
Combination of dl922-947 Oncolytic Adenovirus and G-Quadruplex Binders Uncovers Improved Antitumor Activity in Breast Cancer

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Adenovirus
2.2. Sulforhodamine B (SRB) Cytotoxicity Assays
2.3. G4 Motif Formation and Cell Cycle Analyses
2.4. AdGFP Infection
2.5. Evaluation of dl922-947 DNA Amplification
2.6. Induction of Cell Senescence
2.7. CAR Modualtion by G4 Binders and STING Signaling Modulation by G4 Binders/dl922-947
2.8. Statistical Analysis
3. Results
3.1. Anti-Proliferative Effects of B19, PDS, and dl922-947
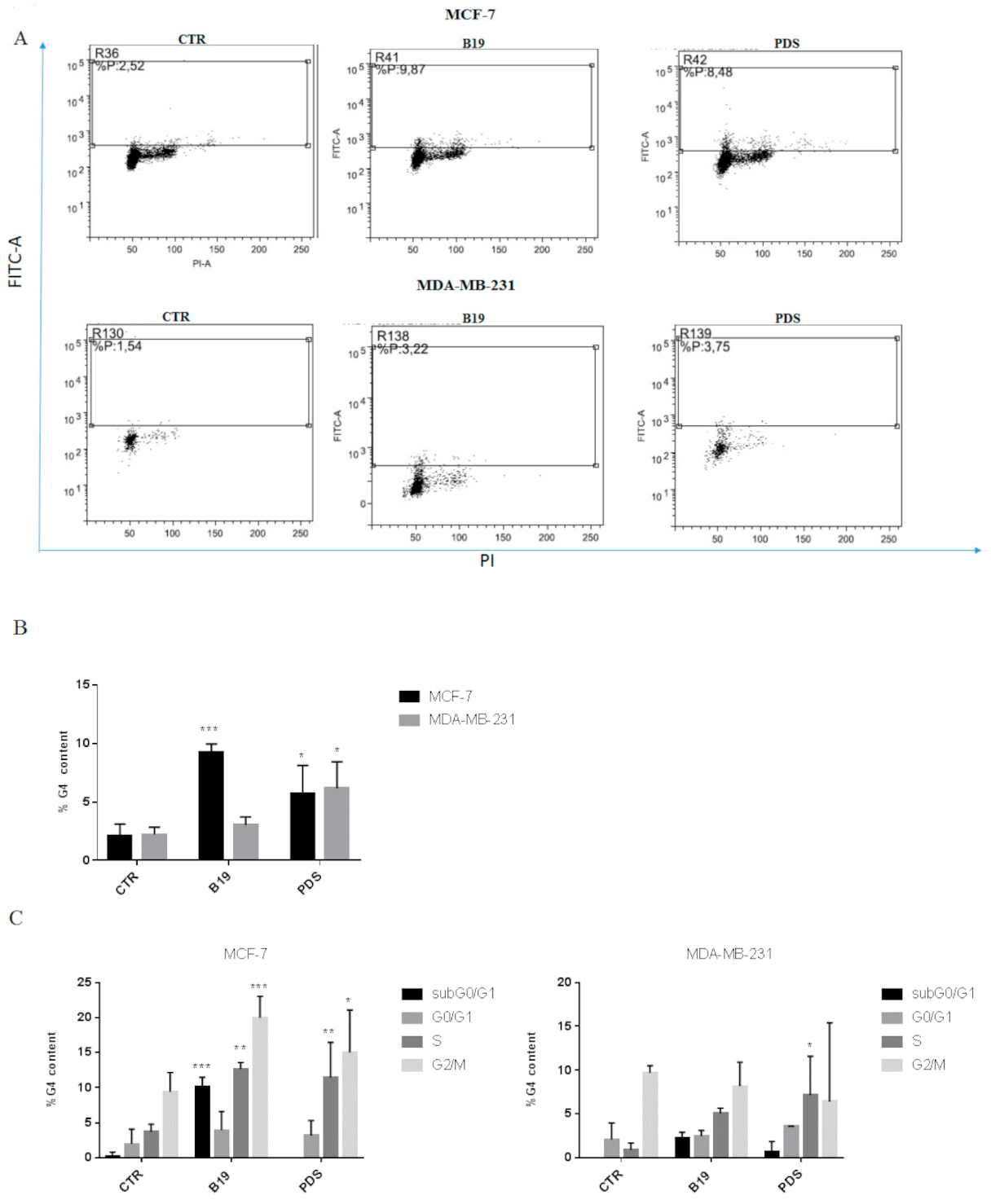
3.2. G4 Motifs in Breast Cancer Cells
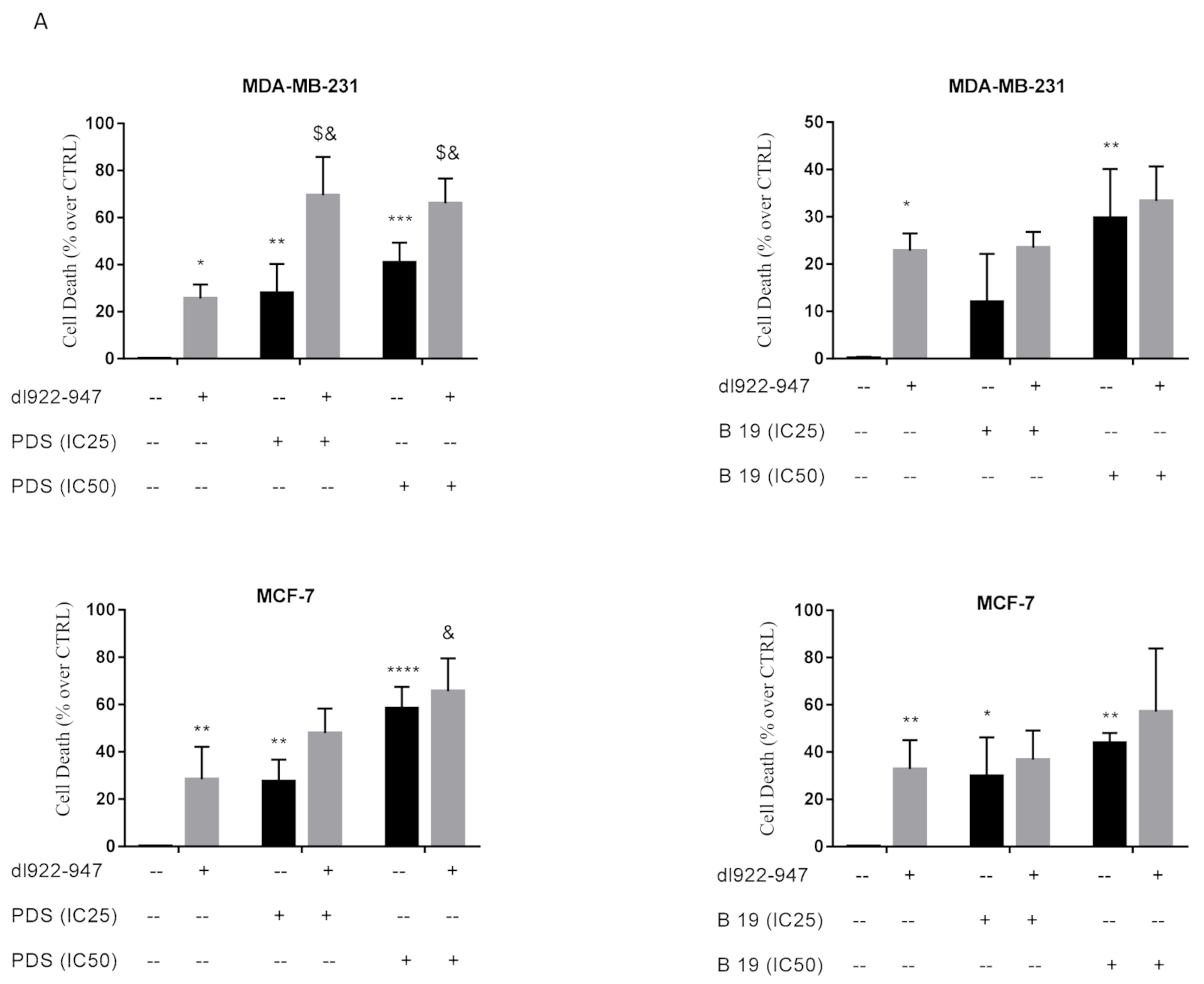
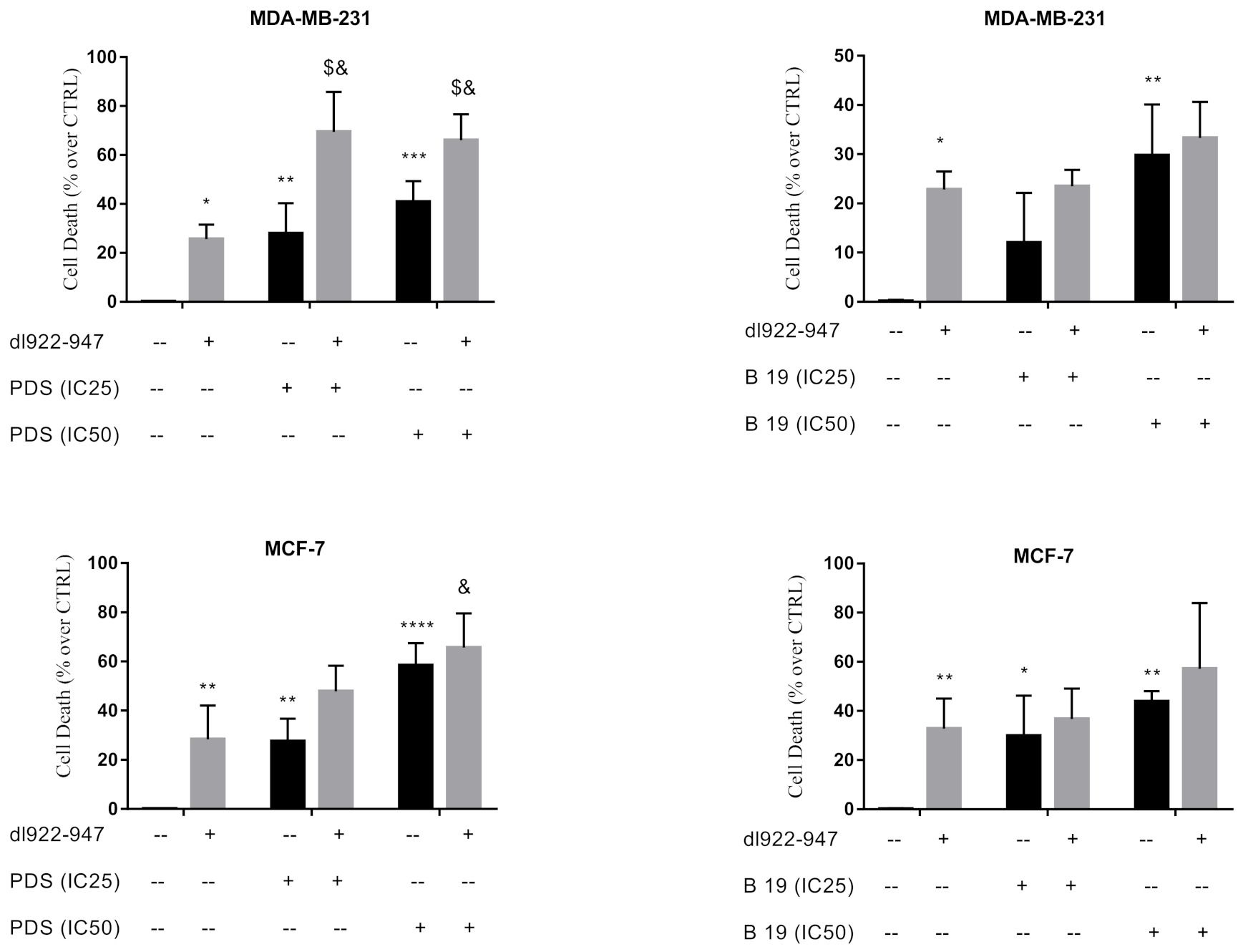
3.3. Cytotoxic Effects of dl922-947/G4 Binder Combination
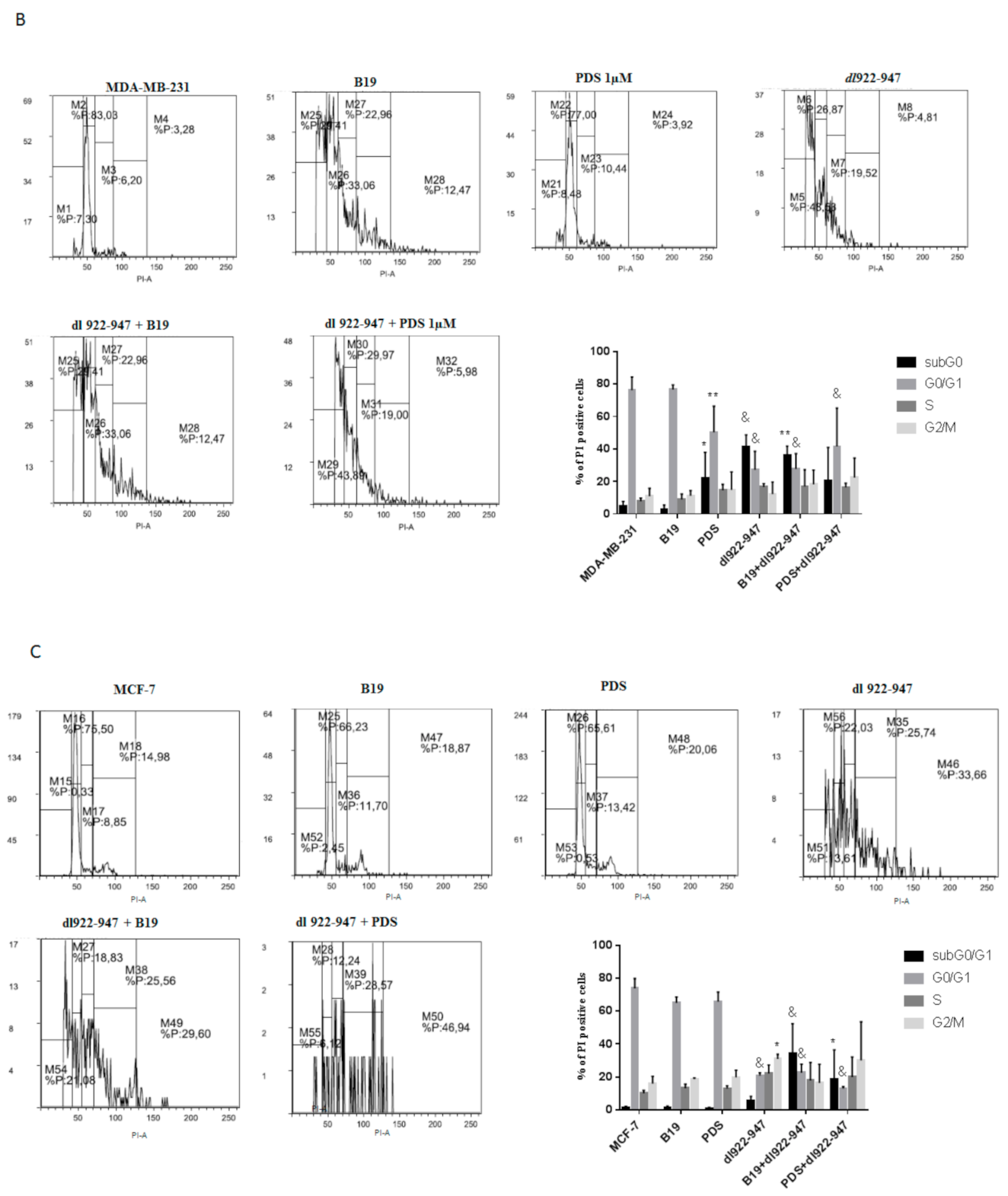
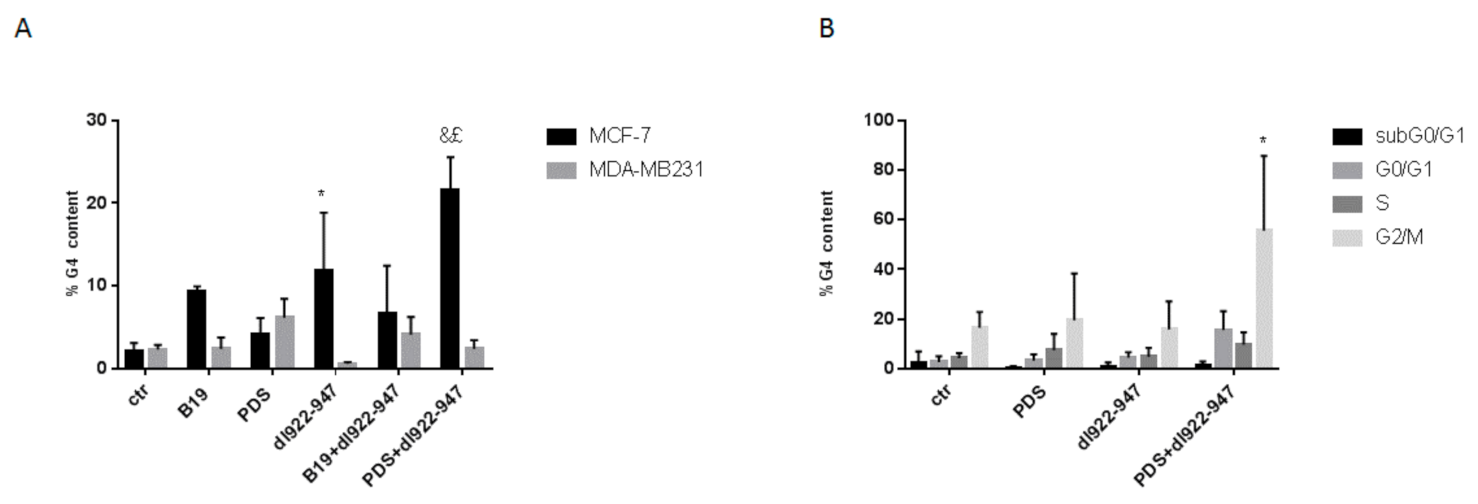
3.4. dl922-947/G4 Binder Combination and Cell Cycle
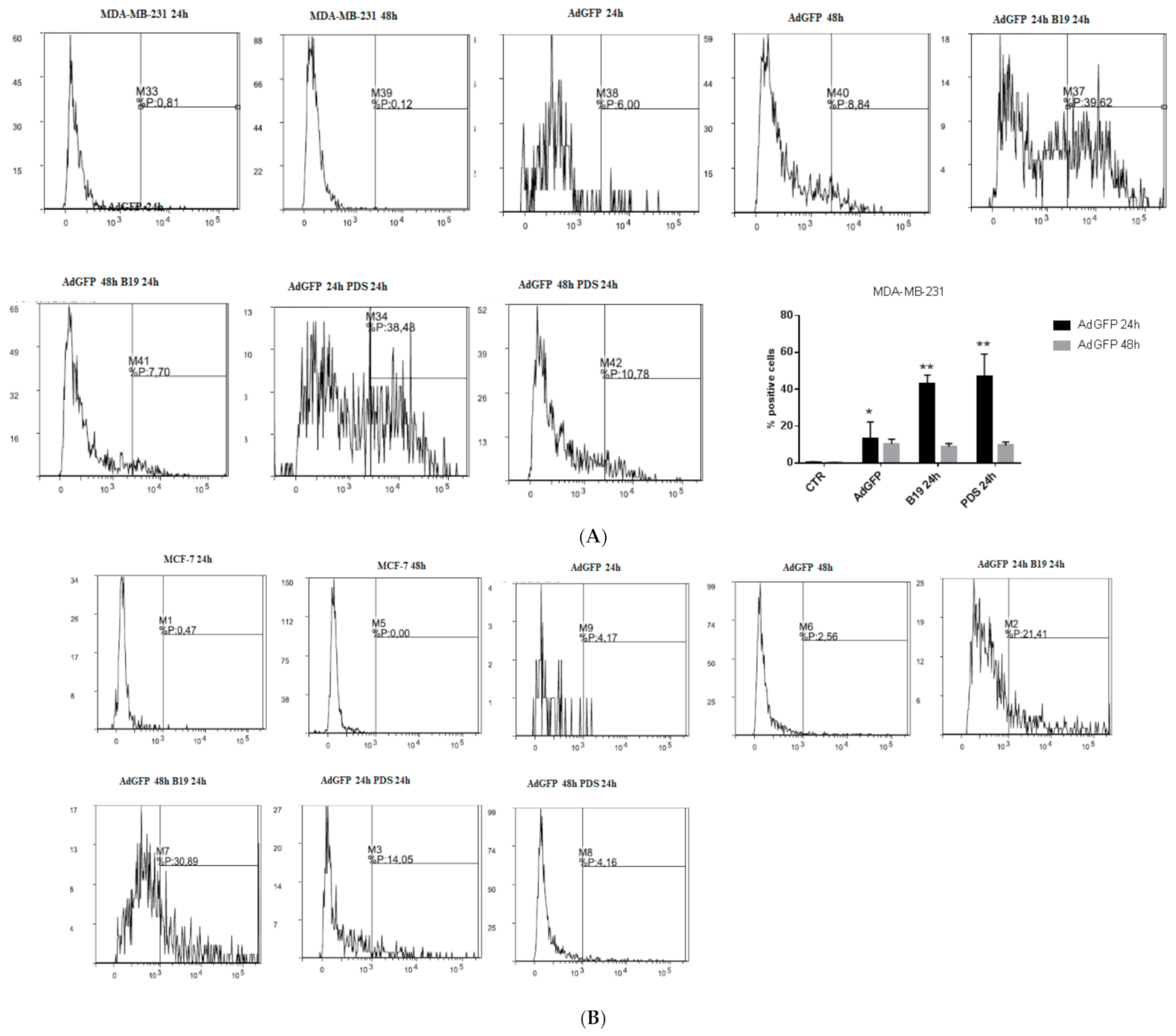
3.5. G4-Binders Enhance Adenovirus Entry in Breast Cancer Cells
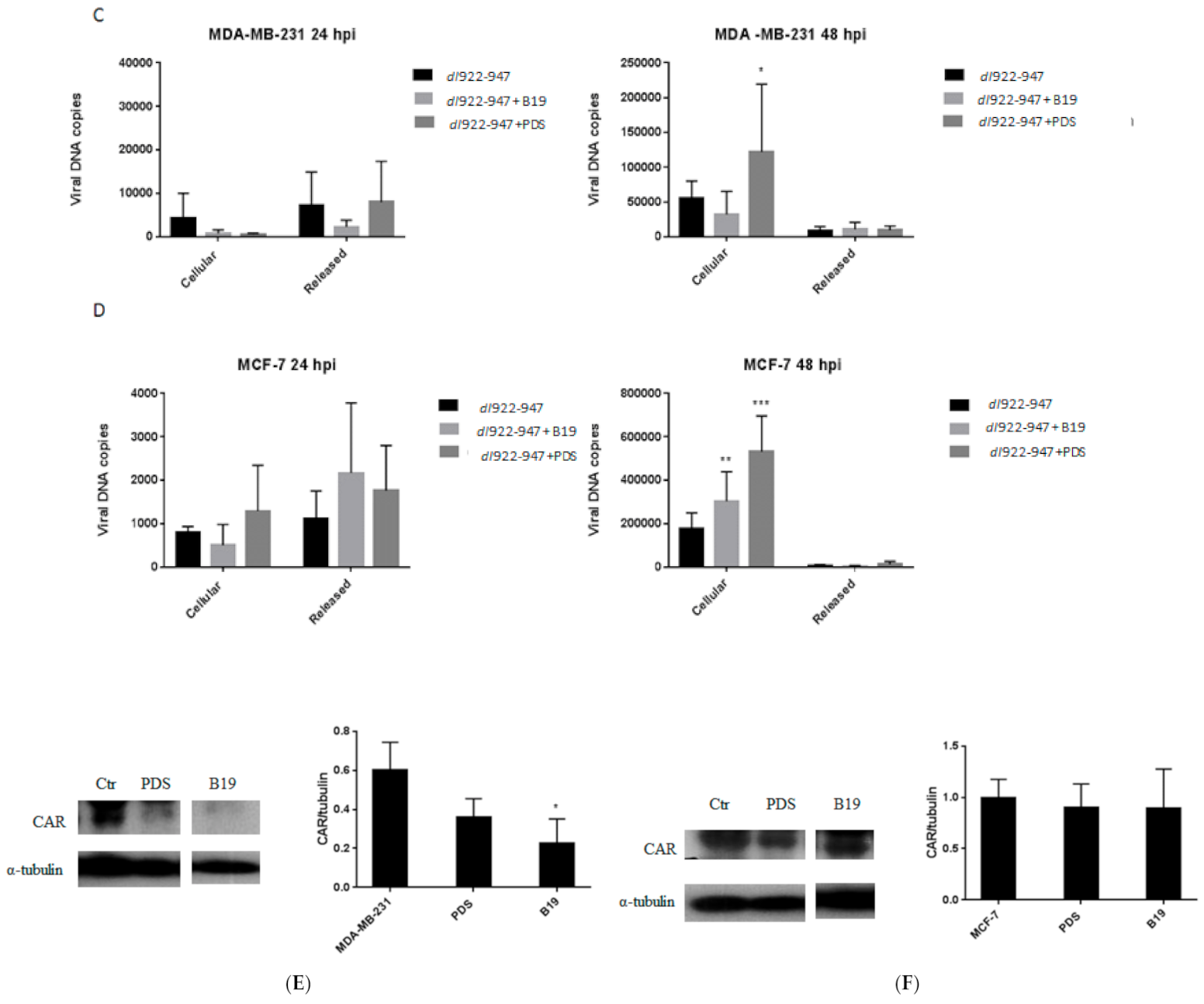
3.6. DNA Amplification of dl922-947
3.7. G4 Binder Effect on Coxsackie-Adenovirus Receptor (CAR) Expression
3.8. dl922-947 and Its Combination with PDS Induce G4 Motifs in MCF-7 Cells
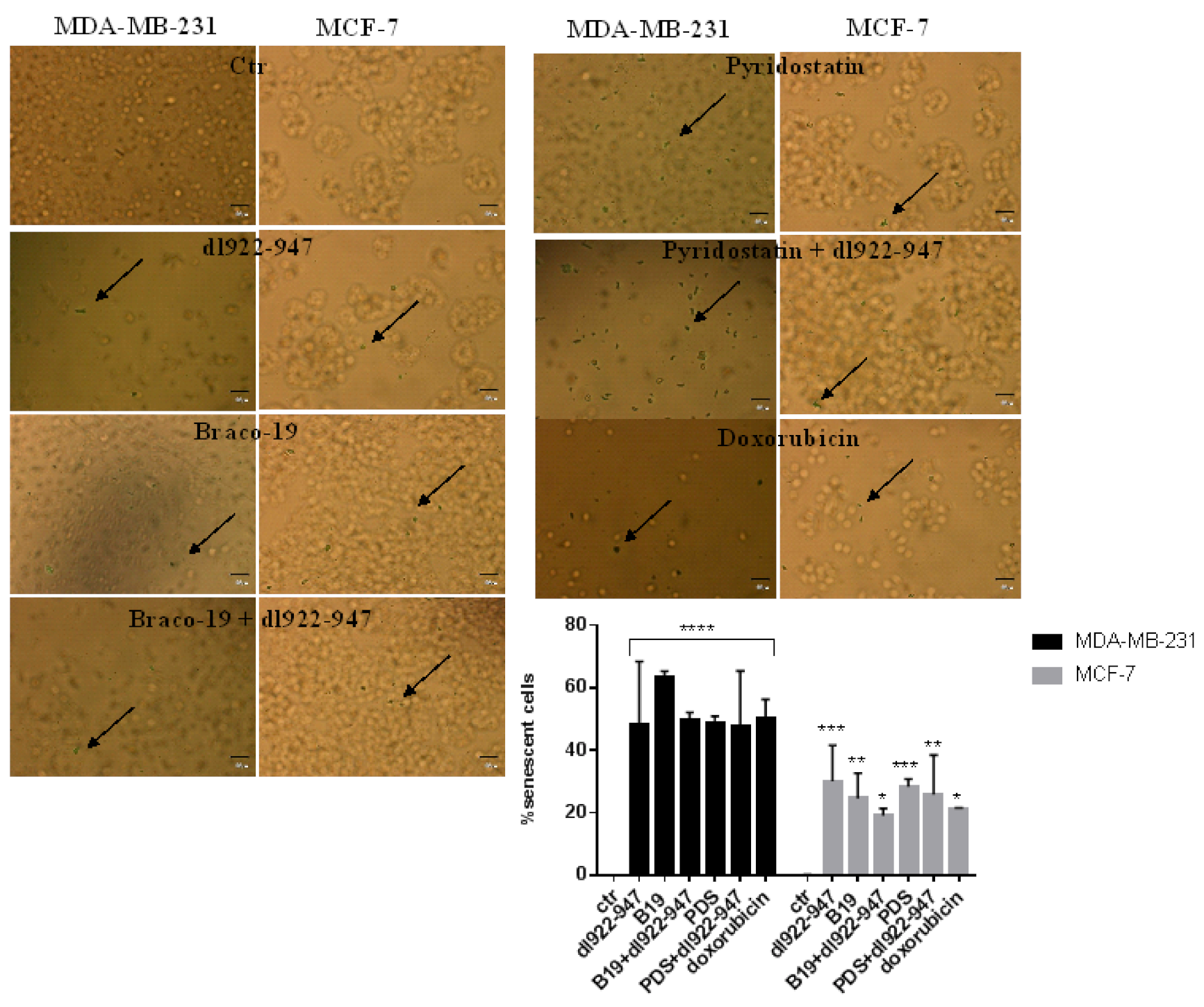
3.9. Induction of Senescence
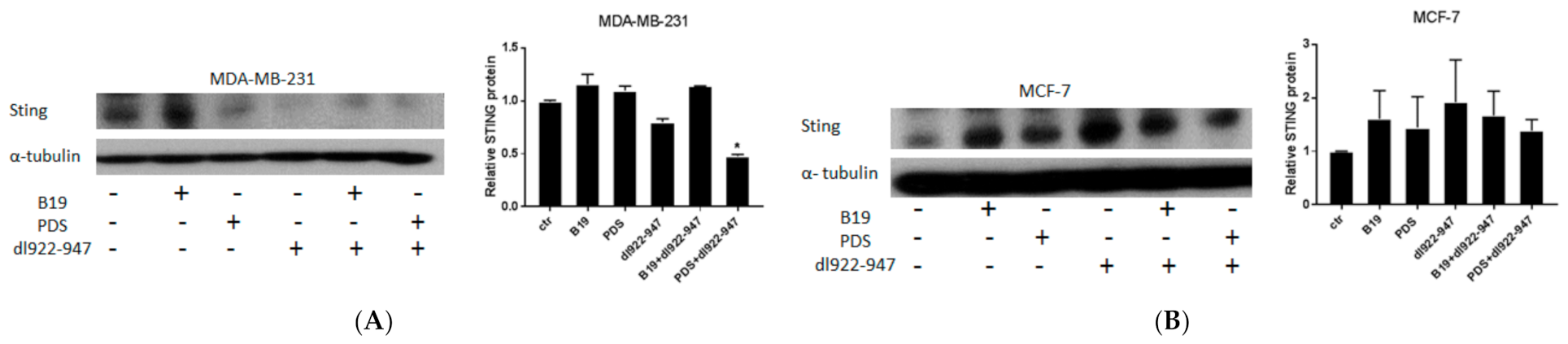
3.10. STING Induction by G4 Binders in Breast Cancer Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Amato, J.; Miglietta, G.; Morigi, R.; Iaccarino, N.; Locatelli, A.; Leoni, A.; Novellino, E.; Pagano, B.; Capranico, G.; Randazzo, A. Monohydrazone Based G-Quadruplex Selective Ligands Induce DNA Damage and Genome Instability in Human Cancer Cells. J. Med. Chem. 2020, 63, 3090–3103. [Google Scholar] [CrossRef] [PubMed]
- Amato, J.; Pagano, A.; Capasso, D.; Di Gaetano, S.; Giustiniano, M.; Novellino, E.; Randazzo, A.; Pagano, B. Targeting the BCL2 Gene Promoter G-Quadruplex with a New Class of Furopyridazinone-Based Molecules. ChemMedChem 2018, 13, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Burge, S.; Parkinson, G.N.; Hazel, P.; Todd, A.K.; Neidle, S. Quadruplex DNA: Sequence, topology and structure. Nucleic Acids Res. 2006, 34, 5402–5415. [Google Scholar] [CrossRef] [PubMed]
- Hänsel-Hertsch, R.; Di Antonio, M.; Balasubramanian, S. DNA G-quadruplexes in the human genome: Detection, functions and therapeutic potential. Nat. Rev. Mol. Cell Biol. 2017, 18, 279–284. [Google Scholar] [CrossRef]
- Nakanishi, C.; Seimiya, H. G-quadruplex in cancer biology and drug discovery. Biochem. Biophys. Res. Commun. 2020, 531, 45–50. [Google Scholar] [CrossRef]
- Ivancich, M.; Schrank, Z.; Wojdyla, L.; Leviskas, B.; Kuckovic, A.; Sanjali, A.; Puri, N. Treating Cancer by Targeting Telomeres and Telomerase. Antioxidants 2017, 6, 15. [Google Scholar] [CrossRef]
- Di Somma, S.; Amato, J.; Iaccarino, N.; Pagano, B.; Randazzo, A.; Portella, G.; Malfitano, A.M. G-Quadruplex Binders Induce Immunogenic Cell Death Markers in Aggressive Breast Cancer Cells. Cancers 2019, 11, 1797. [Google Scholar] [CrossRef]
- Malfitano, A.M.; Di Somma, S.; Iannuzzi, C.A.; Pentimalli, F.; Portella, G. Virotherapy: From single agents to combinatorial treatments. Biochem. Pharmacol. 2020, 177, 113986. [Google Scholar] [CrossRef]
- Malfitano, A.M.; Pisanti, S.; Napolitano, F.; Di Somma, S.; Martinelli, R.; Portella, G. Tumor-Associated Macrophage Status in Cancer Treatment. Cancers 2020, 15, 1987. [Google Scholar] [CrossRef]
- Di Somma, S.; Napolitano, F.; Portella, G.; Malfitano, A.M. Cross Talk of Macrophages with Tumor Microenvironment Cells and Modulation of Macrophages in Cancer by Virotherapy. Biomedicines 2021, 9, 1309. [Google Scholar] [CrossRef]
- Di Somma, S.; Iannuzzi, C.A.; Passaro, C.; Forte, I.M.; Iannone, R.; Gigantino, V.; Indovina, P.; Botti, G.; Giordano, A.; Formisano, P.; et al. The Oncolytic Virus dl922-947 Triggers Immunogenic Cell Death in Mesothelioma and Reduces Xenograft Growth. Front. Oncol. 2019, 9, 564. [Google Scholar] [CrossRef]
- Passaro, C.; Borriello, F.; Vastolo, V.; Di Somma, S.; Scamardella, E.; Gigantino, V.; Franco, R.; Marone, G.; Portella, G. The oncolytic virus dl922-947 reduces IL-8/CXCL8 and MCP-1/CCL2 expression and impairs angiogenesis and macrophage infiltration in anaplastic thyroid carcinoma. Oncotarget 2016, 7, 1500–1515. [Google Scholar] [CrossRef]
- Bazan-Peregrino, M.; Carlisle, R.C.; Hernandez-Alcoceba, R.; Iggo, R.; Homicsko, K.; Fisher, K.D.; Halldén, G.; Mautner, V.; Shen, Y.; Seymour, L.W. Comparison of molecular strategies for breast cancer virotherapy using oncolytic adenovirus. Hum. Gene Ther. 2008, 19, 873–886. [Google Scholar] [CrossRef]
- Du, W.; Searle, J.S. The rb pathway and cancer therapeutics. Curr. Drug Targets 2009, 10, 581–589. [Google Scholar] [CrossRef]
- Libertini, S.; Abagnale, A.; Passaro, C.; Botta, G.; Barbato, S.; Chieffi, P.; Portella, G. AZD1152 negatively affects the growth of anaplastic thyroid carcinoma cells and enhances the effects of oncolytic virus dl922-947. Endocr.-Relat. Cancer 2011, 18, 129–141. [Google Scholar] [CrossRef]
- Iannuzzi, C.A.; Indovina, P.; Forte, I.M.; Di Somma, S.; Malfitano, A.M.; Bruno, M.; Portella, G.; Pentimalli, F.; Giordano, A. Pharmacological Inhibition of WEE1 Potentiates the Antitumoral Effect of the dl922-947 Oncolytic Virus in Malignant Mesothelioma Cell Lines. Int. J. Mol. Sci. 2020, 21, 7333. [Google Scholar] [CrossRef]
- Passaro, C.; Volpe, M.; Botta, G.; Scamardella, E.; Perruolo, G.; Gillespie, D.; Libertini, S.; Portella, G. PARP inhibitor olaparib increases the oncolytic activity of dl922-947 in in vitro and in vivo model of anaplastic thyroid carcinoma. Mol. Oncol. 2015, 9, 78–92. [Google Scholar] [CrossRef]
- Botta, G.; Passaro, C.; Libertini, S.; Abagnale, A.; Barbato, S.; Maione, A.S.; Hallden, G.; Beguinot, F.; Formisano, P.; Portella, G. Inhibition of autophagy enhances the effects of E1A-defective oncolytic adenovirus dl922-947 against glioma cells in vitro and in vivo. Hum. Gene Ther. 2012, 23, 623–634. [Google Scholar] [CrossRef]
- Libertini, S.; Iacuzzo, I.; Perruolo, G.; Scala, S.; Ieranò, C.; Franco, R.; Hallden, G.; Portella, G. Bevacizumab increases viral distribution in human anaplastic thyroid carcinoma xenografts and enhances the effects of E1A-defective adenovirus dl922-947. Clin. Cancer Res. 2008, 14, 6505–6514. [Google Scholar] [CrossRef]
- Passaro, C.; Abagnale, A.; Libertini, S.; Volpe, M.; Botta, G.; Cella, L.; Pacelli, R.; Halldèn, G.; Gillespie, D.; Portella, G. Ionizing radiation enhances dl922-947-mediated cell death of anaplastic thyroid carcinoma cells. Endocr.-Relat. Cancer 2013, 20, 633–647. [Google Scholar] [CrossRef]
- Rodriguez, R.; Miller, K.M.; Forment, J.V.; Bradshaw, C.R.; Nikan, M.; Britton, S.; Oelschlaegel, T.; Xhemalce, B.; Balasubramanian, S.; Jackson, S.P. Small-molecule-induced DNA damage identifies alternative DNA structures in human genes. Nat. Chem. Biol. 2012, 8, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Botta, G.; Perruolo, G.; Libertini, S.; Cassese, A.; Abagnale, A.; Beguinot, F.; Formisano, P.; Portella, G. PED/PEA-15 modulates coxsackievirus-adenovirus receptor expression and adenoviral infectivity via ERK-mediated signals in glioma cells. Hum. Gene Ther. 2010, 21, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Malfitano, A.M.; Sosa, S.; Laezza, C.; De Bortoli, M.; Tubaro, A.; Bifulco, M. Rimonabant reduces keratinocyte viability by induction of apoptosis and exerts topical anti-inflammatory activity in mice. Br. J. Pharmacol. 2011, 162, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Manna, S.; Florio, D.; Iacobucci, I.; Napolitano, F.; Benedictis, I.; Malfitano, A.M.; Monti, M.; Ravera, M.; Gabano, E.; Marasco, D. A Comparative Study of the Effects of Platinum (II) Complexes on β-Amyloid Aggregation: Potential Neurodrug Applications. Int. J. Mol. Sci. 2021, 22, 3015. [Google Scholar] [CrossRef] [PubMed]
- Laezza, C.; Malfitano, A.M.; Di Matola, T.; Ricchi, P.; Bifulco, M. Involvement of Akt/NF-κB pathway in N6-isopentenyladenosine-induced apoptosis in human breast cancer cells. Mol. Carcinog. 2010, 49, 892–901. [Google Scholar] [CrossRef] [PubMed]
- Malfitano, A.M.; Laezza, C.; Pisanti, S.; Gazzerro, P.; Bifulco, M. Rimonabant (SR141716) exerts anti-proliferative and immunomodulatory effects in human peripheral blood mononuclear cells. Br. J. Pharmacol. 2008, 153, 1003–1010. [Google Scholar] [CrossRef] [PubMed]
- Pisanti, S.; Picardi, P.; Ciaglia, E.; Margarucci, L.; Ronca, R.; Giacomini, A.; Malfitano, A.M.; Casapullo, A.; Laezza, C.; Gazzerro, P.; et al. Antiangiogenic effects of N6-isopentenyladenosine, an endogenous isoprenoid end product, mediated by AMPK activation. FASEB J. 2014, 28, 1132–1144. [Google Scholar] [CrossRef]
- Ciaglia, E.; Pisanti, S.; Picardi, P.; Laezza, C.; Malfitano, A.M.; D’Alessandro, A.; Gazzerro, P.; Vitale, M.; Carbone, E.; Bifulco, M. N6-isopentenyladenosine, an endogenous isoprenoid end product, directly affects cytotoxic and regulatory functions of human NK cells through FDPS modulation. J. Leukoc. Biol. 2013, 94, 1207–1219. [Google Scholar] [CrossRef]
- De Magis, A.; Kastl, M.; Brossart, P.; Heine, A.; Paeschke, K. BG-flow, a new flow cytometry tool for G-quadruplex quantification in fixed cells. BMC Biol. 2021, 19, 45. [Google Scholar] [CrossRef]
- Martin, T.A.; Watkins, G.; Jiang, W.G. The Coxsackie-adenovirus receptor has elevated expression in human breast cancer. Clin. Exp. Med. 2005, 5, 122–128. [Google Scholar] [CrossRef]
- Lohard, S.; Bourgeois, N.; Maillet, L.; Gautier, F.; Fétiveau, A.; Lasla, H.; Nguyen, F.; Vuillier, C.; Dumont, A.; Moreau-Aubry, A.; et al. STING-dependent paracriny shapes apoptotic priming of breast tumors in response to anti-mitotic treatment. Nat. Commun. 2020, 11, 259. [Google Scholar] [CrossRef]
- Varshney, D.; Spiegel, J.; Zyner, K.; Tannahill, D.; Balasubramanian, S. The regulation and functions of DNA and RNA G-quadruplexes. Nat. Rev. Mol. Cell Biol. 2020, 21, 459–474. [Google Scholar] [CrossRef]
- Carvalho, J.; Mergny, J.L.; Salgado, G.F.; Queiroz, J.A.; Cruz, C. G-quadruplex, Friend or Foe: The Role of the G-quartet in Anticancer Strategies. Trends Mol. Med. 2020, 26, 848–861. [Google Scholar] [CrossRef]
- Ju, H.P.; Wang, Y.Z.; You, J.; Hou, X.M.; Xi, X.G.; Dou, S.X.; Li, W.; Wang, P.Y. Folding Kinetics of Single Human Telomeric G-Quadruplex Affected by Cisplatin. ACS Omega 2016, 1, 244–250. [Google Scholar] [CrossRef]
- Mach, N.; Gao, J.; Schaffarczyk, L.; Janz, S.; Ehrke-Schulz, E.; Dittmar, T.; Ehrhardt, A.; Zhang, W. Spectrum-Wide Exploration of Human Adenoviruses for Breast Cancer Therapy. Cancers 2020, 12, 1403. [Google Scholar] [CrossRef]
- Vindrieux, D.; Le Corre, L.; Hsieh, J.T.; Métivier, R.; Escobar, P.; Caicedo, A.; Brigitte, M.; Lazennec, G. Coxsackie and adenovirus receptor is a target and a mediator of estrogen action in breast cancer. Endocr.-Relat. Cancer 2011, 18, 311–321. [Google Scholar] [CrossRef]
- Long, W.; Zheng, B.X.; Li, Y.; Huang, X.H.; Lin, D.M.; Chen, C.C.; Hou, J.Q.; Ou, T.M.; Wong, W.L.; Zhang, K.; et al. Rational design of small-molecules to recognize G-quadruplexes of c-MYC promoter and telomere and the evaluation of their in vivo antitumor activity against breast cancer. Nucleic Acids Res. 2022, 50, 1829–1848. [Google Scholar] [CrossRef]
- Loo, T.M.; Miyata, K.; Tanaka, Y.; Takahashi, A. Cellular senescence and senescence-associated secretory phenotype via the cGAS-STING signaling pathway in cancer. Cancer Sci. 2020, 111, 304–311. [Google Scholar] [CrossRef]
- Miglietta, G.; Russo, M.; Duardo, R.C.; Capranico, G. G-quadruplex binders as cytostatic modulators of innate immune genes in cancer cells. Nucleic Acids Res. 2021, 49, 6673–6686. [Google Scholar] [CrossRef]
- Hong, C.; Schubert, M.; Tijhuis, A.E.; Requesens, M.; Roorda, M.; van den Brink, A.; Ruiz, L.A.; Bakker, P.L.; van der Sluis, T.; Pieters, W.; et al. cGAS-STING drives the IL-6-dependent survival of chromosomally instable cancers. Nature 2022, 607, 366–373. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | |||
| MDA-MB-231 | 1.1 µM | ||
| MCF-7 | 2.5 µM | ||
| BHT-101 | 3.1 µM | ||
| PC3 | 4.6 µM | ||
| B | |||
| MDA-MB-231 | MCF-7 | ||
| B19 IC25 | 0.3 µM | 1.2 µM | |
| B19 IC50 | 1.1 µM | 2.5 µM | |
| PDS IC25 | 0.5 µM | 0.8 µM | |
| PDS IC50 | 1.0 µM | 1.5 µM | |
| dl922-947 IC25 | 0.5 p.f.u. | 0.1 p.f.u. | |
| dl922-947 IC50 | 0.8 p.f.u. | 0.3 p.f.u. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Napolitano, F.; Di Somma, S.; Castellano, G.; Amato, J.; Pagano, B.; Randazzo, A.; Portella, G.; Malfitano, A.M. Combination of dl922-947 Oncolytic Adenovirus and G-Quadruplex Binders Uncovers Improved Antitumor Activity in Breast Cancer. Cells 2022, 11, 2482. https://doi.org/10.3390/cells11162482
Napolitano F, Di Somma S, Castellano G, Amato J, Pagano B, Randazzo A, Portella G, Malfitano AM. Combination of dl922-947 Oncolytic Adenovirus and G-Quadruplex Binders Uncovers Improved Antitumor Activity in Breast Cancer. Cells. 2022; 11(16):2482. https://doi.org/10.3390/cells11162482
Chicago/Turabian StyleNapolitano, Fabiana, Sarah Di Somma, Giuliano Castellano, Jussara Amato, Bruno Pagano, Antonio Randazzo, Giuseppe Portella, and Anna Maria Malfitano. 2022. "Combination of dl922-947 Oncolytic Adenovirus and G-Quadruplex Binders Uncovers Improved Antitumor Activity in Breast Cancer" Cells 11, no. 16: 2482. https://doi.org/10.3390/cells11162482