 14, 427–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Neugebauer, V. Differential roles of mGluR1 and mGluR5 in brief and prolonged nociceptive processing in central amygdala neurons. J. Neurophysiol. 2004, 91, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Neugebauer, V.; Li, W.; Bird, G.C.; Bhave, G.; Gereau, R.W.t. Synaptic plasticity in the amygdala in a model of arthritic pain: Differential roles of metabotropic glutamate receptors 1 and 5. J. Neurosci. 2003, 23, 52–63. [Google Scholar] [CrossRef] [Green Version]
- Ren, W.; Neugebauer, V. Pain-related increase of excitatory transmission and decrease of inhibitory transmission in the central nucleus of the amygdala are mediated by mGluR1. Mol. Pain 2010, 6, 93. [Google Scholar] [CrossRef] [Green Version]
- Ji, G.; Sun, H.; Fu, Y.; Li, Z.; Pais-Vieira, M.; Galhardo, V.; Neugebauer, V. Cognitive impairment in pain through amygdala-driven prefrontal cortical deactivation. J. Neurosci. 2010, 30, 5451–5464. [Google Scholar] [CrossRef]
- Ji, G.; Neugebauer, V. Pain-related deactivation of medial prefrontal cortical neurons involves mGluR1 and GABA(A) receptors. J. Neurophysiol. 2011, 106, 2642–2652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, G.; Neugebauer, V. CB1 augments mGluR5 function in medial prefrontal cortical neurons to inhibit amygdala hyperactivity in an arthritis pain model. Eur. J. Neurosci. 2014, 39, 455–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boccella, S.; Marabese, I.; Iannotta, M.; Belardo, C.; Neugebauer, V.; Mazzitelli, M.; Pieretti, G.; Maione, S.; Palazzo, E. Metabotropic Glutamate Receptor 5 and 8 Modulate the Ameliorative Effect of Ultramicronized Palmitoylethanolamide on Cognitive Decline Associated with Neuropathic Pain. Int. J. Mol. Sci. 2019, 20, 1757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truini, A.; Piroso, S.; Pasquale, E.; Notartomaso, S.; Di Stefano, G.; Lattanzi, R.; Battaglia, G.; Nicoletti, F.; Cruccu, G. N-acetyl-cysteine, a drug that enhances the endogenous activation of group-II metabotropic glutamate receptors, inhibits nociceptive transmission in humans. Mol. Pain 2015, 11, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernabucci, M.; Notartomaso, S.; Zappulla, C.; Fazio, F.; Cannella, M.; Motolese, M.; Battaglia, G.; Bruno, V.; Gradini, R.; Nicoletti, F. N-Acetyl-cysteine causes analgesia by reinforcing the endogenous activation of type-2 metabotropic glutamate receptors. Mol. Pain 2012, 8, 77. [Google Scholar] [CrossRef]
- Sharpe, E.F.; Kingston, A.E.; Lodge, D.; Monn, J.A.; Headley, P.M. Systemic pre-treatment with a group II mGlu agonist, LY379268, reduces hyperalgesia in vivo. Br. J. Pharmacol. 2002, 135, 1255–1262. [Google Scholar] [CrossRef] [Green Version]
- Mazzitelli, M.; Neugebauer, V. Amygdala group II mGluRs mediate the inhibitory effects of systemic group II mGluR activation on behavior and spinal neurons in a rat model of arthritis pain. Neuropharmacology 2019, 158, 107706. [Google Scholar] [CrossRef]
- Simmons, R.M.; Webster, A.A.; Kalra, A.B.; Iyengar, S. Group II mGluR receptor agonists are effective in persistent and neuropathic pain models in rats. Pharmacol. Biochem. Behav. 2002, 73, 419–427. [Google Scholar] [CrossRef]
- Johnson, M.P.; Muhlhauser, M.A.; Nisenbaum, E.S.; Simmons, R.M.; Forster, B.M.; Knopp, K.L.; Yang, L.; Morrow, D.; Li, D.L.; Kennedy, J.D.; et al. Broad spectrum efficacy with LY2969822, an oral prodrug of metabotropic glutamate 2/3 receptor agonist LY2934747, in rodent pain models. Br. J. Pharmacol. 2017, 174, 822–835. [Google Scholar] [CrossRef] [Green Version]
- Zammataro, M.; Chiechio, S.; Montana, M.C.; Traficante, A.; Copani, A.; Nicoletti, F.; Gereau, R.W.t. mGlu2 metabotropic glutamate receptors restrain inflammatory pain and mediate the analgesic activity of dual mGlu2/mGlu3 receptor agonists. Mol. Pain 2011, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Chiechio, S.; Caricasole, A.; Barletta, E.; Storto, M.; Catania, M.V.; Copani, A.; Vertechy, M.; Nicolai, R.; Calvani, M.; Melchiorri, D.; et al. L-Acetylcarnitine induces analgesia by selectively up-regulating mGlu2 metabotropic glutamate receptors. Mol. Pharmacol. 2002, 61, 989–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ardizzone, A.; Fusco, R.; Casili, G.; Lanza, M.; Impellizzeri, D.; Esposito, E.; Cuzzocrea, S. Effect of Ultra-Micronized-Palmitoylethanolamide and Acetyl-l-Carnitine on Experimental Model of Inflammatory Pain. Int. J. Mol. Sci. 2021, 22, 1967. [Google Scholar] [CrossRef] [PubMed]
- Notartomaso, S.; Scarselli, P.; Mascio, G.; Liberatore, F.; Mazzon, E.; Mammana, S.; Gugliandolo, A.; Cruccu, G.; Bruno, V.; Nicoletti, F.; et al. N-Acetylcysteine causes analgesia in a mouse model of painful diabetic neuropathy. Mol. Pain 2020, 16, 1744806920904292. [Google Scholar] [CrossRef] [PubMed]
- Chiechio, S.; Zammataro, M.; Morales, M.E.; Busceti, C.L.; Drago, F.; Gereau, R.W.t.; Copani, A.; Nicoletti, F. Epigenetic modulation of mGlu2 receptors by histone deacetylase inhibitors in the treatment of inflammatory pain. Mol. Pharmacol. 2009, 75, 1014–1020. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Hirasawa, S.; Wroblewska, B.; Grajkowska, E.; Zhou, J.; Kozikowski, A.; Wroblewski, J.; Neale, J.H. Antinociceptive effects of N-acetylaspartylglutamate (NAAG) peptidase inhibitors ZJ-11, ZJ-17 and ZJ-43 in the rat formalin test and in the rat neuropathic pain model. Eur. J. Neurosci. 2004, 20, 483–494. [Google Scholar] [CrossRef]
- Nonaka, T.; Yamada, T.; Ishimura, T.; Zuo, D.; Moffett, J.R.; Neale, J.H.; Yamamoto, T. A role for the locus coeruleus in the analgesic efficacy of N-acetylaspartylglutamate peptidase (GCPII) inhibitors ZJ43 and 2-PMPA. Mol. Pain 2017, 13, 1744806917697008. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Zhou, S.; Carlton, S.M. Group II metabotropic glutamate receptor activation attenuates peripheral sensitization in inflammatory states. Neuroscience 2008, 154, 754–766. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, T.; Saito, O.; Aoe, T.; Bartolozzi, A.; Sarva, J.; Zhou, J.; Kozikowski, A.; Wroblewska, B.; Bzdega, T.; Neale, J.H. Local administration of N-acetylaspartylglutamate (NAAG) peptidase inhibitors is analgesic in peripheral pain in rats. Eur. J. Neurosci. 2007, 25, 147–158. [Google Scholar] [CrossRef]
- Yang, D.; Gereau, R.W.t. Peripheral group II metabotropic glutamate receptors (mGluR2/3) regulate prostaglandin E2-mediated sensitization of capsaicin responses and thermal nociception. J. Neurosci. 2002, 22, 6388–6393. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Gereau, R.W.t. Peripheral group II metabotropic glutamate receptors mediate endogenous anti-allodynia in inflammation. Pain 2003, 106, 411–417. [Google Scholar] [CrossRef]
- Carlton, S.M.; Du, J.; Zhou, S. Group II metabotropic glutamate receptor activation on peripheral nociceptors modulates TRPV1 function. Brain Res. 2009, 1248, 86–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlton, S.M.; Zhou, S.; Govea, R.; Du, J. Group II/III metabotropic glutamate receptors exert endogenous activity-dependent modulation of TRPV1 receptors on peripheral nociceptors. J. Neurosci. 2011, 31, 12727–12737. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.; Park, E.H.; Cho, H.Y.; Kim, Y.I.; Han, H.C. Peripheral group II and III metabotropic glutamate receptors in the knee joint attenuate carrageenan-induced nociceptive behavior in rats. Neurosci. Lett. 2013, 542, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.Y.; Chen, S.R.; Chen, H.; Pan, H.L. Functional plasticity of group II metabotropic glutamate receptors in regulating spinal excitatory and inhibitory synaptic input in neuropathic pain. J. Pharmacol. Exp. Ther. 2011, 336, 254–264. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Zhang, J.; Shi, J.; Feng, Y.; Sun, Z.S.; Li, H. Antinociceptive synergistic effect of spinal mGluR2/3 antagonist and glial cells inhibitor on peripheral inflammation-induced mechanical hypersensitivity. Brain Res. Bull. 2009, 79, 219–223. [Google Scholar] [CrossRef]
- Cao, D.Y.; Bai, G.; Ji, Y.; Traub, R.J. Epigenetic upregulation of metabotropic glutamate receptor 2 in the spinal cord attenuates oestrogen-induced visceral hypersensitivity. Gut 2015, 64, 1913–1920. [Google Scholar] [CrossRef]
- Neto, F.L.; Castro-Lopes, J.M. Antinociceptive effect of a group II metabotropic glutamate receptor antagonist in the thalamus of monoarthritic rats. Neurosci. Lett. 2000, 296, 25–28. [Google Scholar] [CrossRef]
- Yamamoto, T.; Kozikowski, A.; Zhou, J.; Neale, J.H. Intracerebroventricular administration of N-acetylaspartylglutamate (NAAG) peptidase inhibitors is analgesic in inflammatory pain. Mol. Pain 2008, 4, 31. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Zuo, D.; Yamamoto, T.; Olszewski, R.T.; Bzdega, T.; Moffett, J.R.; Neale, J.H. NAAG peptidase inhibition in the periaqueductal gray and rostral ventromedial medulla reduces flinching in the formalin model of inflammation. Mol. Pain 2012, 8, 67. [Google Scholar] [CrossRef]
- Tanabe, Y.; Masu, M.; Ishii, T.; Shigemoto, R.; Nakanishi, S. A family of metabotropic glutamate receptors. Neuron 1992, 8, 169–179. [Google Scholar] [CrossRef]
- Gu, G.; Lorrain, D.S.; Wei, H.; Cole, R.L.; Zhang, X.; Daggett, L.P.; Schaffhauser, H.J.; Bristow, L.J.; Lechner, S.M. Distribution of metabotropic glutamate 2 and 3 receptors in the rat forebrain: Implication in emotional responses and central disinhibition. Brain Res. 2008, 1197, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.A.; Johnson, B.G.; Zhang, C.; Salhoff, C.; Kingston, A.E.; Calligaro, D.O.; Monn, J.A.; Schoepp, D.D.; Marek, G.J. CNS distribution of metabotropic glutamate 2 and 3 receptors: Transgenic mice and [(3)H]LY459477 autoradiography. Neuropharmacology 2013, 66, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Mazzitelli, M.; Palazzo, E.; Maione, S.; Neugebauer, V. Group II Metabotropic Glutamate Receptors: Role in Pain Mechanisms and Pain Modulation. Front. Mol. Neurosci. 2018, 11, 383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montana, M.C.; Gereau, R.W. Metabotropic glutamate receptors as targets for analgesia: Antagonism, activation, and allosteric modulation. Curr. Pharm. Biotechnol. 2011, 12, 1681–1688. [Google Scholar] [CrossRef]
- Kalivas, P.W. The glutamate homeostasis hypothesis of addiction. Nat. Rev. Neurosci. 2009, 10, 561–572. [Google Scholar] [CrossRef]
- Chiechio, S.; Nicoletti, F. Metabotropic glutamate receptors and the control of chronic pain. Curr. Opin. Pharmacol. 2012, 12, 28–34. [Google Scholar] [CrossRef]
- Neale, J.H.; Olszewski, R.T.; Zuo, D.; Janczura, K.J.; Profaci, C.P.; Lavin, K.M.; Madore, J.C.; Bzdega, T. Advances in understanding the peptide neurotransmitter NAAG and appearance of a new member of the NAAG neuropeptide family. J. Neurochem. 2011, 118, 490–498. [Google Scholar] [CrossRef] [Green Version]
- Neale, J.H. N-acetylaspartylglutamate is an agonist at mGluR(3) in vivo and in vitro. J. Neurochem. 2011, 119, 891–895. [Google Scholar] [CrossRef] [Green Version]
- Neale, J.H.; Olszewski, R.T.; Gehl, L.M.; Wroblewska, B.; Bzdega, T. The neurotransmitter N-acetylaspartylglutamate in models of pain, ALS, diabetic neuropathy, CNS injury and schizophrenia. Trends Pharmacol. Sci. 2005, 26, 477–484. [Google Scholar] [CrossRef]
- Sheahan, T.D.; Valtcheva, M.V.; McIlvried, L.A.; Pullen, M.Y.; Baranger, D.A.A.; Gereau, R.W.t. Metabotropic Glutamate Receptor 2/3 (mGluR2/3) Activation Suppresses TRPV1 Sensitization in Mouse, But Not Human, Sensory Neurons. eNeuro 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Davidson, S.; Golden, J.P.; Copits, B.A.; Ray, P.R.; Vogt, S.K.; Valtcheva, M.V.; Schmidt, R.E.; Ghetti, A.; Price, T.J.; Gereau, R.W.t. Group II mGluRs suppress hyperexcitability in mouse and human nociceptors. Pain 2016, 157, 2081–2088. [Google Scholar] [CrossRef] [PubMed]
- Gerber, G.; Zhong, J.; Youn, D.; Randic, M. Group II and group III metabotropic glutamate receptor agonists depress synaptic transmission in the rat spinal cord dorsal horn. Neuroscience 2000, 100, 393–406. [Google Scholar] [CrossRef]
- Neugebauer, V.; Chen, P.S.; Willis, W.D. Groups II and III metabotropic glutamate receptors differentially modulate brief and prolonged nociception in primate STT cells. J. Neurophysiol. 2000, 84, 2998–3009. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Neugebauer, V. Differential changes of group II and group III mGluR function in central amygdala neurons in a model of arthritic pain. J. Neurophysiol. 2006, 96, 1803–1815. [Google Scholar] [CrossRef]
- Han, J.S.; Fu, Y.; Bird, G.C.; Neugebauer, V. Enhanced group II mGluR-mediated inhibition of pain-related synaptic plasticity in the amygdala. Mol. Pain 2006, 2, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adedoyin, M.O.; Vicini, S.; Neale, J.H. Endogenous N-acetylaspartylglutamate (NAAG) inhibits synaptic plasticity/transmission in the amygdala in a mouse inflammatory pain model. Mol. Pain 2010, 6, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiritoshi, T.; Neugebauer, V. Group II mGluRs modulate baseline and arthritis pain-related synaptic transmission in the rat medial prefrontal cortex. Neuropharmacology 2015, 95, 388–394. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Kadakia, F.; Davidson, S. Group II metabotropic glutamate receptor expressing neurons in anterior cingulate cortex become sensitized after inflammatory and neuropathic pain. Mol. Pain 2020, 16, 1744806920915339. [Google Scholar] [CrossRef]
- Conn, P.J.; Pin, J.P. Pharmacology and functions of metabotropic glutamate receptors. Annu. Rev. Pharmacol. Toxicol. 1997, 37, 205–237. [Google Scholar] [CrossRef] [Green Version]
- Schoepp, D.D.; Jane, D.E.; Monn, J.A. Pharmacological agents acting at subtypes of metabotropic glutamate receptors. Neuropharmacology 1999, 38, 1431–1476. [Google Scholar] [CrossRef]
- Shigemoto, R.; Kinoshita, A.; Wada, E.; Nomura, S.; Ohishi, H.; Takada, M.; Flor, P.J.; Neki, A.; Abe, T.; Nakanishi, S.; et al. Differential presynaptic localization of metabotropic glutamate receptor subtypes in the rat hippocampus. J. Neurosci. 1997, 17, 7503–7522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cartmell, J.; Schoepp, D.D. Regulation of neurotransmitter release by metabotropic glutamate receptors. J. Neurochem. 2000, 75, 889–907. [Google Scholar] [CrossRef] [PubMed]
- Lavreysen, H.; Dautzenberg, F.M. Therapeutic potential of group III metabotropic glutamate receptors. Curr. Med. Chem. 2008, 15, 671–684. [Google Scholar] [CrossRef] [PubMed]
- Carlton, S.M.; Hargett, G.L. Colocalization of metabotropic glutamate receptors in rat dorsal root ganglion cells. J. Comp. Neurol. 2007, 501, 780–789. [Google Scholar] [CrossRef] [PubMed]
- Vilar, B.; Busserolles, J.; Ling, B.; Laffray, S.; Ulmann, L.; Malhaire, F.; Chapuy, E.; Aissouni, Y.; Etienne, M.; Bourinet, E.; et al. Alleviating pain hypersensitivity through activation of type 4 metabotropic glutamate receptor. J. Neurosci. 2013, 33, 18951–18965. [Google Scholar] [CrossRef] [Green Version]
- Lourenco Neto, F.; Schadrack, J.; Berthele, A.; Zieglgansberger, W.; Tolle, T.R.; Castro-Lopes, J.M. Differential distribution of metabotropic glutamate receptor subtype mRNAs in the thalamus of the rat. Brain Res. 2000, 854, 93–105. [Google Scholar] [CrossRef]
- Ohishi, H.; Akazawa, C.; Shigemoto, R.; Nakanishi, S.; Mizuno, N. Distributions of the mRNAs for L-2-amino-4-phosphonobutyrate-sensitive metabotropic glutamate receptors, mGluR4 and mGluR7, in the rat brain. J. Comp. Neurol. 1995, 360, 555–570. [Google Scholar] [CrossRef]
- Nakajima, Y.; Iwakabe, H.; Akazawa, C.; Nawa, H.; Shigemoto, R.; Mizuno, N.; Nakanishi, S. Molecular characterization of a novel retinal metabotropic glutamate receptor mGluR6 with a high agonist selectivity for L-2-amino-4-phosphonobutyrate. J. Biol. Chem. 1993, 268, 11868–11873. [Google Scholar] [CrossRef]
- Palazzo, E.; Marabese, I.; de Novellis, V.; Rossi, F.; Maione, S. Metabotropic Glutamate Receptor 7: From Synaptic Function to Therapeutic Implications. Curr. Neuropharmacol. 2016, 14, 504–513. [Google Scholar] [CrossRef] [Green Version]
- Corti, C.; Restituito, S.; Rimland, J.M.; Brabet, I.; Corsi, M.; Pin, J.P.; Ferraguti, F. Cloning and characterization of alternative mRNA forms for the rat metabotropic glutamate receptors mGluR7 and mGluR8. Eur. J. Neurosci. 1998, 10, 3629–3641. [Google Scholar] [CrossRef]
- Pamidimukkala, J.; Hoang, C.J.; Hay, M. Expression of metabotropic glutamate receptor 8 in autonomic cell groups of the medulla oblongata of the rat. Brain Res. 2002, 957, 162–173. [Google Scholar] [CrossRef]
- Tong, Q.; Kirchgessner, A.L. Localization and function of metabotropic glutamate receptor 8 in the enteric nervous system. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, G992–G1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- G, S.; Suvarna, P.; Hadigal, S.; Kamath, P.; Prabhu, N.; Shenoy, K.A.; Lc, P. Can Metabotropic Glutamate Receptor 7 (mGluR 7) be a Novel Target for Analgesia? J. Clin. Diagn. Res. 2014, 8, HC16–HC18. [Google Scholar] [CrossRef] [PubMed]
- Dolan, S.; Gunn, M.D.; Biddlestone, L.; Nolan, A.M. The selective metabotropic glutamate receptor 7 allosteric agonist AMN082 inhibits inflammatory pain-induced and incision-induced hypersensitivity in rat. Behav. Pharmacol. 2009, 20, 596–604. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, E.; Romano, R.; Luongo, L.; Boccella, S.; De Gregorio, D.; Giordano, M.E.; Rossi, F.; Marabese, I.; Scafuro, M.A.; de Novellis, V.; et al. MMPIP, an mGluR7-selective negative allosteric modulator, alleviates pain and normalizes affective and cognitive behavior in neuropathic mice. Pain 2015, 156, 1060–1073. [Google Scholar] [CrossRef]
- Moloney, R.D.; Golubeva, A.V.; O’Connor, R.M.; Kalinichev, M.; Dinan, T.G.; Cryan, J.F. Negative allosteric modulation of the mGlu7 receptor reduces visceral hypersensitivity in a stress-sensitive rat strain. Neurobiol. Stress 2015, 2, 28–33. [Google Scholar] [CrossRef] [Green Version]
- Marabese, I.; de Novellis, V.; Palazzo, E.; Scafuro, M.A.; Vita, D.; Rossi, F.; Maione, S. Effects of (S)-3,4-DCPG, an mGlu8 receptor agonist, on inflammatory and neuropathic pain in mice. Neuropharmacology 2007, 52, 253–262. [Google Scholar] [CrossRef]
- Li, Y.L.; Chang, X.R.; Ma, J.T.; Zhao, X.; Yin, L.T.; Yan, L.J.; Guo, J.H.; Zhang, C.; Yang, X.R. Activation of peripheral group III metabotropic glutamate receptors suppressed formalin-induced nociception. Clin. Exp. Pharmacol. Physiol. 2022, 49, 319–326. [Google Scholar] [CrossRef]
- Goudet, C.; Chapuy, E.; Alloui, A.; Acher, F.; Pin, J.P.; Eschalier, A. Group III metabotropic glutamate receptors inhibit hyperalgesia in animal models of inflammation and neuropathic pain. Pain 2008, 137, 112–124. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.R.; Pan, H.L. Distinct roles of group III metabotropic glutamate receptors in control of nociception and dorsal horn neurons in normal and nerve-injured Rats. J. Pharmacol. Exp. Ther. 2005, 312, 120–126. [Google Scholar] [CrossRef] [Green Version]
- Rovira, X.; Trapero, A.; Pittolo, S.; Zussy, C.; Faucherre, A.; Jopling, C.; Giraldo, J.; Pin, J.P.; Gorostiza, P.; Goudet, C.; et al. OptoGluNAM4.1, a Photoswitchable Allosteric Antagonist for Real-Time Control of mGlu4 Receptor Activity. Cell Chem. Biol. 2016, 23, 929–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Jiang, C.; Ba, X.; Yang, S.; Wu, J.; Huang, Z.; Jin, G.; Hao, Y. Selective activation of metabotropic glutamate receptor 7 blocks paclitaxel-induced acute neuropathic pain and suppresses spinal glial reactivity in rats. Psychopharmacology 2021, 238, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, E.; Marabese, I.; Soukupova, M.; Luongo, L.; Boccella, S.; Giordano, C.; de Novellis, V.; Rossi, F.; Maione, S. Metabotropic glutamate receptor subtype 8 in the amygdala modulates thermal threshold, neurotransmitter release, and rostral ventromedial medulla cell activity in inflammatory pain. J. Neurosci. 2011, 31, 4687–4697. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, E.; Fu, Y.; Ji, G.; Maione, S.; Neugebauer, V. Group III mGluR7 and mGluR8 in the amygdala differentially modulate nocifensive and affective pain behaviors. Neuropharmacology 2008, 55, 537–545. [Google Scholar] [CrossRef] [Green Version]
- Marabese, I.; Rossi, F.; Palazzo, E.; de Novellis, V.; Starowicz, K.; Cristino, L.; Vita, D.; Gatta, L.; Guida, F.; Di Marzo, V.; et al. Periaqueductal gray metabotropic glutamate receptor subtype 7 and 8 mediate opposite effects on amino acid release, rostral ventromedial medulla cell activities, and thermal nociception. J. Neurophysiol. 2007, 98, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Marabese, I.; Boccella, S.; Iannotta, M.; Luongo, L.; de Novellis, V.; Guida, F.; Serra, N.; Farina, A.; Maione, S.; Palazzo, E. Metabotropic glutamate receptor subtype 7 in the dorsal striatum oppositely modulates pain in sham and neuropathic rats. Neuropharmacology 2018, 135, 86–99. [Google Scholar] [CrossRef]
- Liu, X.H.; Han, M.; Zhu, J.X.; Sun, N.; Tang, J.S.; Huo, F.Q.; Li, J.; Xu, F.Y.; Du, J.Q. Metabotropic glutamate subtype 7 and 8 receptors oppositely modulate cardiac nociception in the rat nucleus tractus solitarius. Neuroscience 2012, 220, 322–329. [Google Scholar] [CrossRef]
- Kahl, E.; Fendt, M. Metabotropic Glutamate Receptors 7 within the Nucleus Accumbens are Involved in Relief Learning in Rats. Curr. Neuropharmacol. 2016, 14, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Rossi, F.; Marabese, I.; De Chiaro, M.; Boccella, S.; Luongo, L.; Guida, F.; De Gregorio, D.; Giordano, C.; de Novellis, V.; Palazzo, E.; et al. Dorsal striatum metabotropic glutamate receptor 8 affects nocifensive responses and rostral ventromedial medulla cell activity in neuropathic pain conditions. J. Neurophysiol. 2014, 111, 2196–2209. [Google Scholar] [CrossRef] [Green Version]
- Palazzo, E.; Boccella, S.; Marabese, I.; Perrone, M.; Belardo, C.; Iannotta, M.; Scuteri, D.; De Dominicis, E.; Pagano, M.; Infantino, R.; et al. Homo-AMPA in the periaqueductal grey modulates pain and rostral ventromedial medulla activity in diabetic neuropathic mice. Neuropharmacology 2022, 212, 109047. [Google Scholar] [CrossRef]
- Hosseini, M.; Parviz, M.; Shabanzadeh, A.P.; Zamani, E. Evaluation of the Effect of (S)-3,4-Dicarboxyphenylglycine as a Metabotropic Glutamate Receptors Subtype 8 Agonist on Thermal Nociception Following Central Neuropathic Pain. Asian Spine J. 2021, 15, 200–206. [Google Scholar] [CrossRef]
- Hosseini, M.; Parviz, M.; Shabanzadeh, A.P.; Zamani, E.; Mohseni-Moghaddam, P.; Gholami, L.; Mehrabadi, S. The inhibiting role of periaqueductal gray metabotropic glutamate receptor subtype 8 in a rat model of central neuropathic pain. Neurol. Res. 2020, 42, 515–521. [Google Scholar] [CrossRef]
- Park, E.H.; Lee, S.W.; Moon, S.W.; Suh, H.R.; Kim, Y.I.; Han, H.C. Activation of peripheral group III metabotropic glutamate receptors inhibits pain transmission by decreasing neuronal excitability in the CFA-inflamed knee joint. Neurosci. Lett. 2019, 694, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.M.; Chen, S.R.; Pan, H.L. Effects of activation of group III metabotropic glutamate receptors on spinal synaptic transmission in a rat model of neuropathic pain. Neuroscience 2009, 158, 875–884. [Google Scholar] [CrossRef] [Green Version]
- Han, J.S.; Bird, G.C.; Neugebauer, V. Enhanced group III mGluR-mediated inhibition of pain-related synaptic plasticity in the amygdala. Neuropharmacology 2004, 46, 918–926. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Palazzo, E.; Maione, S.; Neugebauer, V. Differential effects of mGluR7 and mGluR8 activation on pain-related synaptic activity in the amygdala. Neuropharmacology 2011, 61, 1334–1344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marabese, I.; de Novellis, V.; Palazzo, E.; Mariani, L.; Siniscalco, D.; Rodella, L.; Rossi, F.; Maione, S. Differential roles of mGlu8 receptors in the regulation of glutamate and gamma-aminobutyric acid release at periaqueductal grey level. Neuropharmacology 2005, 49 Suppl. 1, 157–166. [Google Scholar] [CrossRef]
- Chen, G.; Zhang, Y.Q.; Qadri, Y.J.; Serhan, C.N.; Ji, R.R. Microglia in Pain: Detrimental and Protective Roles in Pathogenesis and Resolution of Pain. Neuron 2018, 100, 1292–1311. [Google Scholar] [CrossRef] [Green Version]
- Grace, P.M.; Tawfik, V.L.; Svensson, C.I.; Burton, M.D.; Loggia, M.L.; Hutchinson, M.R. The Neuroimmunology of Chronic Pain: From Rodents to Humans. J. Neurosci. 2021, 41, 855–865. [Google Scholar] [CrossRef]
- Ji, R.R.; Berta, T.; Nedergaard, M. Glia and pain: Is chronic pain a gliopathy? Pain 2013, 154 Suppl. 1, S10–S28. [Google Scholar] [CrossRef]
- Murray, I.; Bhanot, G.; Bhargava, A. Neuron-Glia-Immune Triad and Cortico-Limbic System in Pathology of Pain. Cells 2021, 10, 1553. [Google Scholar] [CrossRef] [PubMed]
- Tozaki-Saitoh, H.; Tsuda, M. Microglia-neuron interactions in the models of neuropathic pain. Biochem. Pharmacol. 2019, 169, 113614. [Google Scholar] [CrossRef]
- Ji, R.R.; Donnelly, C.R.; Nedergaard, M. Astrocytes in chronic pain and itch. Nat. Rev. Neurosci. 2019, 20, 667–685. [Google Scholar] [CrossRef] [PubMed]
- Spampinato, S.F.; Copani, A.; Nicoletti, F.; Sortino, M.A.; Caraci, F. Metabotropic Glutamate Receptors in Glial Cells: A New Potential Target for Neuroprotection? Front. Mol. Neurosci. 2018, 11, 414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.; Pow, D.V. Astrocytes: Glutamate transport and alternate splicing of transporters. Int. J. Biochem. Cell Biol. 2010, 42, 1901–1906. [Google Scholar] [CrossRef]
- Allen, N.J.; Eroglu, C. Cell Biology of Astrocyte-Synapse Interactions. Neuron 2017, 96, 697–708. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Barres, B.A. Reactive Astrocytes: Production, Function, and Therapeutic Potential. Immunity 2017, 46, 957–967. [Google Scholar] [CrossRef] [Green Version]
- Han, R.T.; Kim, R.D.; Molofsky, A.V.; Liddelow, S.A. Astrocyte-immune cell interactions in physiology and pathology. Immunity 2021, 54, 211–224. [Google Scholar] [CrossRef]
- Tay, T.L.; Savage, J.C.; Hui, C.W.; Bisht, K.; Tremblay, M.E. Microglia across the lifespan: From origin to function in brain development, plasticity and cognition. J. Physiol. 2017, 595, 1929–1945. [Google Scholar] [CrossRef] [Green Version]
- Cornell, J.; Salinas, S.; Huang, H.Y.; Zhou, M. Microglia regulation of synaptic plasticity and learning and memory. Neural. Regen. Res. 2022, 17, 705–716. [Google Scholar] [CrossRef]
- Colonna, M.; Butovsky, O. Microglia Function in the Central Nervous System During Health and Neurodegeneration. Annu. Rev. Immunol. 2017, 35, 441–468. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.A.; Boddeke, H.W.; Kettenmann, H. Microglia in Physiology and Disease. Annu. Rev. Physiol. 2017, 79, 619–643. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.R.; Nackley, A.; Huh, Y.; Terrando, N.; Maixner, W. Neuroinflammation and Central Sensitization in Chronic and Widespread Pain. Anesthesiology 2018, 129, 343–366. [Google Scholar] [CrossRef]
- Vergne-Salle, P.; Bertin, P. Chronic pain and neuroinflammation. Joint Bone Spine 2021, 88, 105222. [Google Scholar] [CrossRef] [PubMed]
- Ohishi, H.; Ogawa-Meguro, R.; Shigemoto, R.; Kaneko, T.; Nakanishi, S.; Mizuno, N. Immunohistochemical localization of metabotropic glutamate receptors, mGluR2 and mGluR3, in rat cerebellar cortex. Neuron 1994, 13, 55–66. [Google Scholar] [CrossRef]
- Ohishi, H.; Shigemoto, R.; Nakanishi, S.; Mizuno, N. Distribution of the messenger RNA for a metabotropic glutamate receptor, mGluR2, in the central nervous system of the rat. Neuroscience 1993, 53, 1009–1018. [Google Scholar] [CrossRef]
- Petralia, R.S.; Wang, Y.X.; Niedzielski, A.S.; Wenthold, R.J. The metabotropic glutamate receptors, mGluR2 and mGluR3, show unique postsynaptic, presynaptic and glial localizations. Neuroscience 1996, 71, 949–976. [Google Scholar] [CrossRef]
- Schools, G.P.; Kimelberg, H.K. mGluR3 and mGluR5 are the predominant metabotropic glutamate receptor mRNAs expressed in hippocampal astrocytes acutely isolated from young rats. J. Neurosci. Res. 1999, 58, 533–543. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, K.; Sloan, S.A.; Bennett, M.L.; Scholze, A.R.; O’Keeffe, S.; Phatnani, H.P.; Guarnieri, P.; Caneda, C.; Ruderisch, N.; et al. An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J. Neurosci. 2014, 34, 11929–11947. [Google Scholar] [CrossRef]
- Aronica, E.; van Vliet, E.A.; Mayboroda, O.A.; Troost, D.; da Silva, F.H.; Gorter, J.A. Upregulation of metabotropic glutamate receptor subtype mGluR3 and mGluR5 in reactive astrocytes in a rat model of mesial temporal lobe epilepsy. Eur. J. Neurosci. 2000, 12, 2333–2344. [Google Scholar] [CrossRef]
- Ferraguti, F.; Corti, C.; Valerio, E.; Mion, S.; Xuereb, J. Activated astrocytes in areas of kainate-induced neuronal injury upregulate the expression of the metabotropic glutamate receptors 2/3 and 5. Exp. Brain Res. 2001, 137, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Dolan, S.; Kelly, J.G.; Monteiro, A.M.; Nolan, A.M. Up-regulation of metabotropic glutamate receptor subtypes 3 and 5 in spinal cord in a clinical model of persistent inflammation and hyperalgesia. Pain 2003, 106, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Kellner, V.; Kersbergen, C.J.; Li, S.; Babola, T.A.; Saher, G.; Bergles, D.E. Dual metabotropic glutamate receptor signaling enables coordination of astrocyte and neuron activity in developing sensory domains. Neuron 2021, 109, 2545–2555.e2547. [Google Scholar] [CrossRef] [PubMed]
- Berger, J.V.; Dumont, A.O.; Focant, M.C.; Vergouts, M.; Sternotte, A.; Calas, A.G.; Goursaud, S.; Hermans, E. Opposite regulation of metabotropic glutamate receptor 3 and metabotropic glutamate receptor 5 by inflammatory stimuli in cultured microglia and astrocytes. Neuroscience 2012, 205, 29–38. [Google Scholar] [CrossRef]
- Vermeiren, C.; Hemptinne, I.; Vanhoutte, N.; Tilleux, S.; Maloteaux, J.M.; Hermans, E. Loss of metabotropic glutamate receptor-mediated regulation of glutamate transport in chemically activated astrocytes in a rat model of amyotrophic lateral sclerosis. J. Neurochem. 2006, 96, 719–731. [Google Scholar] [CrossRef]
- Bonifacino, T.; Cattaneo, L.; Gallia, E.; Puliti, A.; Melone, M.; Provenzano, F.; Bossi, S.; Musante, I.; Usai, C.; Conti, F.; et al. In-vivo effects of knocking-down metabotropic glutamate receptor 5 in the SOD1(G93A) mouse model of amyotrophic lateral sclerosis. Neuropharmacology 2017, 123, 433–445. [Google Scholar] [CrossRef]
- Shrivastava, A.N.; Kowalewski, J.M.; Renner, M.; Bousset, L.; Koulakoff, A.; Melki, R.; Giaume, C.; Triller, A. beta-amyloid and ATP-induced diffusional trapping of astrocyte and neuronal metabotropic glutamate type-5 receptors. Glia 2013, 61, 1673–1686. [Google Scholar] [CrossRef]
- Kumar, A.; Dhull, D.K.; Mishra, P.S. Therapeutic potential of mGluR5 targeting in Alzheimer’s disease. Front. Neurosci. 2015, 9, 215. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.; Otgontenger, U.; Jamsranjav, A.; Kim, S.S. Deleterious Alteration of Glia in the Brain of Alzheimer’s Disease. Int. J. Mol. Sci. 2020, 21, 6676. [Google Scholar] [CrossRef]
- Randall, P.A.; Vetreno, R.P.; Makhijani, V.H.; Crews, F.T.; Besheer, J. The Toll-Like Receptor 3 Agonist Poly(I:C) Induces Rapid and Lasting Changes in Gene Expression Related to Glutamatergic Function and Increases Ethanol Self-Administration in Rats. Alcohol. Clin. Exp. Res. 2019, 43, 48–60. [Google Scholar] [CrossRef]
- Osikowicz, M.; Skup, M.; Mika, J.; Makuch, W.; Czarkowska-Bauch, J.; Przewlocka, B. Glial inhibitors influence the mRNA and protein levels of mGlu2/3, 5 and 7 receptors and potentiate the analgesic effects of their ligands in a mouse model of neuropathic pain. Pain 2009, 147, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Cahoy, J.D.; Emery, B.; Kaushal, A.; Foo, L.C.; Zamanian, J.L.; Christopherson, K.S.; Xing, Y.; Lubischer, J.L.; Krieg, P.A.; Krupenko, S.A.; et al. A transcriptome database for astrocytes, neurons, and oligodendrocytes: A new resource for understanding brain development and function. J. Neurosci. 2008, 28, 264–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusnakova, V.; Honsa, P.; Dzamba, D.; Stahlberg, A.; Kubista, M.; Anderova, M. Heterogeneity of astrocytes: From development to injury - single cell gene expression. PLoS ONE 2013, 8, e69734. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; McConnell, E.; Pare, J.F.; Xu, Q.; Chen, M.; Peng, W.; Lovatt, D.; Han, X.; Smith, Y.; Nedergaard, M. Glutamate-dependent neuroglial calcium signaling differs between young and adult brain. Science 2013, 339, 197–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danjo, Y.; Shigetomi, E.; Hirayama, Y.J.; Kobayashi, K.; Ishikawa, T.; Fukazawa, Y.; Shibata, K.; Takanashi, K.; Parajuli, B.; Shinozaki, Y.; et al. Transient astrocytic mGluR5 expression drives synaptic plasticity and subsequent chronic pain in mice. J. Exp. Med. 2022, 219. [Google Scholar] [CrossRef]
- Presto, P.; Mazzitelli, M.; Junell, R.; Griffin, Z.; Neugebauer, V. Sex differences in pain along the neuraxis. Neuropharmacology 2022, 210, 109030. [Google Scholar] [CrossRef]
- Boulware, M.I.; Mermelstein, P.G. Membrane estrogen receptors activate metabotropic glutamate receptors to influence nervous system physiology. Steroids 2009, 74, 608–613. [Google Scholar] [CrossRef] [Green Version]
- Meitzen, J.; Mermelstein, P.G. Estrogen receptors stimulate brain region specific metabotropic glutamate receptors to rapidly initiate signal transduction pathways. J. Chem. Neuroanat. 2011, 42, 236–241. [Google Scholar] [CrossRef] [Green Version]
- Gross, K.S.; Mermelstein, P.G. Estrogen receptor signaling through metabotropic glutamate receptors. Vitam. Horm. 2020, 114, 211–232. [Google Scholar] [CrossRef]
- Tonn Eisinger, K.R.; Larson, E.B.; Boulware, M.I.; Thomas, M.J.; Mermelstein, P.G. Membrane estrogen receptor signaling impacts the reward circuitry of the female brain to influence motivated behaviors. Steroids 2018, 133, 53–59. [Google Scholar] [CrossRef]
- Tonn Eisinger, K.R.; Gross, K.S.; Head, B.P.; Mermelstein, P.G. Interactions between estrogen receptors and metabotropic glutamate receptors and their impact on drug addiction in females. Horm. Behav. 2018, 104, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Tabatadze, N.; Huang, G.; May, R.M.; Jain, A.; Woolley, C.S. Sex Differences in Molecular Signaling at Inhibitory Synapses in the Hippocampus. J. Neurosci. 2015, 35, 11252–11265. [Google Scholar] [CrossRef] [PubMed]
- Gintzler, A.R.; Liu, N.J. Arbiters of endogenous opioid analgesia: Role of CNS estrogenic and glutamatergic systems. Transl. Res. 2021, 234, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.J.; Murugaiyan, V.; Storman, E.M.; Schnell, S.A.; Kumar, A.; Wessendorf, M.W.; Gintzler, A.R. Plasticity of Signaling by Spinal Estrogen Receptor alpha, kappa-Opioid Receptor, and Metabotropic Glutamate Receptors over the Rat Reproductive Cycle Regulates Spinal Endomorphin 2 Antinociception: Relevance of Endogenous-Biased Agonism. J. Neurosci. 2017, 37, 11181–11191. [Google Scholar] [CrossRef]
- Wang, Y.; Ma, Y.; Hu, J.; Cheng, W.; Jiang, H.; Zhang, X.; Li, M.; Ren, J.; Li, X. Prenatal chronic mild stress induces depression-like behavior and sex-specific changes in regional glutamate receptor expression patterns in adult rats. Neuroscience 2015, 301, 363–374. [Google Scholar] [CrossRef]
- Zuena, A.R.; Mairesse, J.; Casolini, P.; Cinque, C.; Alema, G.S.; Morley-Fletcher, S.; Chiodi, V.; Spagnoli, L.G.; Gradini, R.; Catalani, A.; et al. Prenatal restraint stress generates two distinct behavioral and neurochemical profiles in male and female rats. PLoS ONE 2008, 3, e2170. [Google Scholar] [CrossRef] [Green Version]
- Duvoisin, R.M.; Villasana, L.; Pfankuch, T.; Raber, J. Sex-dependent cognitive phenotype of mice lacking mGluR8. Behav. Brain Res. 2010, 209, 21–26. [Google Scholar] [CrossRef] [Green Version]
- De Jesus-Burgos, M.I.; Gonzalez-Garcia, S.; Cruz-Santa, Y.; Perez-Acevedo, N.L. Amygdalar activation of group I metabotropic glutamate receptors produces anti- and pro-conflict effects depending upon animal sex in a sexually dimorphic conditioned conflict-based anxiety model. Behav. Brain Res. 2016, 302, 200–212. [Google Scholar] [CrossRef]
- Veinante, P.; Yalcin, I.; Barrot, M. The amygdala between sensation and affect: A role in pain. J. Mol. Psychiatry 2013, 1, 9. [Google Scholar] [CrossRef] [Green Version]
- Vachon-Presseau, E.; Centeno, M.V.; Ren, W.; Berger, S.E.; Tetreault, P.; Ghantous, M.; Baria, A.; Farmer, M.; Baliki, M.N.; Schnitzer, T.J.; et al. The Emotional Brain as a Predictor and Amplifier of Chronic Pain. J. Dent. Res. 2016, 95, 605–612. [Google Scholar] [CrossRef] [Green Version]
- Elman, I.; Borsook, D. Threat Response System: Parallel Brain Processes in Pain vis-a-vis Fear and Anxiety. Front. Psychiatry 2018, 9, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zammataro, M.; Merlo, S.; Barresi, M.; Parenti, C.; Hu, H.; Sortino, M.A.; Chiechio, S. Chronic Treatment with Fluoxetine Induces Sex-Dependent Analgesic Effects and Modulates HDAC2 and mGlu2 Expression in Female Mice. Front. Pharmacol. 2017, 8, 743. [Google Scholar] [CrossRef] [PubMed] [Green Version]


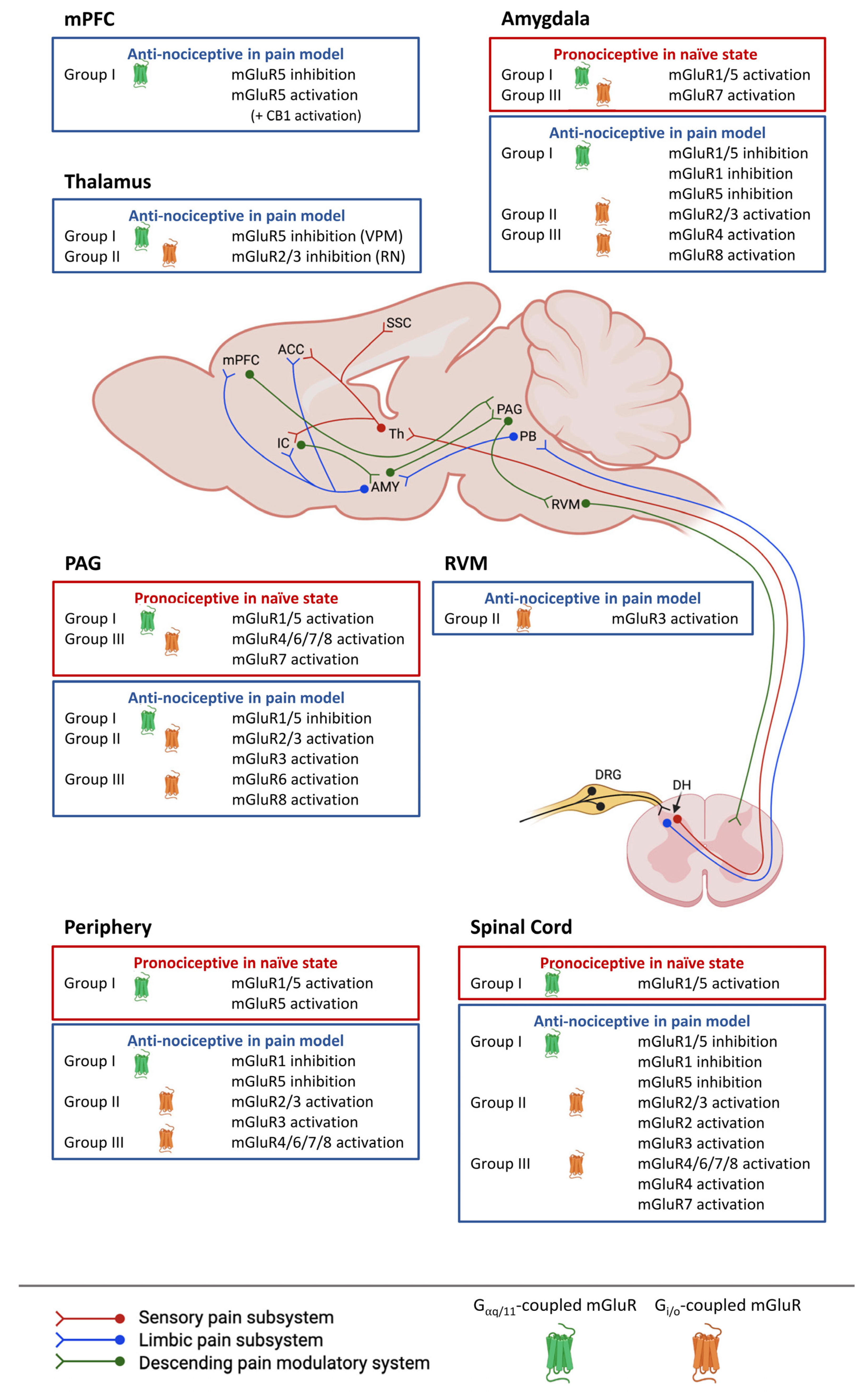{kind=link}
| Receptor | Drug Type | Compound | Route of Administration | Species | Pain Model | Effect | References |
|---|---|---|---|---|---|---|---|
| Systemic | |||||||
| mGluR1 | NAM | EMQMCM | i.p. | Rat | Normal | No effect | [37] |
| Formalin | Inhibition | ||||||
| FTIDC | i.p. | Mouse | Formalin | Inhibition | [38] | ||
| A-841720, A-794282, A-794278, A-850002 | i.p. | Rat | Skin incision, MIA, CFA, CCI, SNL | Inhibition | [39,41] | ||
| LY456236 | i.p. | Mouse | Formalin | Inhibition | [40] | ||
| Rat | SNL | ||||||
| mGluR5 | NAM | MPEP, MTEP | i.p., oral | Rat | Normal | No effect | [37,42] |
| i.v. | Inhibition | [46] | |||||
| Fenobam, MPEP, MTEP | i.p. | Mouse | Formalin, CFA | Inhibition | [26,27,38] | ||
| Oral, i.p. | Rat | Formalin, CFA, carrageenan, CCI, SNL, visceral, skin incision | [28,37,40,42,43,44,45] | ||||
| Periphery | |||||||
| mGluR1/5 | Agonist | DHPG | i.pl. | Rat | Normal | Facilitation | [47,48] |
| Mouse | [49] | ||||||
| mGluR1 | NAM | CPG, CPCCOEt, LY367385 | i.pl. | Mouse | Normal | No effect | [49] |
| Rat | [48] | ||||||
| Masseter muscle | Normal, exogenous glutamate, DHPG or CHPG-induced hyperalgesia | [50] | |||||
| i.pl. | [47] | ||||||
| Mouse | Formalin | Inhibition | [49] | ||||
| Capsaicin | |||||||
| Rat | [48] | ||||||
| IL-1β-injection | [51] | ||||||
| mGluR5 | Agonist | CHPG | i.pl. | Rat | Normal | Facilitation | [47] |
| NAM | MPEP, SIB1893 | i.pl. | Mouse | Normal | No effect | [49] | |
| Rat | [48] | ||||||
| Masseter muscle | Normal, exogenous glutamate, DHPG or CHPG-induced hyperalgesia | Inhibition | [50] | ||||
| i.pl. | [47] | ||||||
| CFA, carrageenan | |||||||
| Skin incision | [52] | ||||||
| Mouse | Formalin | [49] | |||||
| Capsaicin | |||||||
| Rat | [48] | ||||||
| IL-1β-injection | [51] | ||||||
| Photoswitchable NAM | JF-NP-26 | Light-mediated activation on the paw | Mouse | Formalin | Inhibition | [53] | |
| Spinal | |||||||
| mGluR1/5 | Agonist | DHPG, 1S,3R-ACPD | i.th. | Rat | Normal | Facilitation | [54,55,56,57,58] |
| Mouse | [59,60,61] | ||||||
| Sheep | [62] | ||||||
| Mouse | CFA | [60] | |||||
| Rat | Inhibition | [58] | |||||
| Formalin | Facilitation | [63] | |||||
| Antagonist | LY393053, S-4C3HPG, AIDA | i.th. | Rat | SNI, CFA, CCI | Inhibition | [64,65,66,67,68] | |
| mGluR1 | NAM | CPCCOEt | i.th | Mouse | Formalin | Inhibition | [59] |
| Rat | CCI | [56,67] | |||||
| mGluR5 | NAM | Fenobam, MPEP | i.th. | Mouse | Formalin | Inhibition | [59] |
| Rat | SNI, CFA, CCI, paclitaxel, streptozotocin | [56,64,65,67,69,70] | |||||
| Brain | |||||||
| mGluR1/5 | Agonist | DHPG | CeA | Rat | Normal | Facilitation | [71,72] |
| dlPAG | |||||||
| Mouse | [73] | ||||||
| Formalin | Inhibition | [74] | |||||
| Antagonist | AIDA | dlPAG | Mouse | Normal | Inhibition | [73] | |
| mGluR1 | NAM | CPCCOEt | CeA | Rat | K/C-monoarthritis | Inhibition | [75] |
| BLA | Carrageenan | [76] | |||||
| mGluR5 | NAM | MPEP, MTEP | CeA | Rat | K/C-monoarthritis | Inhibition | [75] |
| Dorsal striatum | No effect | ||||||
| BLA | Carrageenan | [76] | |||||
| mPFC | SNL | Inhibition | [77] | ||||
| Photoswitchable NAM | Alloswitch-1 | Amygdala | Mouse | CFA | Inhibition | [78] | |
| JF-NP-26 | Light-mediated activation in the VPM of the thalamus | CCI, formalin | [53] | ||||
| Lentivirus | EF1α-mGluR5-IRES-Zsgreen1 | mPFC | Rat | Normal | Inhibition | [77] | |
| PAM | VU0360172 | mPFC | Rat | K/C-monoarthritis | Inhibition | [79] | |
| ACC | No effect | ||||||
| Receptor | Drug Type | Compound | Route of Administration | Species | Pain Model | Effect | References |
|---|---|---|---|---|---|---|---|
| Systemic | |||||||
| mGluR2/3 | Endogenous activation | NAC | i.p. | Mouse | Normal | Inhibition | [94] |
| Formalin, CCI, CFA | [95] | ||||||
| Agonist | LY379268, LY354740, LY389795 | Rat | Normal | No effect | [96,97] | ||
| Formalin, carrageenan, SNL, K/C-monoarthritis | Inhibition | [96,97,98] | |||||
| Mouse | CCI | [45] | |||||
| LY2969822 | Oral | Rat | Formalin, capsaicin, CFA, SNL, skin incision, visceral pain | [99] | |||
| Mouse | Acid acetic writhing | ||||||
| mGluR2 knock-out | Endogenous activation | NAC | i.p. | Mouse | Formalin | Inhibition | [95] |
| mGluR3 knock-out | Agonist | LY379268 | No effect | [100] | |||
| mGluR2 | Receptor expression Potentiation | LAC | s.c., oral | Rat | Normal | Inhibition | [101] |
| CCI | |||||||
| Carrageenan (synergism with PEA) | [102] | ||||||
| Mouse | CFA, streptozotocin | [103] | |||||
| SAHA | s.c. | Formalin | [104] | ||||
| mGluR3 | Receptor activator (NAAG inhibitor) | ZJ-11, ZJ-43 or 2-PMPA | i.p., i.v. | Rat | Formalin, SNL | Inhibition | [105,106] |
| Periphery | |||||||
| mGluR2/3 | Agonist | LY314582, 2R,4R-APDC, L-CCG-I, SLx-3095-1, DCG4 | i.pl. | Rat | Normal | No effect | [47,48,107,108] |
| Mouse | [109,110] | ||||||
| Rat | Capsaicin | Inhibition | [111,112] | ||||
| Carrageenan | [108] | ||||||
| Formalin | |||||||
| Infl. soup | [107] | ||||||
| IL-1β- injection | [51] | ||||||
| Knee joint | Carrageenan-induced monoarthritis | [113] | |||||
| i.pl. | Mouse | Carrageenan, PGE2-injection | [109,110] | ||||
| Antagonist | APICA | i.pl. | Rat | Capsaicin | Facilitation | [112] | |
| LY341495 | Mouse | Carrageenan, PGE2-injection | [110] | ||||
| mGluR3 | Receptor activator (NAAG inhibitor) | ZJ-43, 2-PMPA | s.c. | Rat | Carrageenan, formalin | Inhibition | [108] |
| Spinal | |||||||
| mGluR2/3 | Agonist | L-CCG-I | i.th. | Sheep | Normal | Inhibition | [62] |
| DCG-IV | Rat | Facilitation | [114] | ||||
| SNL | Inhibition | ||||||
| 1S,3S-ACPD, 2R,4R-APDC | Normal | No effect | [54,68] | ||||
| CFA | [115] | ||||||
| Capsaicin | [68] | ||||||
| Inhibition | |||||||
| CCI | [67] | ||||||
| Antagonist | LY341495 | i.th. | Rat | CFA | Inhibition | [115] | |
| No effect | |||||||
| mGluR2 | mGluR2 expression (HDAC inhibitor) | SAHA | i.th. | Rat | Estrogen injection | Inhibition | [116] |
| mGluR3 | Receptor activator (NAAG inhibitor) | ZJ-11, ZJ-17 | i.th | Rat | Formalin, PSNL | Inhibition | [105] |
| Brain | |||||||
| mGluR2/3 | Agonist | L-CCG-I | dlPAG | Mouse | Normal | Facilitatory | [73] |
| Formalin | Inhibition | [74] | |||||
| LY379268 | CeA | Rat | K/C- monoarthritis | Inhibition | [97] | ||
| Antagonist | EGLU | Reticular nucleus of the thalamus | Rat | CFA | Inhibition | [117] | |
| Paratenial nucleus of the thalamus, medial corticohypothalamic tract region | No effect | ||||||
| mGluR3 | Receptor activator (NAAG inhibitor) | JZ-43, 2-PMPA | i.c.v. | Rat | Formalin | Inhibition | [118] |
| Locus coeruleus | [106] | ||||||
| PAG, RVM | [119] | ||||||
| Receptor | Drug Type | Compound | Route of Administration | Species | Pain Model | Effect | References |
|---|---|---|---|---|---|---|---|
| Systemic | |||||||
| mGluR4 | Agonist | LSP4-2022 | i.p. | Rat | Carrageenan | Inhibition | [145] |
| mGluR7 | PAM | AMN082 | i.p. | Rat | Normal | No effect | [45,154] |
| Carrageenan, skin incision, CCI | Inhibition | ||||||
| NAM | MMPIP, | s.c. | Mouse | SNI | Inhibition | [155] | |
| ADX71743 | Rat | Visceral pain | [156] | ||||
| mGluR8 | Agonist | DCPG | i.p. | Mouse | Carrageenan, formalin, CCI | Inhibition | [157] |
| Periphery | |||||||
| mGluR4/6/7/8 | Agonist | L-AP4, L-SOP | i.pl. | Rat | Formalin | Inhibition | [158] |
| Capsaicin | [112] | ||||||
| Knee joint | Carrageenan | [113] | |||||
| Spinal | |||||||
| mGluR4/6/7/8 | Agonist | L-AP4, ACPT-I | i.th. | Rat | Carrageenan | Inhibition | [68,159] |
| Formalin | [63,159] | ||||||
| CFA | [159] | ||||||
| Vincristine-injection | |||||||
| SNL | [160] | ||||||
| CCI | [67,159] | ||||||
| mGluR4 | Agonist | LSP4-2022 | i.th. | Mouse | Carrageenan | Inhibition | [145] |
| Rat | Normal | No effect | |||||
| PAM | PHCCC | Carrageenan, CCI | Inhibition | [159] | |||
| Photoswitchable NAM | OptoGluNAM4.1 | i.th. | Mouse | CFA | No effect | [161] | |
| mGluR4 knock-out | Agonist | LSP4-2022 | i.th. | Mouse | Carrageenan | No effect | [145] |
| mGluR4 knock-down | Antisense oligonucleotides (i.th.) | LSP4-2022 | i.p. | Rat | Carrageenan | Slight inhibition | [145] |
| mGluR7 | PAM | AMN082 | i.th. | Rat | Carrageenan, skin incision | Inhibition | [154] |
| Paclitaxel | [162] | ||||||
| Brain | |||||||
| mGluR4/6/7/8 | Agonist | L-SOP | dlPAG | Mouse | Normal | Facilitation | [73] |
| Antagonist | MSOP | CeA | Rat | Normal | No effect | [163] | |
| Carrageenan | |||||||
| mGluR4 | PAM | LSP4-2022 | Amygdala | Mouse | CFA i.pl. | Inhibition | [12] |
| Photoswitchable PAM | Optogluram | ||||||
| mGluR7 | PAM | AMN082 | CeA | Rat | K/C- monoarthritis | No effect | [164] |
| Normal | Facilitation | ||||||
| dlPAG | [165] | ||||||
| Dorsal striatum | [166] | ||||||
| SNI | Inhibition | ||||||
| Nucleus of the solitary tract | Normal | Inhibition | [167] | ||||
| Nucleus accumbens | [168] | ||||||
| mGluR8 | Agonist | DCPG | dlPAG | Mouse | Carrageenan | Inhibition | [157] |
| CeA | Rat | Carrageenan | [163] | ||||
| Dorsal striatum | SNI | [169] | |||||
| Normal | No effect | ||||||
| PAM | AZ2216052 | Dorsal striatum | Rat | Normal | No effect | [169] | |
| SNI | Inhibition | ||||||
| mGluR6 | Agonist | Homo-AMPA | dlPAG | Mouse | Normal | Inhibition | [170] |
| Streptozotocin | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazzitelli, M.; Presto, P.; Antenucci, N.; Meltan, S.; Neugebauer, V. Recent Advances in the Modulation of Pain by the Metabotropic Glutamate Receptors. Cells 2022, 11, 2608. https://doi.org/10.3390/cells11162608
Mazzitelli M, Presto P, Antenucci N, Meltan S, Neugebauer V. Recent Advances in the Modulation of Pain by the Metabotropic Glutamate Receptors. Cells. 2022; 11(16):2608. https://doi.org/10.3390/cells11162608
Chicago/Turabian StyleMazzitelli, Mariacristina, Peyton Presto, Nico Antenucci, Shakira Meltan, and Volker Neugebauer. 2022. "Recent Advances in the Modulation of Pain by the Metabotropic Glutamate Receptors" Cells 11, no. 16: 2608. https://doi.org/10.3390/cells11162608