
Arms Race between the Host and Pathogen Associated with Fusarium Head Blight of Wheat

Abstract
:1. Introduction
2. Identification of Pathogenic Strains of Wheat FHB and Its Pathogenesis
3. Discovery and Identification of Wheat Resistance Germplasm Resources
4. The Resistance Mechanism of Wheat to FHB
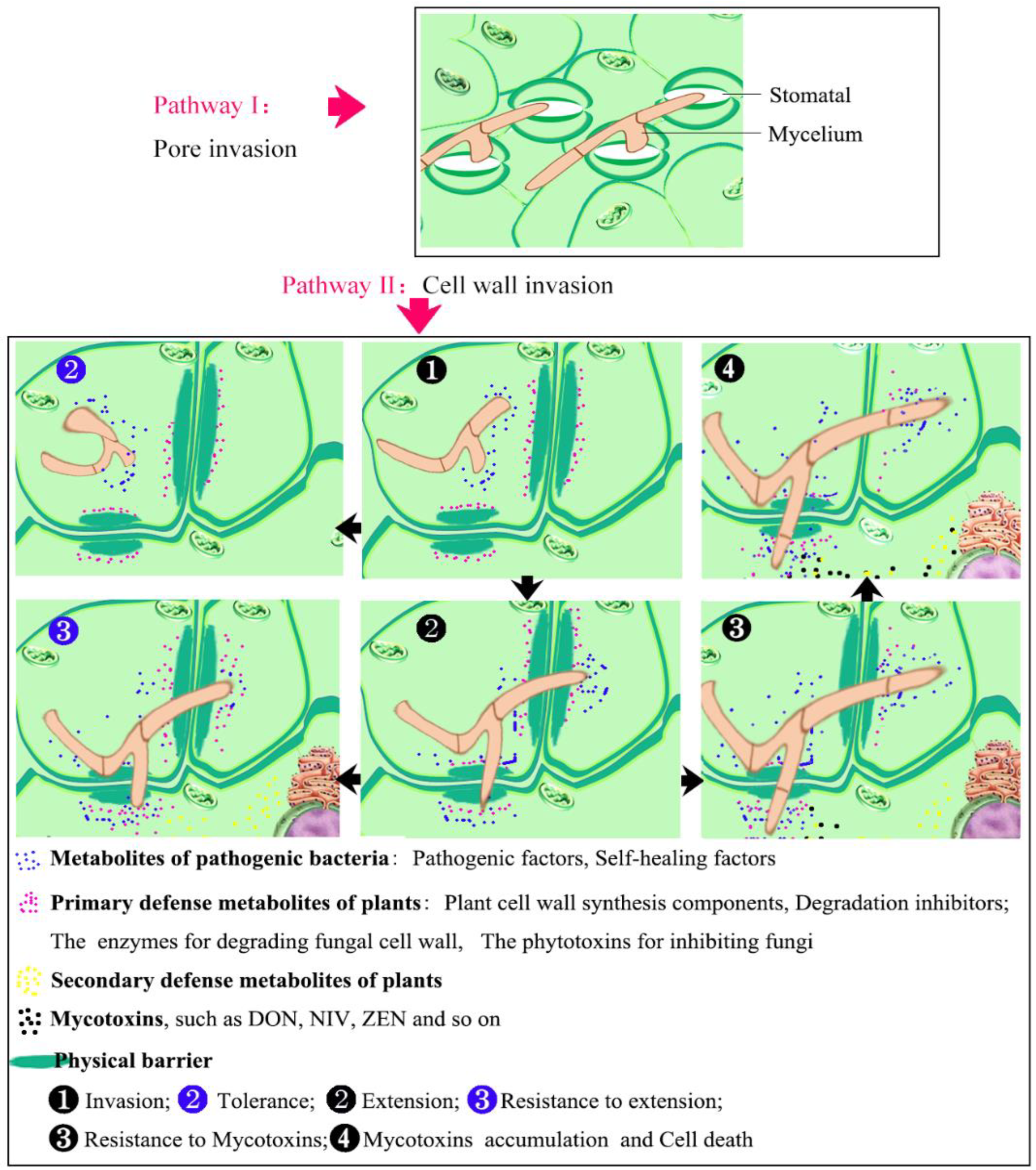
4.1. Morphological and Physiological Mechanisms of Wheat Response to FHB
4.2. Resistance QTL in Plant to FHB
4.3. Resistance Genes and Their Mechanisms in Response to FHB in Plant
4.4. CERK1-Mediated ROS Homeostasis Perhaps Performs a Vital Role in Response to FHB in Plant
5. Biological and Chemical Control
5.1. Biological Control
5.2. Chemical Control
6. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AMP | adenosine monophosphate |
| ACT | agmatinecoumaroyl transferase |
| AOS | allene oxide synthase |
| BIK1 | botrytis-induced kinase 1 |
| CDPK | calcium-dependent protein kinase |
| CEBiP | chitin elicitor-binding protein |
| DMI | demethylase inhibitor |
| DON | deoxynivalenol |
| DGK | diacylglycerol kinase |
| ET | ethylene |
| Fg | Fusarium graminearum |
| FHB | Fusarium head blight |
| GWAS | genome-wide association studies |
| GLI1 | glycerol kinase |
| GEF | guanine nucleotide exchange factor |
| GMP | guanosine monophosphate |
| G protein | GTP binding protein |
| HCAAs | hydroxycinnamic acid amides |
| IMP | inosine monophosphate |
| JA | jasmonic acid |
| LYK | LysM-containing receptor-like kinase |
| MAPK | mitogen-activated protein kinase |
| NAC | N-acetylcysteine-like transcription factor |
| NIV | nivalenol |
| NOX/RBOH | NADPH oxidase/respiratory burst oxidase homolog |
| Osp24 | orphan protein secreted from FHB-causing fungi |
| FROG | orphan protein secreted from plant under FHB stress |
| PAMP | pathogen-associated molecular pattern |
| PRR | pattern recognition receptors |
| PBL | PBS-like kinase |
| PLD | phenylpropanoid |
| PA | phosphatidic acid |
| NUT | plant SCREW unresponsive receptor kinase |
| PG | polygalacturonase |
| PFT | pore-forming toxin-like |
| PGIP | polygalacturonase inhibitory protein |
| QTL | quantitative trait loci |
| ROS | reactive oxygen species |
| RACK | receptor for activated C-kinase |
| RLCK | receptor-like cytoplasmic kinases |
| RLK | receptor-like protein kinase |
| RRI | resistance related induced |
| ROP/RAC | Rho-type GTPases, called ROPs (Rho of plants) or RACs (for the sequence similarity they share with animal Racs, a Rho subfamily) |
| SNP | single-nucleotide polymorphism |
| SNARE | soluble N-ethylmaleimide-sensitive factor attachment protein receptor |
| SCREW | small phytocytokines regulating defense and water loss |
| SnRK1 | SNF1-related protein kinase family 1 |
| TaCERK | chitin elicitor receptor kinase in wheat |
| UGT | uridine diphosphate (UDP)-glucose transferases |
| WAK | wall-associated kinases |
| WRKY | a plant-specific transcription factor family named for its conserved sequence consisting of 7 amino acids: WRKYGQK |
| ZEN | zearalenol |
References
- Covarelli, L.; Beccari, G.; Prodi, A.; Generotti, S.; Etruschi, F.; Juan, C.; Ferrer, E.; Mañes, J. Fusarium Species, chemotype characterization and trichothecene contamination of durum and soft wheat in an area of central Italy. J. Sci. Food Agric. 2015, 95, 540–551. [Google Scholar] [CrossRef] [PubMed]
- He, X.Y.; Dreisigacker, S.; Singh, R.P.; Singh, P.K. Genetics for low correlation between fusarium head blight disease and deoxynivalenol (DON) content in a bread wheat mapping population. Theor. Appl. Genet. 2019, 132, 2401–2411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.Q.; Xie, Q.; Li, G.Q.; Jia, H.Y.; Zhou, J.Y.; Kong, Z.X.; Li, N.; Yuan, Y. Germplasms, genetics and genomics for better control of disastrous wheat fusarium head blight. Theor. Appl. Genet. 2020, 133, 1541–1568. [Google Scholar] [CrossRef]
- Brisco, E.I.; Brown, L.K.; Olson, E.L. Fusarium head blight resistance in Aegilops Tauschii. Genet. Resour. Crop Evol. 2017, 64, 2049–2058. [Google Scholar] [CrossRef]
- Zhang, A.M.; Yang, W.L.; Li, X.; Sun, J.Z. Current status and perspective on research against fusarium head blight in wheat. Hereditas 2018, 40, 858–873. [Google Scholar] [CrossRef]
- Bai, G.H.; Shaner, G.E. Variation in Fusarium graminearum and cultivar resistance to wheat scab. Plant Dis. 1996, 80, 975–979. [Google Scholar] [CrossRef]
- Frohberg, R.C.; Stack, R.W.; Olson, T.; Miller, J.D.; Megoum, M. Registration of ‘Alsen’ Wheat. Crop Sci. 2006, 46, 2311–2312. [Google Scholar] [CrossRef]
- Nishio, Z.; Takata, K.; Tabiki, T.; Ito, M.; Takenaka, S.; Kuwabara, T.; Iriki, N.; Ban, T. Diversity of resistance to fusarium head blight in japanese winter wheat. Breed. Sci. 2004, 54, 79–84. [Google Scholar] [CrossRef] [Green Version]
- Badea, A.; Eudes, F.; Graf, R.J.; Laroche, A.; Gaudet, D.A.; Sadasivaiah, R.S. Phenotypic and marker-assisted evaluation of spring and winter wheat germplasm for resistance to fusarium head blight. Euphytica 2008, 164, 803–819. [Google Scholar] [CrossRef]
- Bai, G.H.; Su, Z.Q.; Cai, J. Wheat resiatance to fusarium head blight. Can. J. Plant Pathol. 2018, 40, 336–346. [Google Scholar] [CrossRef]
- Bluhm, B.H.; Zhao, X.; Flaherty, J.E.; Xu, J.R.; Dunkle, L.D. RAS2 regulates growth and pathogenesis in Fusarium graminearum. Mol. Plant Microbe Interact. 2007, 20, 627–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, L.J.; Tang, H.Y.; Wang, W.Q.; Yuan, T.L.; Wei, W.Q.; Pang, B.; Gong, X.M.; Wang, S.F.; Li, Y.J.; Zhang, D.; et al. A linear nonribosomal octapeptide from Fusarium graminearum facilitates cell-to-cell invasion of wheat. Nat. Commun. 2019, 10, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palacios, S.A.; Canto, A.D.; Erazo, J.; Torres, A.M. Fusarium cerealis causing fusarium head blight of durum wheat and its associated mycotoxins. Int. J. Food Microbiol. 2021, 346, 109161. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Chi, H.W.; Li, W.D.; Zhang, L.; Zhang, L.Y.; Chen, L.; Zou, S.S.; Liu, H.X.; Liang, Y.C.; Yu, J.F.; et al. FgPsd2, a phosphatidylserine decarboxylase of Fusarium graminearum, regulates development and virulence. Fungal Genet. Biol. 2021, 146, 103483. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Gardiner, D.M.; Kazan, K. Transcriptome analysis reveals infection strategies employed by Fusarium graminearum as a root pathogen. Microbiol. Res. 2022, 256, 126951. [Google Scholar] [CrossRef]
- Spanic, V.; Cosic, J.; Zdunic, Z.; Drezner, G. Characterization of agronomical and quality traits of winter wheat (Triticum aestivum L.) for fusarium head blight pressure in different environments. Agronomy 2021, 11, 213. [Google Scholar] [CrossRef]
- Drakopoulos, D.; Kgi, A.; Six, J.; Zorn, A.; Vogelgsang, S. The agronomic and economic viability of innovative cropping systems to reduce fusarium head blight and related mycotoxins in wheat. Agric. Syst. 2021, 192, 103198. [Google Scholar] [CrossRef]
- Senatore, M.T.; Ward, T.J.; Cappelletti, E.; Beccari, G.; Prodi, A. Species diversity and mycotoxin production by members of the fusarium tricinctum species complex associated with fusarium head blight of wheat and barley in Italy. Int. J. Food Microbiol. 2021, 358, 109298. [Google Scholar] [CrossRef]
- Abaya, A.; Serajazari, M.; Hsiang, T. Control of fusarium head blight using the endophytic fungus, Simplicillium lamellicola, and its effect on the growth of Triticum aestivum. Biol. Control 2021, 160, 104684. [Google Scholar] [CrossRef]
- Pellan, L.; Dieye, C.A.T.; Durand, N.; Fontana, A.; Schorr-Galindo, S.; Strub, C. Biocontrol agents reduce progression and mycotoxin production of Fusarium graminearum in spikelets and straws of wheat. Toxins 2021, 13, 597. [Google Scholar] [CrossRef]
- Goncharov, A.A.; Gorbatova, A.S.; Sidorova, A.A.; Tiunov, A.V.; Bocharov, G.A. Mathematical modelling of the interaction of winter wheat (Triticum aestivum L.) and fusarium species (Fusarium spp.). Ecol. Model. 2022, 465, 109856. [Google Scholar] [CrossRef]
- Li, T.; Bai, G.H.; Wu, S.Y.; Gu, S.L. Quantitative trait loci for resistance to fusarium head blight in the chinese wheat landrace Huangfangzhu. Euphytica 2012, 185, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.H.; Pan, H.Y.; Bai, G.H. Quantitative trait loci responsible for fusarium head blight resistance in chinese landrace baishanyuehuang. Theor. Appl. Genet. 2012, 125, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Bai, G.H. Quantitative trait loci for fusarium head blight resistance in Huangcandou× ‘jagger’ wheat population. Crop Sci. 2014, 54, 2520–2528. [Google Scholar] [CrossRef]
- Chen, S.L.; Zhang, Z.L.; Sun, Y.Y.; Li, D.S.; Gao, D.R.; Zhan, K.H.; Cheng, S.H. Identification of quantitative trait loci for fusarium head blight (FHB) resistance in the cross between wheat landrace N553 and elite cultivar Yangmai 13. Mol. Breed. 2021, 41, 1–13. [Google Scholar] [CrossRef]
- Lin, F.; Xue, Z.Z.; Zhang, C.Q.; Zhang, Z.K.; Kong, G.Q.; Yao, D.G.; Tian, D.G.; Zhu, H.L.; Li, C.J.; Cao, Y.; et al. Mapping QTL Associated with resistance to fusarium head blight in the Nanda2419×Wangshuibai population. II: Type I resistance. Theor. Appl. Genet. 2006, 112, 528–535. [Google Scholar] [CrossRef]
- Lu, W.Z.; Cheng, S.H.; Wang, Y.Z. Study on Fusarium Head Blight of Wheat, 1st ed.; Science Press: Peking, China, 2001; pp. 3–12. Available online: https://book.sciencereading.cn/shop/book/Booksimple/onlineRead.do?id=B2B4195C7F400419F83C82EA84B62B57C000&readMark=0 (accessed on 26 May 2022).
- Lewandowski, S.M.; Bushnell, W.R.; Evans, C.K. Distribution of mycelial colonies and lesions in field-grown barley inoculated with Fusarium graminearum. Phytopathology 2006, 96, 567–581. [Google Scholar] [CrossRef] [Green Version]
- Wanjiru, W.M.; Kang, Z.S.; Buchenauer, H. Importance of cell wall degrading enzymes produced by Fusarium graminearum during infection of wheat heads. Eur. J. Plant Pathol. 2002, 108, 803–810. [Google Scholar] [CrossRef]
- Xu, Y.A.; Hideki, N. The infection process of wheat SCAB pathogen. J. Nanjing Agric. Univ. 1989, 12, 33–38. [Google Scholar]
- Liu, P.; Zhang, X.X.; Zhang, F.; Xu, M.Z.; Ye, Z.X.; Wang, K.; Liu, S.; Han, X.L.; Cheng, Y.; Zhong, K.L.; et al. Virus-derived SiRNA activates plant immunity by interfering with ROS scavenging. Mol. Plant 2021, 14, 1088–1103. [Google Scholar] [CrossRef]
- Wu, B.Y.; Li, P.; Hong, X.F.; Xu, C.H.; Wang, R.; Liang, Y. The receptor-like cytosolic kinase RIPK activates NADP-malic enzyme 2 to generate NADPH for fueling the ROS production. Mol. Plant 2022, 15, 887–903. [Google Scholar] [CrossRef] [PubMed]
- Hao, G.X.; Helene, T.; Susan, M. Chitin triggers tissue-specific immunity in wheat associated with fusarium head blight. Front. Plant Sci. 2022, 13, 832502. [Google Scholar] [CrossRef] [PubMed]
- Strange, R.N.; Smith, H. Specificity of choline and betaine as stimalants of Fusarium graminearum. Trans. Br. Mycol. Soc. 1978, 70, 187–192. [Google Scholar] [CrossRef]
- Strange, R.N.; Deramo, A.; Smith, H. Virulence Enhancement of Fusarium graminearum by chloline and betaine and of Botrytis Cinerea by other constituents of wheat germ. Trans. Br. Mycol. Soc. 1978, 70, 201–207. [Google Scholar] [CrossRef]
- Walter, S.; Nicholson, P.; Doohan, F.M. Action and reaction of host and pathogen during fusarium head blight disease. New Phytol. 2010, 185, 54–66. [Google Scholar] [CrossRef]
- Ding, S.; Mehrabi, R.; Koten, C.; Kang, Z.; Wei, Y.; Seong, K.; Kistler, H.C.; Xu, J.R. Transducin beta-like gene FTL1 is essential for pathogenesis in Fusarium graminearum. Eukaryot. Cell 2009, 8, 867–876. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.Y.; Seo, J.A.; Kim, J.E.; Han, K.H.; Shim, W.B.; Yun, S.H.; Lee, Y.W. Functional analyses of heterotrimeric G protein Gα and Gβ subunits in Gibberella zeae. Microbilogy 2008, 154, 392–401. [Google Scholar] [CrossRef] [Green Version]
- Sridhar, P.S.; Trofimova, D.; Subramaniam, R.; Fundora, D.G.P.; Foroud, N.A.; Allingham1, J.S.; Loewen, M.C. Ste2 receptor-mediated chemotropism of Fusarium graminearum contributes to its pathogenicity against wheat. Sci. Rep. 2020, 10, 10770. [Google Scholar] [CrossRef]
- Chong, X.F.; Wang, C.Y.; Wang, Y.; Wang, Y.X.; Zhang, L.Y.; Liang, Y.C.; Chen, L.; Zou, S.S.; Dong, H.S. The dynamin-like GTPase FgSey1 plays a critical role in fungal development and virulence in Fusarium graminearum. Appl. Environ. Microbiol. 2020, 86, e02720-19. [Google Scholar] [CrossRef]
- Wang, C.Y.; Wang, Y.X.; Wang, Y.; Wang, Z.D.; Zhang, L.Y.; Liang, Y.C.; Chen, L.; Zou, S.S.; Dong, H.S. The ADP-ribosylation Factor-like small GTPase FgArl1 participates in growth, pathogenicity and DON production in Fusarium graminearum. Fungal Biol. 2020, 124, 969–980. [Google Scholar] [CrossRef]
- Yang, C.D.; Li, J.J.; Chen, X.; Zhang, X.Z.; Liao, D.H.; Yun, Y.Z.; Zheng, W.H.; Abubakar, Y.S.; Li, G.P.; Wang, Z.H.; et al. FgVps9, a Rab5 GEF, is critical for DON biosynthesis and pathogenicity in Fusarium graminearum. Front. Microbiol. 2020, 11, 1714. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.Q.; Ren, X.X.; Wang, X.T.; Wan, Q.; Ding, K.J.; Chen, L. FgRad50 regulates fungal development, pathogenicity, cell wall integrity and the DNA damage response in Fusarium graminearum. Front. Microbiol. 2020, 10, 2970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.M.; Dai, Y.F.; Huang, Y.; Wang, K.; Lu, P.; Xu, H.F.; Xu, J.R.; Liu, H.Q. The SR-protein FgSrp2 regulates vegetative growth, sexual reproduction and pre-mRNA processing by interacting with FgSrp1 in Fusarium graminearum. Curr. Genet. 2020, 66, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; He, D.; Liu, X.; Ji, F.; Xu, J.H.; Shi, J.R. The pyruvate dehydrogenase kinase 2 (PDK2) is associated with conidiation, mycelial growth, and pathogenicity in Fusarium graminearum. Food Prod. Process. Nutr. 2020, 2, 11. [Google Scholar] [CrossRef]
- Adnan, M.; Fang, W.Q.; Sun, P.; Zheng, Y.L.; Abubakar, Y.S.; Zhang, J.; Lou, Y.; Zheng, W.H.; Lu, G.D. R-SNARE FgSec22 is essential for growth, pathogenicity and DON production of Fusarium graminearum. Curr. Genet. 2020, 66, 421–435. [Google Scholar] [CrossRef]
- Sun, M.L.; Bian, Z.Y.; Luan, Q.Q.; Chen, Y.T.; Wang, W.; Dong, Y.R.; Chen, L.F.; Hao, C.F.; Xu, J.R.; Liu, H.Q. Stage-specific regulation of purine metabolism during infectious growth and sexual reproduction in Fusarium graminearum. New Phytol. 2021, 230, 757–773. [Google Scholar] [CrossRef]
- Wan, Y.F.; Yan, J.; Yang, J.L.; Liu, F.G. Study on resistance to fusarium head blight in wild plants related to triticale. Acta Phytopathol. Sin. 1997, 27, 107–111. [Google Scholar]
- Gagkaeva, T.Y. Importance of fusarium head blight in Russia and the search for new sources of genetic resistance in wheat and barley. In Proceedings of the National Fusarium Head Blight Forum, Bloomington, MN, USA, 13–15 December 2003; pp. 219–222. [Google Scholar]
- Qi, L.L.; Pumphrey, M.O.; Friebe, B.; Chen, P.D.; Gill, B.S. Molecular cytogenetic characterization of alien introgressions with gene Fhb3 for resistance to fusarium head blight disease of wheat. Theor. Appl. Genet. 2008, 117, 1155–1166. [Google Scholar] [CrossRef]
- Cainong, J.C.; Bockus, W.W.; Feng, Y.G.; Chen, P.D.; Qi, L.L.; Sehgal, S.K.; Danilova, T.V.; Koo, D.H.; Friebe, B.; Gill, B.S. Chromosome engineering, mapping, and transferring of resistance to fusarium head blight disease from Elymus Tsukushiensis into Wheat. Theor. Appl. Genet. 2015, 128, 1019–1027. [Google Scholar] [CrossRef]
- Su, Z.Q.; Bernardo, A.; Tian, B.; Chen, H.; Wang, S.; Ma, H.X.; Cai, S.B.; Liu, D.T.; Zhang, D.D.; Li, T.; et al. A deletion mutation in TaHRC confers Fhb1 resistance to fusarium head blight in wheat. Nat. Genet. 2019, 51, 1099–1105. [Google Scholar] [CrossRef]
- Ma, X. Cloning and Function Analysis of FHB Resistance-Related Genes from Wheat. Ph.D. Thesis, Shandong Agricultural University, Tai’an, China, 2014. [Google Scholar]
- Li, T.; Li, A.; Li, L. Fusarium head blight in wheat: From phenotyping to resistance improvement. Sci. Technol. Rev. 2016, 34, 75–80. [Google Scholar]
- Jiang, P.; Zhang, X.; Wu, L.; He, Y.; Zhuang, W.W.; Cheng, X.X.; Ge, W.Y.; Ma, H.X.; Kong, L.R. A novel QTL on chromosome 5AL of Yangmai 158 increases resistance to fusarium head blight in wheat. Plant Pathol. 2020, 69, 249–258. [Google Scholar] [CrossRef]
- Miller, J.D.; Young, J.C.; Sampson, D.R. Deoxynivalenol and fusarium head blight resistance in spring cereals. J. Phytopathol. 1985, 113, 359–367. [Google Scholar] [CrossRef]
- Mesterhazy, A. Types and components of resistance to fusarium head blight of wheat. Plant Breed. 1995, 114, 377–386. [Google Scholar] [CrossRef]
- Foroud, N.A. Investigating the Molecular Mechanisms of Fusarium Head Blight Resistance in Wheat. Ph.D. Thesis, Vancouver University British Columbia, Vancouver, BC, Canada, 2011. [Google Scholar]
- Franco, M.F.; Lori, G.A.; Cendoya, G.; Alonso, M.P.; Panelo, J.S.; Malbra’n, I.; Mirabella, N.E.; Pontaroli, A.C. Spike architecture traits associated with type II resistance to fusarium head blight in bread wheat. Euphytica 2021, 217, 209. [Google Scholar] [CrossRef]
- Lu, Q.X.; Lillemo, M.; Skinnes, H.; He, X.Y.; Shi, J.R. Anther extrusion and plant height are associated with type I resistance to fusarium head blight in bread wheat line ‘Shanghai-3/Catbird’. Theor. Appl. Genet. 2013, 126, 317–334. [Google Scholar] [CrossRef]
- Liu, S.; Griffey, C.A.; Hall, M.D.; McKendry, A.L.; Chen, J.L.; Brooks, W.S.; Brown-Guedira, G.; Sanford, D.V.; Schmale, D.G. Molecular characterization of field resistance to fusarium head blight in two US soft red winter wheat cultivars. Theor. Appl. Genet. 2013, 126, 2485–2498. [Google Scholar] [CrossRef] [Green Version]
- Lionetti, V.; Giancaspro, A.; Fabri, E.; Giove, S.L.; Reem, N.; Zabotina, O.A.; Blanco, A.; Gadaleta, A.; Bellincampi, D. Cell wall traits as potential resources to improve resistance of durum wheat against Fusarium graminearum. BMC Plant Biol. 2015, 15, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.Y.; Hou, S.G.; Rodrigues, O.; Wang, P.; Luo, D.X.; Munemasa, S.; Lei, J.X.; Liu, J.; Ortiz-Morea, F.A.; Wang, X.; et al. Phytocytokine signalling reopens stomata in plant immunity and water loss. Nature 2022, 605, 332–339. [Google Scholar] [CrossRef]
- Makandar, R.; Nalam, V.J.; Lee, H.; Trick, H.N.; Dong, Y.; Shah, J. Salicylic acid regulates basal resistance to fusarium head blight in wheat. Mol. Plant Microbe Interact. 2012, 25, 431–439. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Cho, S.; Muehlbauer, G.J. Transcriptome analysis of a wheat near-isogenic line pair carrying fusarium head blight-resistant and susceptible alleles. Mol. Plant Microbe Interact. 2009, 22, 1366–1378. [Google Scholar] [CrossRef] [PubMed]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D.G. Hormone crosstalk in plant disease and defense: More than just jasmonate-salicylate antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Jin, X.; Jia, X.; Wang, H.; Cao, A.; Zhao, W.; Pei, H.; Xue, Z.; He, L.; Chen, Q.; et al. Transcriptome-based discovery of pathways and genes related to resistance against fusarium head blight in wheat landrace wangshuibai. BMC Genom. 2013, 14, 197. [Google Scholar] [CrossRef] [Green Version]
- Rawat, N.; Pumphrey, M.O.; Liu, S.; Zhang, X.F.; Tiwari, V.K.; Ando, K.; Trick, H.N.; Bockus, W.W.; Akhunov, E.; Anderson, J.A.A.; et al. Wheat Fhb1 encodes a chimeric lectin with agglutinin domains and a pore-forming toxin-like domain conferring resistance to fusarium head blight. Nat. Genet. 2016, 48, 1576–1580. [Google Scholar] [CrossRef]
- He, Y.; Zhang, X.; Zhang, Y.; Ahmad, D.; Wu, L.; Jiang, P.; Ma, H.X. Molecular characterization and expression of PFT, an FHB resistance gene at the Fhb1 QTL in wheat. Phtopathology 2018, 108, 730–736. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.H.; Hou, B.Q.; Su, P.S.; Wu, H.Y.; Wang, G.P.; Kong, L.R.; Ma, X.; Wang, H.W. Application of virus-induced gene silencing for identification of FHB resistant genes. J. Integr. Agric. 2019, 18, 2183–2192. [Google Scholar] [CrossRef]
- Makandar, R.; Nalam, V.; Chaturvedi, R.; Jeannotte, R.; Sparks, A.A.; Shah, J. Involvement of salicylate and jasmonates signaling pathways in Arabidopsis interaction with Fusarium graminearum. Mol. Plant Microbe Interact. 2010, 23, 861–870. [Google Scholar] [CrossRef] [Green Version]
- Gottwald, S.; Samans, B.; Lück, S.; Friedt, W. Jasmonate and ethylene dependent defence gene expression and suppression of fungal virulence factors: Two essential mechanisms of fusarium head blight resistance in wheat. BMC Genom. 2012, 13, 369. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Du, X.Y.; Liu, G.J.; Yang, Z.D.; Hou, W.Q.; Wang, H.W.; Feng, D.S.; Li, A.F.; Kong, L.R. Cloning and characterization of a novel UDP-glycosyltransferase gene induced by DON from wheat. J. Integr. Agric. 2015, 14, 830–838. [Google Scholar] [CrossRef] [Green Version]
- Zheng, T.; Hua, C.; Li, L.; Sun, Z.X.; Yuan, M.M.; Bai, G.H.; Humphreys, G.; Li, T. Integration of meta-QTL discovery with omics: Towards a molecular breeding platform for improving wheat resistance to fusarium head blight. Crop J. 2021, 9, 739–749. [Google Scholar] [CrossRef]
- Li, G.Q.; Jia, L.; Zhou, J.Y.; Fan, J.C.; Yan, H.S.; Shi, J.X.; Wang, X.; Fan, M.; Xue, S.L.; Cao, S.Y.; et al. Evaluation and precise mapping of QFhb.nau-2B conferring resistance against Fusarium infection and spread within spikes in wheat (Triticum aestivum L.). Mol. Breed. 2019, 39, 62. [Google Scholar] [CrossRef]
- Draeger, R.; Gosman, N.; Steed, A.; Chandler, E.; Thomsett, M.; Srinivasachary; Schondelmaier, J.; Buerstmayr, H.; Lemmens, M.; Schmolke, M.; et al. Identification of QTLs for resistance to fusarium head blight, DON accumulation and associated traits in the winter wheat variety Arina. Theor. Appl. 2007, 115, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Abate, Z.A.; Lu, H.; Musket, T.; Davis, G.L.; Mckendry, A.L. QTL Associated with fusarium head blight resistance in the soft red winter wheat ernie. Theor. Appl. Genet. 2007, 115, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Brown-Guedira, G.; Van, S.D.; Ohm, H.; Dong, Y.H.; McKendry, A.L. Novel QTL associated with the fusarium head blight resistance in truman soft red winter wheat. Theor. Appl. Genet. 2016, 207, 571–592. [Google Scholar] [CrossRef]
- Wilde, F.; Schon, C.C.; Korzun, V.; Ebmeyer, E.; Schmolke, M.; Hartl, L.; Miedaner, T. Marker-based introduction of three quantitative-trait loci conferring resistance to fusarium head blight into an independent elite winter wheat breeding population. Theor. Appl. Genet. 2008, 117, 29–35. [Google Scholar] [CrossRef]
- Buerstmayr, M.; Alimari, A.; Steiner, B.; Buerstmayr, H. Genetic mapping of QTL for resistance to fusarium head blight spread (type 2 resistance) in a Triticum Dicoccoides × Triticum Durum backcross-derived population. Theor. Appl. Genet. 2013, 126, 2825–2834. [Google Scholar] [CrossRef]
- Yabwalo, D.N.; Mergoum, M.; Berzonsky, W.A. Further characterization of the scab resistance of frontana spring wheat and the relationships between resistance mechanisms. Plant Breed. 2011, 130, 521–525. [Google Scholar] [CrossRef]
- Yang, J.; Bai, G.H.; Shaner, G.E. Novel quantitative trait loci (QTL) for fusarium head blight resistance in wheat cultivar Chokwang. Theor. Appl. Genet. 2005, 111, 1571–1579. [Google Scholar] [CrossRef]
- Somers, D.J.; Fedak, G.; Savard, M. Molecular mapping of novel genes controlling fusarium head blight resistance and deoxynivalenol accumulation in spring wheat. Genome 2003, 46, 555–564. [Google Scholar] [CrossRef]
- Kollers, S.; Rodemann, B.; Ling, J.; Korzun, V.; Ebmeyer, E.; Argillier, O.; Hinze, M.; Plieske, J.; Kulosa, D.; Ganal, M.W.; et al. Whole genome association mapping of fusarium head blight resistance in European winter wheat (Triticum aestivum L.). PLoS ONE 2013, 8, e57500. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.W.; Xu, D.A.; Cheng, S.H.; Gao, C.B.; Xia, X.C.; Hao, Y.F.; He, Z.H. Characterization of fusarium head blight resistance gene Fhb1 and its putative ancestor in chinese wheat germplasm. Acta Agron. Sin. 2018, 44, 473–482. [Google Scholar] [CrossRef]
- Wu, A.; Zhao, C.; Li, H.; Yao, M.J.; Liao, Y.C. Resistance response of two sets of differentiation wheat cultivars upon inoculation with representative Fusarium graminearum Isolates from China. Acta Phytopathol. Sin. 2006, 36, 285–288. [Google Scholar] [CrossRef]
- Jia, H.Y.; Zhou, J.Y.; Xue, S.L.; Li, G.Q.; Yan, H.S.; Ran, C.F.; Zhang, Y.D.; Shi, J.X.; Jia, L.; Wang, X.; et al. A journey to understand wheat fusarium head blight resistance in the chinese wheat landrace Wangshuibai. Crop J. 2018, 6, 48–59. [Google Scholar] [CrossRef]
- Zhu, X.W.; Zhong, S.B.; Chao, S.M.; Gu, Y.Q.; Kianian, S.F.; Elias, E.; Cai, X. Toward a better understanding of the genomic region harboring fusarium head blight resistance QTL Qfhs.ndsu-3AS in durum wheat. Theor. Appl. Genet. 2016, 129, 31–43. [Google Scholar] [CrossRef]
- Li, G.Q.; Zhou, J.Y.; Jia, H.Y.; Gao, Z.X.; Fan, M.; Luo, Y.J.; Zhao, P.T.; Xue, S.L.; Li, N.; Yuan, Y.; et al. Mutation of a histidine-rich calcium-binding -protein gene in wheat confers resistance to fusarium head blight. Nat. Genet. 2019, 51, 1106–1112. [Google Scholar] [CrossRef] [PubMed]
- Cuthbert, P.A.; Somers, D.J.; Thomas, J.; Cloutier, S.; Anita, B.B. Fine mapping Fhb1, a major gene controlling fusarium head blight resistance in bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 2006, 112, 1465–1472. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.Q.; Jin, S.J.; Zhang, D.D.; Bai, G.H. Development and validation of diagnostic markers for Fhb1 region, a major QTL for Fusarium head blight resistance in wheat. Theor. Appl. Genet. 2018, 131, 2371–2380. [Google Scholar] [CrossRef]
- Cuthbert, P.A.; Somers, D.J.; Anita, B.B. Mapping of Fhb2 on chromosome 6BS: A gene controlling fusarium head blight weld resistance in bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 2007, 114, 429–437. [Google Scholar] [CrossRef]
- Xue, S.L.; Li, G.Q.; Jia, H.Y.; Xu, F.; Lin, F.; Tang, M.Z.; Wang, Y.; An, X.; Xu, H.B.; Zhang, L.X.; et al. Fine mapping Fhb4, a major QTL conditioning resistance to fusarium infection in bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 2010, 121, 147–156. [Google Scholar] [CrossRef]
- Xue, S.L.; Xu, F.; Tang, M.Z.; Zhou, Y.; Li, G.Q.; An, X.; Lin, F.; Xu, H.B.; Jia, H.Y.; Zhang, L.X.; et al. Precise mapping Fhb5, a major QTL conditioning resistance to fusarium infection in bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 2011, 123, 1055–1063. [Google Scholar] [CrossRef]
- Wang, H.W.; Sun, S.L.; Ge, W.Y.; Zhao, L.F.; Hou, B.B.; Wang, K.; Lyu, Z.F.; Chen, L.Y.; Xu, S.S.; Guo, J.; et al. Horizontal gene transfer of Fhb7 from fungus underlies fusarium head blight resistance in wheat. Science 2020, 368, eaba5435. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.Q.; Xue, S.L.; Lin, F.; Yang, S.H.; Li, G.Q.; Tang, M.Z.; Kong, Z.X.; Cao, Y.; Zhao, D.M.; Jia, H.Y.; et al. Mapping and validation of scab resistance QTLs in the Nanda2419× Wangshuibai population. Ceral Res. Commun. 2008, 36, 245–251. [Google Scholar] [CrossRef]
- Lin, F.; Xue, S.L.; Zhang, Z.Z.; Zhang, C.Q.; Kong, Z.X.; Yao, G.Q.; Tian, D.G.; Zhu, H.L.; Li, C.J.; Cao, Y.; et al. Mapping QTL Associated with resistance to fusarium head blight in the Nanda2419 x Wangshuibai population. I: Type II Resistance. Theor. Appl. Genet. 2004, 109, 1504–1511. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, W.; Steiner, B.; Ametz, C.; Siegwart, G.; Wiesenberger, G.; Berthiller, F.; Lemmens, M.; Jia, H.Y.; Adam, G.; Muehlbauer, G.J.; et al. Transcriptomic characterization of two major Fusarium resistance quantitative trait loci (QTLs), Fhb1 and qfhs.ifa-5A, identifies novel candidate genes. Mol. Plant Pathol. 2013, 14, 772–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, J.; Wang, Z.; Du, Z.; Che, M.; Zhang, Y.; Quan, W.; Wang, Y.; Jiang, X.; Zhang, Z. Detection and validation of a novel major QTL for resistance to fusarium head blight from Triticum Aestivum in the terminal region of chromosome 7DL. Theor. Appl. Genet. 2019, 132, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Soni, N.; Hegde, N.; Dhariwal, A.; Kushalappa, A.C. Role of laccase gene in wheat NILs differing at QTL-Fhb1 for resistance against fusarium head blight. Plant Sci. 2020, 298, 110574. [Google Scholar] [CrossRef]
- Soni, N.; Altartouri, B.; Hegde, N.; Duggabathi, R.; Farhad, N.F.; Kushalappa, A.C. TaNAC032 Transcription factor regulates ligninbiosynthetic genes to combat fusarium head blight in wheat. Plant Sci. 2021, 304, 110820. [Google Scholar] [CrossRef]
- He, Y.; Ahmad, D.; Zhang, X.; Zhang, Y.; Wu, L.; Jiang, P.; Ma, H.X. Genome-wide analysis of family-1UDP glycosyl transferases (UGT) and identification of UGT genes for FHB resistance in wheat (Triticumaestivum L.). BMC Plant Biol. 2018, 18, 67. [Google Scholar] [CrossRef]
- Zhao, L.F.; Ma, X.; Su, P.S.; Ge, W.Y.; Wu, H.Y.; Guo, X.X.; Li, A.F.; Wang, H.W.; Kong, L.R. Cloning and characterization of a specific UDP-glycosyltransferase gene induced by DON and Fusarium graminearum. Plant Cell Rep. 2018, 37, 641–652. [Google Scholar] [CrossRef]
- Xing, L.P.; Gao, L.; Chen, Q.G.; Pei, H.Y.; Di, Z.C.; Xiao, J.; Wang, H.Y.; Ma, L.L.; Chen, P.D.; Cao, A.Z.; et al. Over-expressing a UDP-glucosyltransferase Gene (Ta-UGT3) enhances fusarium head blight resistance of wheat. Plant Growth Regul. 2018, 84, 561–571. [Google Scholar] [CrossRef]
- Gatti, M.; Choulet, F.; Macadré, C.; Guérard, F.; Seng, J.M.; Langin, T.; Dufresne, M. Molecular cloning and functional characterization of a wheat UDP-glucosyltransferase involved in resistance to fusarium head blight and to mycotoxin accumulation. Front. Plant Sci. 2018, 9, 1853. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Wu, L.; Liu, X.; Jiang, P.; Yu, L.X.; Qiu, J.B.; Wang, G.; Zhang, X.; Ma, H.X. TaUGT6, a Novel UDP-Glycosyl Transferase Gene Enhances the Resistance to FHB and DON Accumulation in Wheat. Plant Growth Regul. 2020, 11, 574775. [Google Scholar] [CrossRef]
- Hu, C.H.; Wang, P.Q.; Zhang, P.P.; Nie, X.M.; Li, B.B.; Tai, L.; Liu, W.T.; Li, W.Q.; Chen, K.M. NADPH oxidases: The vital performers and center hubs during plant growth and signaling. Cells 2020, 9, 437. [Google Scholar] [CrossRef] [Green Version]
- Kage, U.; Yogendra, K.N.; Kushalappa, A.C. TaWRKY70 Transcription factor in wheat QTL-2DL regulates downstream metabolite biosynthetic genes to resist Fusarium graminearum infection spread within spike. Sci. Rep. 2017, 7, 42596. [Google Scholar] [CrossRef] [Green Version]
- Zeiss, D.R.; Piater, L.A.; Dubery, L.A. Hydroxycinnamate amides: Intriguing conjugates of plant protective metabolites. Trends Plant Sci. 2021, 26, 184–195. [Google Scholar] [CrossRef]
- Kage, U.; Karre, S.; Kushalappa, A.C.; McCartney, C. Identification and characterization of a fusarium head blight resistance gene TaACT in wheat QTL-2DL. Plant Biotechnol. J. 2017, 15, 447–457. [Google Scholar] [CrossRef]
- Han, X.Y.; Zhang, L.; Zhao, L.F.; Xue, P.Y.; Qi, T.; Zhang, C.L.; Yuan, H.B.; Zhou, L.X.; Wang, D.W.; Qiu, J.L.; et al. SnRK1 Phosphorylates and destabilizes WRKY3 to enhance barley immunity to powdery mildew. Plant Commut. 2020, 1, 14. [Google Scholar] [CrossRef] [PubMed]
- Wood, A.K.M.; Panwar, V.; Grimwade-Mann, M.; Ashfield, T.; Hammond-Kosack, K.E.; Kanyuka, K. The vesicular trafficking system component MIN7 is required for minimizing Fusarium graminearum infection. J. Exp. Bot. 2021, 72, 5010–5023. [Google Scholar] [CrossRef]
- Perochon, A.; Kahla, A.; Monika, V.; Jia, J.; Malla, K.B.; Craze, M.; Wallington, E.; Doohan, F.M. A wheat NAC interacts with an orphan protein and enhances resistance to fusarium head blight disease. Plant Biotechnol. 2019, 17, 1892–1904. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.R.; Luo, Y.J.; Zhao, P.T.; Jia, H.Y.; Ma, Z.Q. Overexpression of TaJRL53 enhances the fusariun head blight resistance in wheat. Acta Agron. Sin. 2021, 47, 19–29. [Google Scholar] [CrossRef]
- Song, P.W.; Zhang, L.F.; Wu, L.L.; Hu, H.Y.; Liu, Q.L.; Li, D.X.; Hu, P.; Zhou, F.; Bu, R.F.; Wei, Q.C.; et al. A ricin B-like lectin protein physically interacts with TaPFT and is involved in resistance to fusarium head blight in wheat. Phytopathology 2021, 111, 2309–2316. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Song, P.W.; Xu, J.; Wei, Q.C.; Tao, Y.; Ren, Y.M.; Yu, Y.A.; Li, D.X.; Hu, H.Y.; Li, C.W. Genome-wide analysis of serine hydroxymethyltransferase genes in triticeae species reveals that TaSHMT3A-1 regulates fusarium head blight resistance in wheat. Front. Plant Sci. 2022, 13, 847087. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Hei, R.; Yang, Y.; Zhang, S.J.; Wang, Q.H.; Wang, W.; Zhang, Q.; Yan, M.; Zhu, G.R.; Huang, P.P.; et al. An orphan protein of Fusarium graminearum modulates host immunity by mediating proteasomal degradation of TaSnRK1α. Nat. Commun. 2020, 11, 4382. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.Q.; Mu, J.J.; Su, P.S.; Wu, H.Y.; Yu, G.H.; Wang, G.P.; Wang, L.; Ma, X.; Li, A.F.; Wang, H.W.; et al. Multi-functional roles of TaSSI2 involved in fusarium head blight and powdery mildew resistance and drought tolerance. J. Integr. Agric. 2018, 17, 368–380. [Google Scholar] [CrossRef]
- Bahrini, I.; Ogawa, T.; Kobayashi, F.; Kawahigashi, H.; Handa, H. Overexpression of the pathogen-inducible wheat TaWRKY45 gene confers disease resistance to multiple fungi in transgenic wheat plants. Breed. Sci. 2011, 61, 319–326. [Google Scholar] [CrossRef] [Green Version]
- Bahrini, I.; Sugisawa, M.; Kikuchi, R.; Ogawa, T.; Kawahigashi, H.; Ban, T.; Handa, H. Characterization of a wheat transcription factor, TaWRKY45, and its effect on fusarium head blight resistance in transgenic wheat plants. Breed. Sci. 2011, 61, 121–129. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.L.; Wu, T.C.; Xu, G.B.; Qi, H.J.; Zhu, X.L.; Zhang, Z.Y. TaWAK2A-800, a wall-associated kinase, participates positively in resistance to fusarium head blight and sharp eyespot in wheat. Int. J. Mol. Sci. 2021, 22, 11493. [Google Scholar] [CrossRef]
- Paudel, B.; Zhuang, Y.H.; Galla, A.; Dahal, S.; Qiu, Y.J.; Ma, A.; Raihan, T.; Yen, Y. WFhb1-1 plays an important role in resistance against fusarium head blight in wheat. Exp. Bot. 2020, 10, 7794. [Google Scholar] [CrossRef]
- Karre, S.; Kumar, A.; Dhokane, D.; Kushalappa, A.C. Metabolo-transcriptome profiling of barley reveals induction of chitin elicitor receptor kinase gene (HvCERK1) conferring resistance against Fusarium graminearum. Plant Mol. Biol. 2017, 93, 247–267. [Google Scholar] [CrossRef]
- Karre, S.; Kumar, A.; Yogendra, K.; Kage, U.; Kushalappa, A.; Charron, J.B. HvWRKY23 Regulates flavonoid glycoside and hydroxycinnamic acid amide biosynthetic genes in barley to combat fusarium head blight. Plant Mol. Biol. 2019, 100, 591–605. [Google Scholar] [CrossRef]
- Perochon, A.; Jia, J.; Kahla, A.; Arunachalam, C.; Scofield, S.R.; Bowden, S.; Wallington, E.; Doohan, F.M. TaFROG encodes a Pooideae orphan protein that interacts with SnRK1 and enhances resistance to the mycotoxigenic fungus Fusarium graminearum. Plant Physiol. 2015, 169, 2895–2906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perochona, A.; Várya, Z.; Mallaa, K.B.; Halfordb, N.G.; Paulb, M.J.; Doohan, F.M. The wheat SnRK1α family and its contribution to fusarium toxin tolerance. Plant Sci. 2019, 288, 110217. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.T.; Lu, Y.; Li, J.J.; Zhang, X.R.; Hu, F.F.; Zhao, Y.; Zhou, D.X. SnRK1 Stimulates the histone H3K27me3 demethylase JMJ705 to regulate a transcriptional switch to control energy homeostasis. Plant Cell 2021, 33, 3721–3742. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.H.; Wei, X.Y.; Yuan, B.; Yao, L.B.; Ma, T.T.; Zhang, P.P.; Wang, X.; Wang, P.Q.; Liu, W.T.; Li, W.Q.; et al. Genome-wide identification and functional analysis of NADPH oxidase family genes in wheat during development and environmental stress responses. Front. Plant Sci. 2018, 9, 906. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Li, M.; Yu, L.; Zhou, Z.; Liang, X.; Liu, Z.; Cai, G.; Gao, L.; Zhang, X.; Wang, Y.; et al. The FLS2-associated kinase BIK1 directly phosphorylates the NADPH Oxidase RbohD to control plant immunity. Cell Host Microbe 2014, 15, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Akamatsu, A.; Wong, H.L.; Masayuki, F.; Jun, O.; Keita, N.; Uno, K.; Imai, K.; Umemura, K.; Kawasaki, T.; Kawano, Y.; et al. An OsCEBiP/OsCERK1-OsRacGEF1-OsRac1 module is an essential rarly component of rchitin-induced rice immunity. Cell Host Microbe 2013, 13, 465–476. [Google Scholar] [CrossRef] [Green Version]
- Ding, L.N.; Xu, H.B.; Yi, H.Y.; Yang, L.M.; Kong, Z.X.; Zhang, L.X.; Xue, S.L.; Jia, H.Y.; Ma, Z.Q. Resistance to hemi-biotrophic F. graminearum infection is associated with coordinated and ordered expression of diverse defense signaling pathways. PLoS ONE 2011, 6, e19008. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Liang, Y.; Tanaka, K.; Nguyen, C.T.; Jedrzejczak, R.P.; Joachimiak, A.; Stacey, G. The kinase LYK5 is a major chitin receptor in Arabidopsis and forms a chitin-induced complex with related kinase CERK1. Elife 2014, 3, e03766. [Google Scholar] [CrossRef]
- Wan, J.; Tanaka, K.; Zhang, X.C.; Son, G.H.; Brechenmacher, L.; Nguyen, T.H.; Stacey, G. LYK4, a lysin motif receptor-like kinase, is important for chitin signaling and plant innate immunity in Arabidopsis. Plant Physiol. 2012, 160, 396–406. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Wu, Y.; Yang, F.; Zhang, Y.; Chen, S.; Xie, Q.; Tian, X.; Zhou, J.M. BIK1 interacts with PEPRs to mediate ethylene-induced immunity. Proc. Natl. Acad. Sci. USA 2013, 110, 6205–6210. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.L.; Grubb, L.E.; Wang, J.Y.; Liang, X.X.; Li, L.; Gao, C.L.; Ma, M.M.; Feng, F.; Li, M.; Li, L.; et al. A Regulatory module controlling homeostasis of a plant immune kinase. Mol. Cell 2018, 69, 493–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ao, Y.; Li, Z.; Feng, D.; Xiong, F.; Liu, J.; Li, J.F.; Wang, M.; Wang, J.; Liu, B.; Wang, H.B. OsCERK1 and OsRLCK176 play important roles in peptidoglycan and chitin signaling in rice innate immunity. Plant J. 2014, 80, 1072–1084. [Google Scholar] [CrossRef] [PubMed]
- Takeo, S.; Takuto, N.; Daisuke, T.; Yoshitake, D.; Naoko, I.; Yoko, N.; Eiichi, M.; Kazunori, O.; Hisakazu, Y.; Hanae, K.; et al. Two LysM receptor molecules, CEBiP and OsCERK1, cooperatively regulate chitin elicitor signaling in rice. Plant J. 2010, 64, 204–214. [Google Scholar] [CrossRef] [Green Version]
- Masahiro, H.; Rita, B.; Roberta, M.; Alba, S.; Miyu, K.; Yoshitake, D.; Sakiko, A.; Flavia, S.; Alessia, R.; Ken, T.; et al. Chitin-induced activation of immune signaling by the rice receptor CEBiP relies on a unique sandwich-type dimerization. Proc. Natl. Acad. Sci. USA 2013, 111, e404–e413. [Google Scholar] [CrossRef] [Green Version]
- Boisson-Dernier, A.; Lituiev, D.S.; Nestorova, A.; Franck, C.M.; Thirugnanarajah, S.; Grossniklaus, U. ANXUR Receptor-like kinases coordinate cell wall integrity with growth at the pollen tube tip via NADPH oxidases. PLoS Biol. 2013, 11, e1001719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashima, A.; Chen, L.; Thao, N.P.; Fujiwara, M.; Wong, H.L.; Kuwano, M.; Umemura, K.; Shirasu, K.; Kawasaki, T.; Shimamoto, K. RACK1 functions in rice innate immunity by interacting with the Rac1 immune complex. Plant Cell 2008, 20, 2265–2279. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, T.; Yamada, K.; Yoshimura, S.; Yamaguchi, K. Chitin receptor-mediated activation of MAP kinases and ROS production in rice and Arabidopsis. Plant Signal Behav. 2017, 12, e1361076. [Google Scholar] [CrossRef]
- Köster, P.; DeFalco, T.A.; Zipfel, C. Ca2+ signals in plant immunity. EMBO J. 2022, 41, e110741. [Google Scholar] [CrossRef]
- Shinya, T.; Motoyama, N.; Ikeda, A.; Wada, M.; Kamiya, K.; Hayafune, M.; Kaku, H.; Shibuya, N. Functional characterization of CEBiP and CERK1 homologs in Arabidopsis and rice reveals the presence of different chitin receptor systems in plants. Plant Cell Physiol. 2012, 53, 1696–1706. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Fan, X.; He, M.Y.; Hu, Z.Y.; Tang, C.L.; Zhang, S.; Lin, D.X.; Gan, P.F.; Wang, J.F.; Huang, X.L.; et al. Transcriptional repression of TaNOX10 by TaWRKY19 compromises ROS generation and enhances wheat susceptibility to stripe rust. Plant Cell 2022, 34, 784–1803. [Google Scholar] [CrossRef]
- Yamauchi, T.; Yoshioka, M.; Fukazawa, A.; Mori, H.; Nishizawa, N.K.; Tsutsumi, N.; Yoshioka, H.; Nakazono, M. An NADPH oxidase RBOH functions in rice roots during lysigenous aerenchyma formation under oxygen-deficient conditions. Plant Cell 2017, 29, 775–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potocký, M.; Pejchar, P.; Gutkowska, M.; Jiménez-Quesada, M.J.; Potocká, A.; de Dios, A.J.; Kost, B.; Zárský, V. NADPH oxidase activity in pollen tubes is affected by calcium ions, signaling phospholipids and Rac/Rop GTPases. J. Plant Physiol. 2012, 169, 1654–1663. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Ohura, I.; Kawakita, K.; Yokota, N.; Fujiwara, M.; Shimamoto, K.; Doke, N.; Yoshioka, H. Calcium-dependent protein kinases regulate the production of reactive oxygen species by potato NADPH oxidase. Plant Cell 2007, 19, 1065–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubiella, U.; Seybold, H.; Durian, G.; Komander, E.; Lassig, R.; Witte, C.P.; Schulze, W.X.; Romeis, T. Calcium-dependent protein kinase/NADPH oxidase activation circuit is required for rapid defense signal propagation. Proc. Natl. Acad. Sci. USA 2013, 110, 8744–8749. [Google Scholar] [CrossRef] [Green Version]
- Majumdar, A.; Kar, R.K. Congruence between PM H+-ATPase and NADPH oxidase during root growth: A necessary probability. Protoplasma 2019, 255, 1129–1137. [Google Scholar] [CrossRef]
- Hu, C.H.; Zeng, Q.D.; Li, T.; Li, B.B.; Zhang, P.P.; Nie, X.M.; Wang, P.Q.; Liu, W.T.; Li, W.Q.; Kang, Z.S.; et al. Interaction between TaNOX7 and TaCDPK13 contributes to plant fertility and drought tolerance by regulating ROS production. J. Agric. Food Chem. 2020, 68, 7333–7347. [Google Scholar] [CrossRef]
- Gupta, K.; Sengupta, A.; Chakraborty, M.; Gupta, B. Hydrogen peroxide and polyamines act as double-edged swords in plant abiotic stress responses. Front. Plant Sci. 2016, 7, 1343. [Google Scholar] [CrossRef] [Green Version]
- Xu, G. Study on the Biocontrol of Wheat Scab (Fusarium graminearum) with Bacillus subtilis BS3-1 Strain. Master’s thesis, Sichuan Agricultural University, Sichuan, China, 2003. [Google Scholar]
- Lu, W.; Zhao, Q.M.; Chen, Y.L.; Han, M.M.; Cai, J.; Chen, Y.H. Distribution of chitinase in Bacillus thuringriensis and screening of antifungal strains against Fusarium graminearum. Acta Scentiarum Nat. Univ. Nankaiensis 2007, 40, 97–101. [Google Scholar]
- Li, Z.H.; Xiang, J.J.; Chen, J.H.; Ge, C.F.; Ge, S.R. Isolation and identification of actinomycete against Fusarium graminearum Schw. J. Triticeae Crops 2007, 27, 149–152. [Google Scholar]
- Zhang, J.; Chen, J.; Hu, L.F.; Jia, R.M.; Ma, Q.; Tang, J.J.; Wang, Y. Antagonistic action of Streptomyces pratensis S10 on Fusarium graminearum and its complete genome sequence. Environ. Microbiol. 2020, 23, 1925–1940. [Google Scholar] [CrossRef]
- Han, C.Y.; Yu, Z.Y.; Zhang, Y.T.; Wang, Z.Y.; Zhao, J.W.; Huang, S.X.; Ma, Z.H.; Wen, Z.Y.; Liu, C.X.; Xiang, W.S. Discovery of frenolicin B as potential agrochemical fungicide for controlling fusarium head blight on wheat. J. Agric. Food Chem. 2021, 69, 2108–2117. [Google Scholar] [CrossRef]
- Cantoro, R.; Palazzini, J.M.; Yerkovich, N.; Miralles, D.J.; Chulze, S.N. Bacillus Velezensis RC 218 as a biocontrol agent against Fusarium graminearum: Effect on penetration, growth and TRI5 expression in wheat spikes. BioControl 2021, 66, 259–270. [Google Scholar] [CrossRef]
- Hao, Q.Y.; Albaghdady, D.M.D.; Xiao, Y.N.; Xiao, X.Q.; Mo, C.; Tian, T.; Wang, G.F. Endophytic Metarhizium Anisopliae is a potential biocontrol agent against wheat fusarium head blight caused by Fusarium graminearum. J. Plant Pathol. 2021, 103, 875–885. [Google Scholar] [CrossRef]
- Xu, S.; Wang, Y.; Hu, J.Q.; Chen, X.R.; Qiu, Y.F.; Shi, J.R.; Wang, G.; Xu, J.H. Isolation and characterization of Bacillus amyloliquefaciens Mq01, a bifunctional biocontrol bacterium with antagonistic activity against Fusarium graminearum and biodegradation capacity of zearalenone. Food Control 2021, 130, 108259. [Google Scholar] [CrossRef]
- Xu, S.D.; Liu, Y.X.; Gernava, T.; Wang, H.K.; Zhou, Y.Q.; Xia, T.; Cao, S.G.; Berg, G.; Shen, X.X.; Wen, Z.Y.; et al. Fusarium fruiting body microbiome member Pantoea agglomerans inhibits fungal pathogenesis by targeting lipid rafts. Nat. Microbiol. 2022, 7, 1131. [Google Scholar] [CrossRef]
- Song, Y.Y.; Lin, Y.; Lu, H.G.; Luo, Z.X. Sensitivity of Fusarium graminearum from Hubei province to carbendazim, tebuconazole and prochloraz. Chin. J. Pestic. Sci. 2016, 18, 724–728. [Google Scholar] [CrossRef]
- Dai, D.; Jia, X.; Wu, D.; Hou, Y.P.; Chen, Z.J.; Zhou, M.G. Analysis of diffusion path of carbendazim-resistance population of fusarium head blight―based on Fusarium species, mycotoxin chemotype and resistance timing. Chin. J. Pestic. Sci. 2013, 15, 279–285. [Google Scholar] [CrossRef]
- Fu, J.; Zhu, J.Q.; Qing, J.; Li, X.Y.; Zhang, G.H.; Sun, W.Z.; Wu, H.Q.; Zhou, J.; Zhu, X.L.; Zhang, A.C. Resistance monitoring of Fusarium asiaticum to carbendazim in Jiangsu state farms and the efficacy of alternative fungicides to wheat scab. Plant Prot. Sci. 2017, 43, 196–201. [Google Scholar] [CrossRef]
- Rajeshkumar, S.; Malarkodi, C.; Vanaja, M.; Annadurai, G. Anticancer and enhanced antimicrobial activity of biosynthesizd silver nanoparticles against clinical pathogens. J. Mol. Struct. 2016, 1116, 165–173. [Google Scholar] [CrossRef]
- Takemoto, J.Y.; Wegulo, S.N.; Yuen, G.Y.; Stevens, J.A.; Jochum, C.C.; Chang, C.W.T.; Kawasaki, Y.K.; Miller, G.W. Suppression of wheat fusarium head blight by novel amphiphilic aminoglycoside fungicide K20. Fungal Biol. 2018, 122, 465–470. [Google Scholar] [CrossRef]
- Duan, Y.B.; Xiao, X.M.; Li, T.; Chen, W.W.; Wang, J.X.; Fraaije, B.A.; Zhou, M.G. Impact of epoxiconazole on fusarium head blight control, grain yield and deoxynivalenol accumulation in wheat. Pestic. Biochem. Phys. 2018, 152, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.Q.; ZHang, Y.H.; Hua, C.; Li, L.; Sun, Z.X.; Li, T. Effect of vitamin E on the accumulation of deoxynivalenol in wheat. J. Triticeae Crops 2020, 1142–1147. [Google Scholar] [CrossRef]
- Ma, D.C.; Wang, G.X.; Zhu, J.M.; Mu, W.; Dou, D.L.; Feng, L. Green leaf volatile trans-2-hexenal inhibits the growth of Fusarium graminearum by inducing membrane damage, ROS accumulation, and cell dysfunction. J. Agric. Food Chem. 2022, 70, 5646–5657. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| QTL Name | Flanking Marker | on Chr. * | Resistance Type | Representative | Reference |
|---|---|---|---|---|---|
| Fhb1Qfhs.ndsu-3AS | Xwgc501–Xwgc510 | Chr. 3AS | Type II | Triticum dicoccoides | [88] |
| Fhb1(Qfhs.njau-3B) | Xmag8937–Xmag9404 | Chr. 3BS | Type II | Sumai 3 | [89] |
| Fhb1(Qfhs.ndsu-3BS) | XGWM533–XGWM493 | Chr. 3BS | Type II | Sumai 3 | [90] |
| Fhb1(TaHRC) | TaHRC-GSM/TaHRC-Kasp | Chr. 3BS | Type II | Sumai 3 | [91] |
| Fhb2 | Xwmc398–Xgwm644 | Chr. 6BS | Type II | Sumai 3 | [92] |
| Fhb3 | BE586744-STS–BE586111-STS | Chr. 7ALr#1S | Type II | Sumai 3 | [50] |
| Fhb4(Qfhi.nau-4B) | Xhbg226–Xgwm149 | Chr. 4B | Type I | Wangshuibai | [93] |
| Fhb5(Qfhi.nau-5A) | Xbarc56–Xbarc100 | Chr. 5A | Type I | Mianyang 99-323 | [94] |
| Fhb6 | BF202643/HaeIII-BE591682/HaeIII | Chr. 1E | Type II | Sumai 3 | [51] |
| Fhb7 | XsdauK86–XsdauK88 | Chr. 7E/7D | Type V | Elytrigia elongata | [95] |
| Qfhs.nau-2A | Xwmc474–Xsm1021 | Chr. 2A | Type II | Nanda 2419 | [96] |
| Qfhi.nau-2B | Xwmc499–Xmag1729 | Chr. 2B | Type I | Nanda 2419 | [26] |
| Qfhs.nau-2B | Xmag1811.1–Xmag3080 | Chr. 2B | Type II | WSB | [96] |
| QFhb.nau-2B | Xwgrb1561–Xwgrb1420 | Chr. 2B | Type I/Type II | Nanda 2419 | [90] |
| Qfhi.nau-3A | Xwmc169–Xgwm162 | Chr. 3A | Type I | WSB | [26] |
| QFhb-hnau.3BS.1 | gwm533–stm748tcac | Chr. 3BS | Type I/Type II | Landrace N553 | [25] |
| Qfhs.nau-3B | Xgwm389–Xbarc102 | Chr. 3B | Type II | WSB | [96] |
| Qfhi.nau-4A | Xwmc161–Xmag3886 | Chr. 4A | Type I | Nanda 2419 | [26] |
| Qfhi.nau-4B | Xgwm495–Xgwm149 | Chr. 4B | Type I | WSB | [97] |
| Qfhs.ifa-5A | barc186–wmc805 | Chr. 5A | Type I/Type II | Chinese spring | [98] |
| QFhb-5A | Xgwm304–Xgwm415 | Chr. 5AS | Type II | Yangmai 158 | [55] |
| Qfhi.nau-5A | Xbarc180–Xgwm186 | Chr. 5A | Type I | WSB | [97] |
| Qfhs.nau-6B | Xwmc398–Xmag359 | Chr. 6B | Type II | WSB | [96] |
| Qfhi.nau-2D | Xwmc181–Xaf12 | Chr. 2D | Type I | WSB | [26] |
| QFhb-hnau.2DL | AX-110955068–AX-109419238 | Chr. 2DL | Type I/Type II | Yangmai 13 | [25] |
| QFhb.cau-7DL | gwm428 | Chr. 7DL | Type II/Type I | Sumai 3 | [99] |
| Nomenclature | Protein | Gene ID | Location on Chr. */ Flanking Markers | Function |
|---|---|---|---|---|
| FHB7-GST | GST: Glutathione transferase | Tel7E01T1020600.1 | Chr. 7E/7D (XsdauK86, XsdauK88) | A FHB-resistance gene, Fhb7, from Thinopyrum elongatum, introgressions artificially in wheat, confers resistance to FHB without yield penalty [95]. |
| LTP | Lipid transfer protein | Ta.1282.4.S1_at | Chr. 5A In the interval of Qfhs.ifa-5A. | LTPs might confer plant type I resistance against initial fungal penetration of Fg and also contribute to toxin resistance [98]. |
| MIN7 | ARF-GEF protein | TraesCS2A02G202900 TraesCS2B02G230000 TraesCS2D02G212800 | Chr. 2 | TaMIN7 plays positive role, involved in response to Fg by disturbing vesicular trafficking in cells [112]. |
| Qfhs.ifa-5A (Fhb1) | Lipid transfer protein | GenBank: FN564434 | Chr. 3BS | Confers type II resistance [98]. |
| Qfhs.njau-3B | Histidine-rich calcium-binding protein | GenBank: KX022627.1–KX022633.1, MK397611–MK397761 (His: Xmag8937) | Chr. 3BS Xmag8937 and Xmag9404 | Qfhs.njau-3B is a candidate gene on Fhb1, encodes a histidine-rich calcium-binding protein, mutation of it in wheat confers resistance to FHB [89]. |
| TaAOS | Oxidized propadiene synthase | TraesCS4A02G061900 TraesCS4D02G238800 TraesCS4B02G237600 | Chr. 4A/4D/4B | TaAOS involved in JA signaling pathway to enhance plant resistance to FHB. TaAOS-silenced strains exhibit high susceptibility to FHB [70]. |
| TaACT | Agmatine coumaroyl transferase | GenBank: KT962210 | Chr. 2DL (FHB QTL-2DL) (WMC245-GWM608) | TaACT is an important gene in wheat FHB QTL-2DL, conferring type II resistance to Fg by limiting the spread of pathogen from the initial point of infection [110]. |
| TaFROG | Orphan protein | GenBank: KR611570 | Chr. 4A (CM82036) | TaFROG binds to the protein TaSnRK1α to prevent TaSnRK1α from being degraded by ubiquitination, thereby increasing the resistance of the plant to FHB [113]. |
| TaHRC (TaHRC-S) (TaHRC-R) (Fhb1) | Histidine-rich calcium-binding protein | GenBank: CBH32655.1 GenBank: MK450309 GenBank: MK450312 | Chr. 3BS in the interval of QTL-Fhb1 (syn Qfhs.ndsu-3BS; Gwm533,Gwm493) | TaHRC, a key factor of fhb1-mediated FHB resistance, encodes a nuclear protein (histidine-rich calcium-binding protein) with FHB susceptibility. Mutating TaHRC in plants leads to increased resistance to FHB [52]. |
| TaJRL53 | Jacalin-related lectins protein | TraesCS4A02G430200.1 | Chr. 4AL | TaJRL53 enhanced FHB resistance in wheat through regulating ROS synthesis pathway and JA signal transduction pathways [114]. |
| TaLAC4 (Fhb1) | Laccase | TraesCS3B02G392700.1 | At QTL-Fhb1 on Chr. 3BS (syn Qfhs.ndsu-3BS; WMC245,GWM608) | TaLAC4, a wheat laccase, can increase resistance of plants to FHB by promoting the synthesis of lignin in the secondary cell wall to thicken the cell wall and confers type II resistance [100]. |
| TaNACL-D1 | N-Acetylcysteine-like transcription factors | TraesCS5D02G111300 (GenBank: MG701911) | Ch. 5D | TaNACL-D1 interacts with the orphan protein TaFROG to improve the resistance of the plant to FHB [107]. |
| TaNAC032 (Fhb1) | Lignin-biosynthetic | GenBank: MT512636 | At QTL-Fhb1 (syn Qfhs.ndsu-3BS) XSTS3B-80 and XSTS3B-142 | Promotes transcription of lignin genes associated with resistance-related metabolite biosynthesis [101]. |
| TaPFT | Pore-forming toxin-like | GenBank: KX907434. | Chr. 3BS At QTL-Fhb1 (syn Qfhs.ndsu-3BS) | PFT, encoding a perforatoxin analog, is from Fhb1 on the genome of Sumai 3 and shows significant resistance to FHB [68,69]. |
| TaRBL | Ricin B-like lectin protein | TraesCS3A02G078200.1 TraesCS3B02G093100.2 TraesCS3D02G078700.1 | Chr. 3 | TaRBL interacts with TaPFT and is involved in resistance to FHB in wheat [115]. |
| TaSHMT3 | Serine hydroxy methyltransferase | TraesCS3A02G385600 | Chr. 3A | Silencing of TaSHMT3A-1 compromises Fusarium head blight resistance in wheat [116]. |
| TaSnRK1α | The alpha subunit of sucrose non- fermentation-related kinase 1 (SnRK1) | GenBank: KR611568 | Chr. 4A (CM82036) | TaSnRK1α interacts with TaFROG to prevent degradation by ubiquitination and improves the resistance of the plant to FHB [113,117]. |
| TaSSI2-2AL -2BL -2DL | Stearoyl-acyl carrier protein fatty acid desaturase | AA0283540, AA0388780 and AK332689 | Chr. 2 | TaSSI2 probably regulated FHB resistance by depressing the SA signaling pathway in wheat [118]. |
| TaUGT3 (Fhb1) | UDP-glycosyltransferase | GenBank: FJ236328 | Chr. 3BS | Ta-UGT 3 was found to enhance host tolerance against deoxynivalenol (DON) in Arabidopsis [104]. |
| TaUGT5 | UDP-glycosyltransferase | Ta_iwgsc_2bs_vl_5195782 | Chr. 2B | TaUGT5 can reduce the proliferation and destruction of Fg and enhance the ability of FHB resistance in wheat [102]. |
| TaUGT6 | Glycosyltransferases | TraesCS5B02G436300 | Chr. 2B | The protein TaUGT6 can transform the toxin DON (deoxynivalenol) into non-toxic D3G (D3-glucoside); O\overexpression of TaUGT6 in wheat enhanced the resistance of plant to Fg [106]. |
| TaUGT12887 | Glycosyltransferases | TraesCS5B02G148300 GenBank accession JX624788 | In the interval of QTL Fhb1 | Confers plant weak resistance against DON [98]. |
| TaWRKY45 | WRKY-like transcription factor | EMBL/GenBank accession numbers AB603888, AB603889; AB603890 | Chr.2 | Overexpression of the TaWRKY45 transgene conferred an enhanced resistance against Fg in wheat [119,120]. |
| TaWRKY70 | WRKY-like transcription factor | TraesCS2D02G489700 | In the interval of QTL-2DL | TaWRKY70 silencing lines not only increased fungal biomass but also decreased expressions of downstream resistance genes TaACT, TaDGK, and TaGLI1, along with decreased abundances of RRI metabolites biosynthesized by them [108]. |
| TaWAK2A-800 | Wall-associated kinase | TraesCS2A02G071800.1 | Chr.2A | TaWAK2A-800 participates positively in the resistance responses to Fg, possibly through regulating the chitin-triggering immune pathway marker genes, TaCERK1, TaRLCK1B, and TaMPK3 in wheat [121]. |
| UDP-UGT | Uridine diphosphate (UDP)- glucosyltransferase (UGT) | Traes_2BS_14CA35D5D | Chr. 2B | Improving the resistance of plant to FHB and the tolerance to DON as well as potentially conjugating DON into D3G in plants [105]. |
| WFhb1-1 | Putative membrane protein | GenBank # KU304333.1) | In the interval of Qfhb1 | Overexpressing WFhb1-1 in non-Qfhb1-carrier wheat led to a significant resistance to Fg [122]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, C.; Chen, P.; Zhou, X.; Li, Y.; Ma, K.; Li, S.; Liu, H.; Li, L. Arms Race between the Host and Pathogen Associated with Fusarium Head Blight of Wheat. Cells 2022, 11, 2275. https://doi.org/10.3390/cells11152275
Hu C, Chen P, Zhou X, Li Y, Ma K, Li S, Liu H, Li L. Arms Race between the Host and Pathogen Associated with Fusarium Head Blight of Wheat. Cells. 2022; 11(15):2275. https://doi.org/10.3390/cells11152275
Chicago/Turabian StyleHu, Chunhong, Peng Chen, Xinhui Zhou, Yangchen Li, Keshi Ma, Shumei Li, Huaipan Liu, and Lili Li. 2022. "Arms Race between the Host and Pathogen Associated with Fusarium Head Blight of Wheat" Cells 11, no. 15: 2275. https://doi.org/10.3390/cells11152275