
Human Mesenchymal Stromal Cells Enhance Cartilage Healing in a Murine Joint Surface Injury Model

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Samples
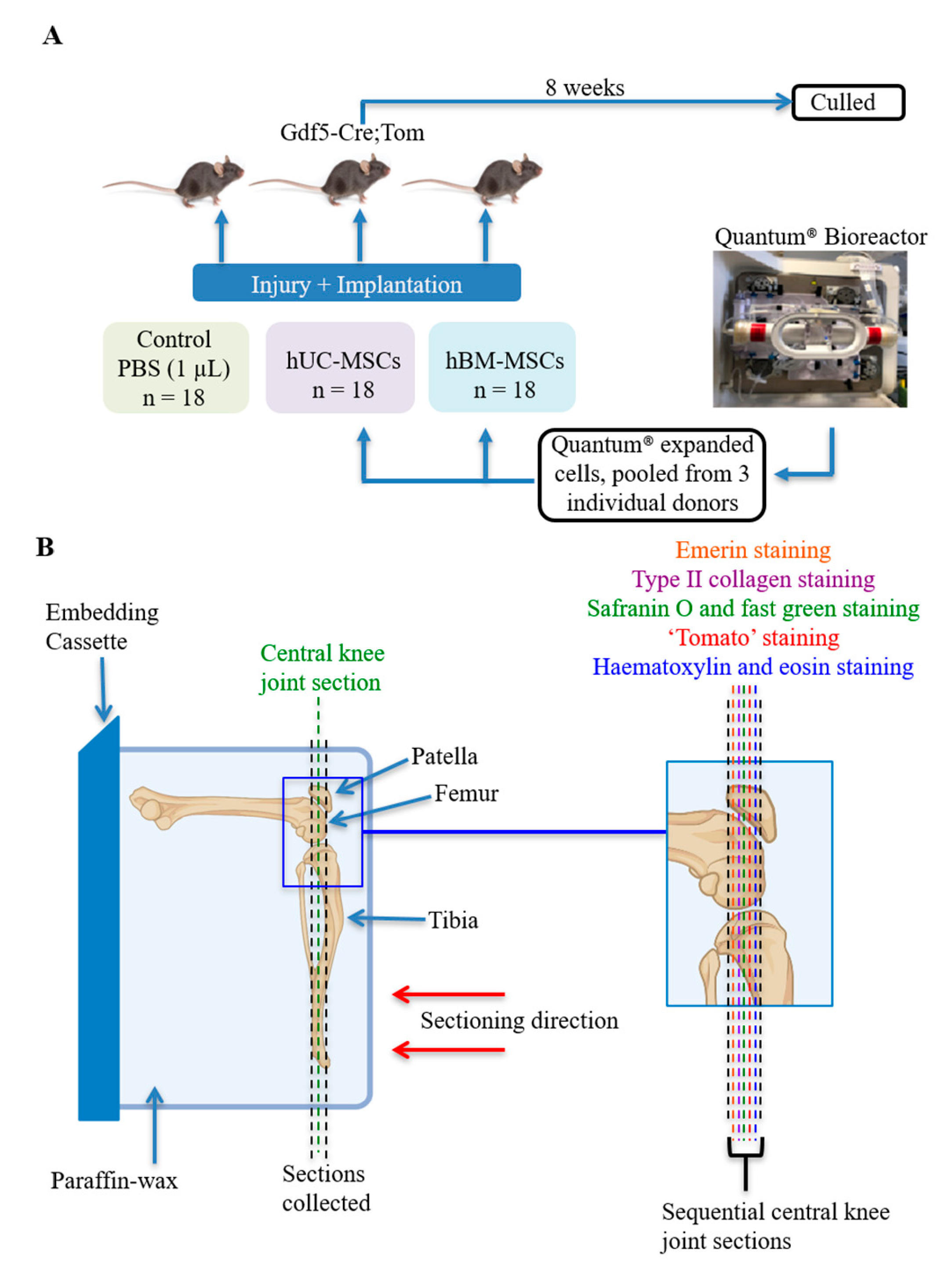
2.2. Mice and In Vivo Procedures
2.3. Histology, Immunohistochemistry and Electrochemiluminescence
2.4. Statistics
3. Results
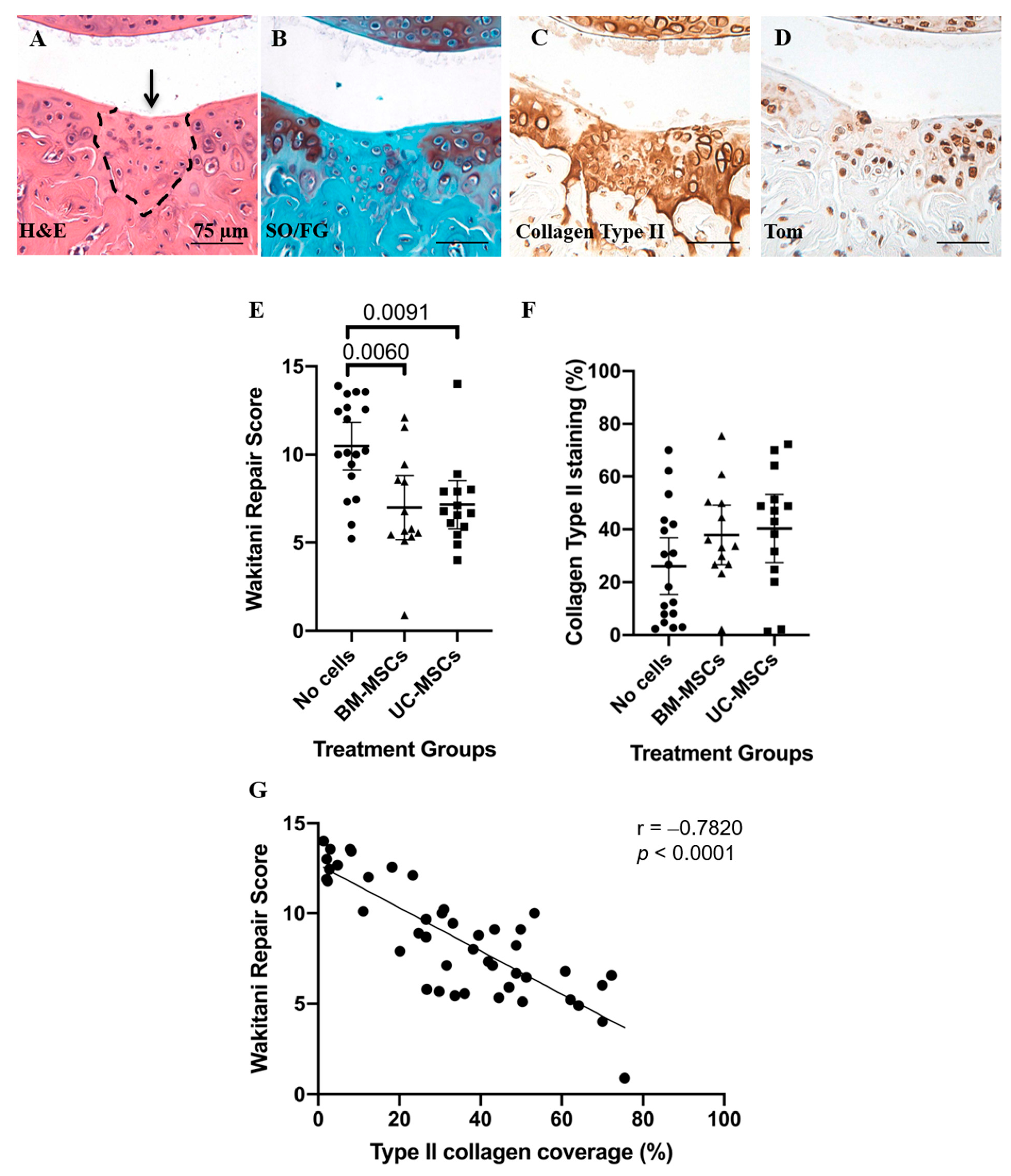
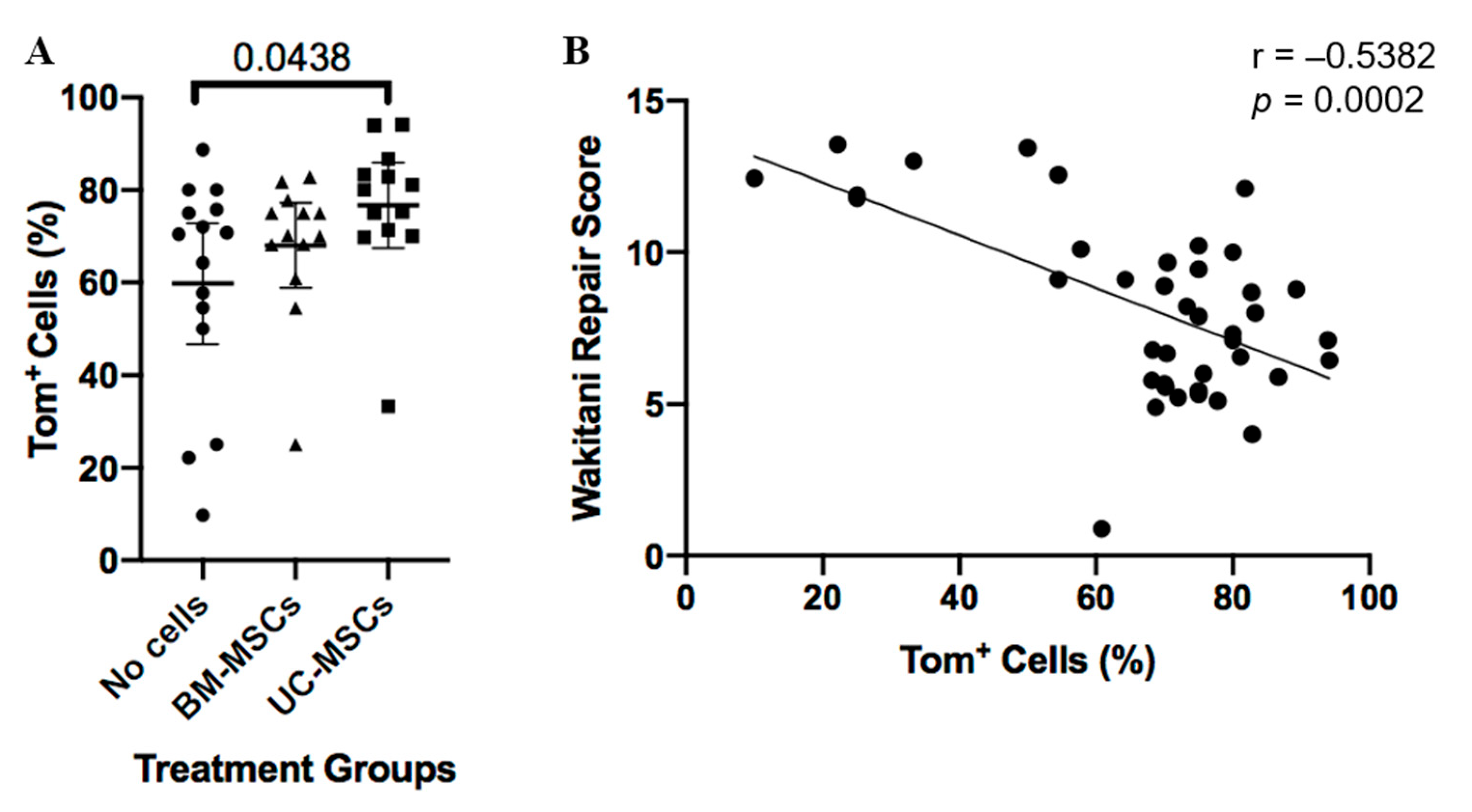
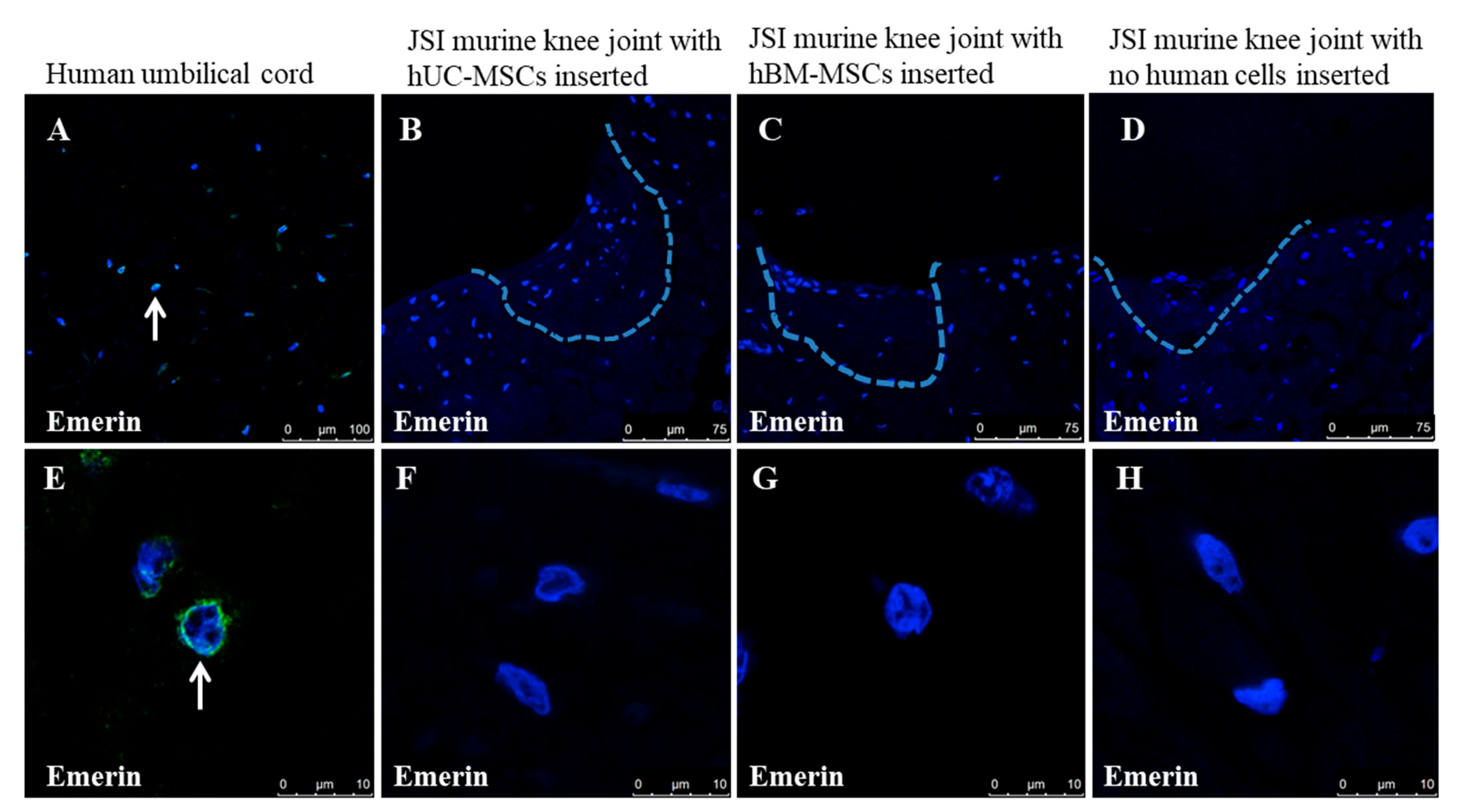
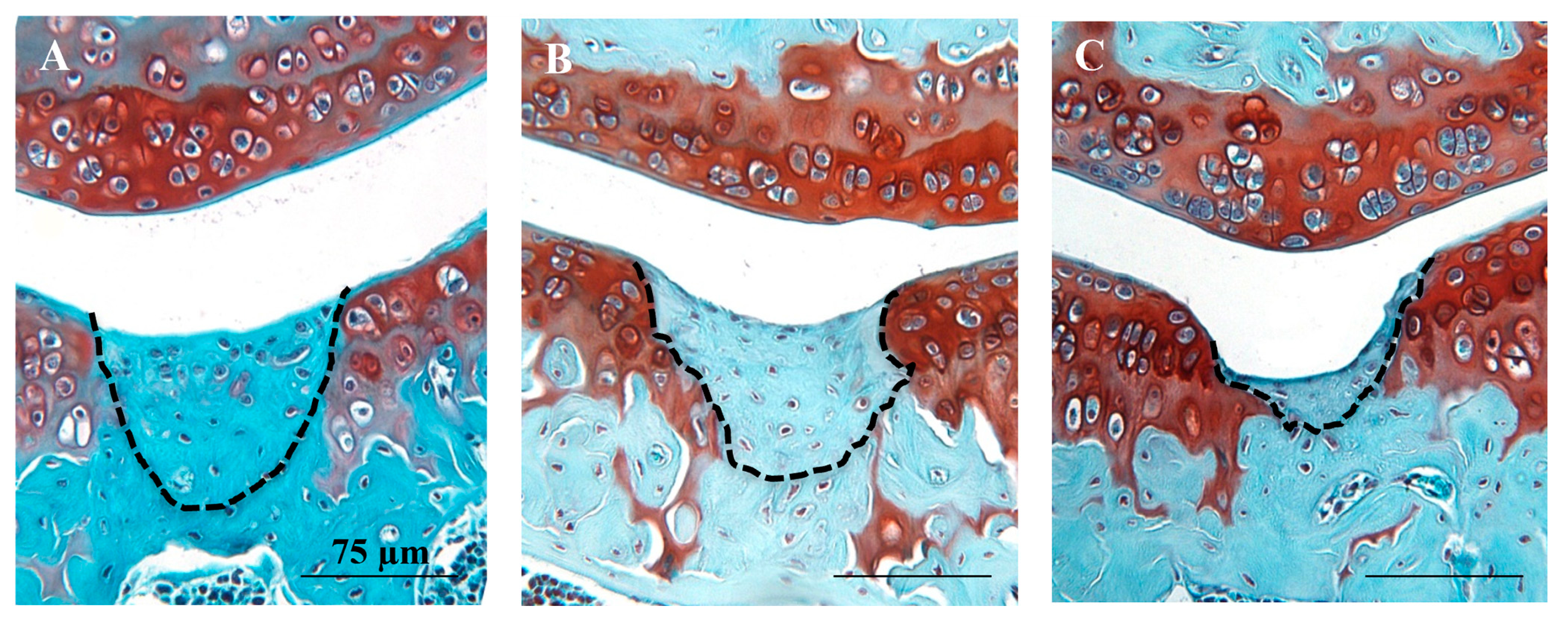
3.1. Histological and Immunohistochemical Analyses of Joint Changes after JSI
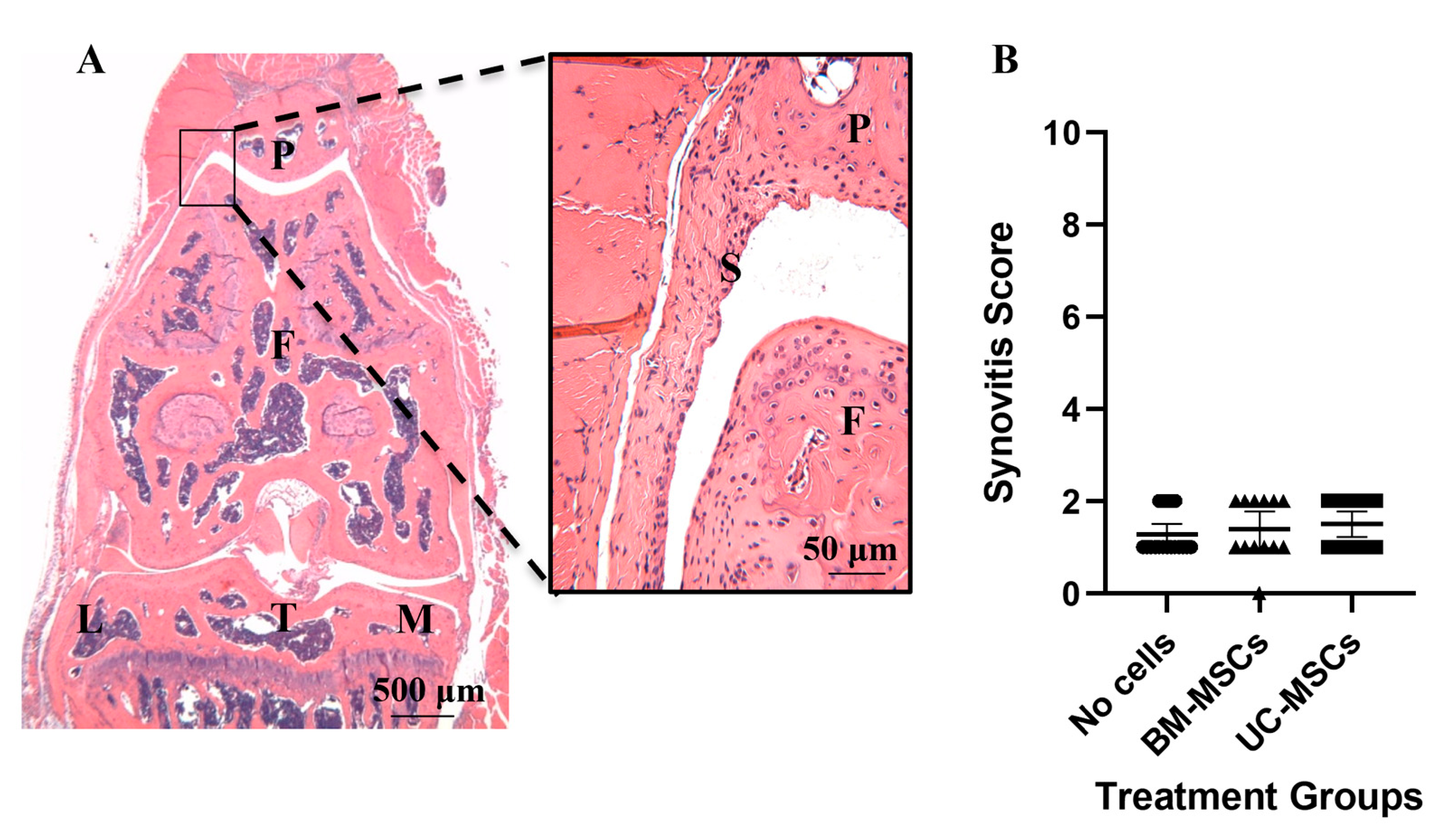
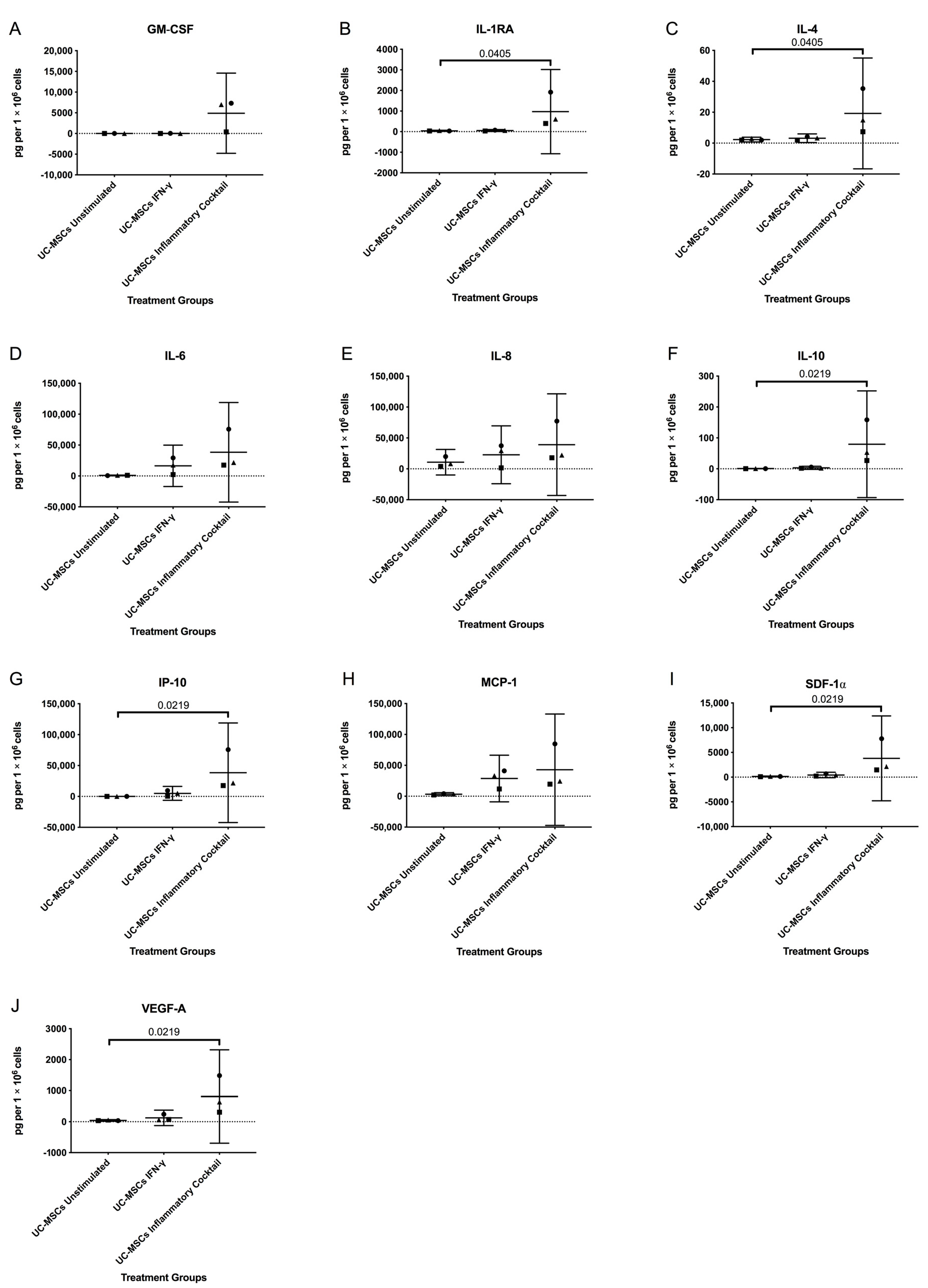
3.2. Inflammatory Markers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Score | Synovial Hyperplasia Severity # | Sub-Synovial Stroma * | Sub-Synovial Inflammation # | Synovial Exudate # |
|---|---|---|---|---|
| Description | Do not score the cells actually attached to the femur. Score the maximum hyperplasia. | Score the stroma adjacent to the synovial membrane. | Score the infiltration of inflammatory cells (neutrophils, macrophages and/or lymphocytes). | Infiltration of inflammatory cells (neutrophils, macrophages and/or lymphocytes) in the synovial cavity. |
| 0 | 1 cell thick | Synovial stroma shows normal cellularity. | No inflammatory cells | No inflammatory cells or fibrin in the synovial cavity. |
| 1 | Mild: 2–3 cells thick | The cellularity is slightly increased. | Occasional scattered inflammatory cells–or perivascular. | Inflammatory cells and/or fibrin colt in the synovial cavity–may be restricted to recesses. |
| 2 | Moderate: 4–5 cells thick | The cellularity is moderately increased, multinucleated cells might occur. | Focal areas of dense subsynovial WBC infiltrate–but still predominantly normal subsynovial areolar connective tissue present. | |
| 3 | Severe: >6 cells thick | The cellularity is greatly increased, multinucleated giant cells, pannus formation and rheumatoid granulomas might occur. | Widespread dense subsynovial WBC infiltrate–markedly reduced or little/no normal areolar connective tissue evident or lymphoid follicle formation. |
References
- Widuchowski, W.; Widuchowski, J.; Trzaska, T. Articular cartilage defects: Study of 25,124 knee arthroscopies. Knee 2007, 14, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Davies-Tuck, M.L.; Wluka, A.E.; Wang, Y.; Teichtahl, A.J.; Jones, G.; Ding, C.; Cicuttini, F.M. The natural history of cartilage defects in people with knee osteoarthritis. Osteoarthr. Cartil. 2008, 16, 337–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heir, S.; Nerhus, T.K.; Røtterud, J.H.; Løken, S.; Ekeland, A.; Engebretsen, L.; Årøen, A. Focal cartilage defects in the knee impair quality of life as much as severe osteoarthritis a comparison of knee injury and osteoarthritis outcome score in 4 patient categories scheduled for knee surgery. Am. J. Sports Med. 2010, No. 38, 231–237. [Google Scholar] [CrossRef] [Green Version]
- Minas, T.; Gomoll, A.H.; Solhpour, S.; Rosenberger, R.; Probst, C.; Bryant, T. Autologous chondrocyte implantation for joint preservation in patients with early osteoarthritis. Clin. Orthop. Relat. Res. 2010, 468, 147–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Registry, N.J. National joint registry—17th annual report 2020. Natl. Jt. Regist. 2020, 130–147. [Google Scholar]
- McCarthy, H.S.; Richardson, J.B.; Parker, J.C.E.; Roberts, S. Evaluating joint morbidity after chondral harvest for autologous chondrocyte implantation (ACI): A study of ACI-treated ankles and hips with a knee chondral harvest. Cartilage 2016, 7, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Brittberg, M.; Lindahl, A.; Nilsson, A.; Ohlsson, C.; Isaksson, O.; Peterson, L. Treatment of deep cartilage defects in the knee with autologous chondrocyte transplantation. N. Engl. J. Med. 1994, 331, 889–895. [Google Scholar] [CrossRef]
- Kim, S.H.; Djaja, Y.P.; Park, Y.-B.; Park, J.-G.; Ko, Y.-B.; Ha, C.-W. Intra-articular injection of culture-expanded mesenchymal stem cells without adjuvant surgery in knee osteoarthritis: A systematic review and meta-analysis. Am. J. Sports Med. 2020, 48, 2839–2849. [Google Scholar] [CrossRef] [PubMed]
- Mennan, C.; Wright, K.; Bhattacharjee, A.; Balain, B.; Richardson, J.; Roberts, S. Isolation and characterisation of mesenchymal stem cells from different regions of the human umbilical cord. BioMed Res. Int. Corp. 2013, 2013, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Mennan, C.; Brown, S.; McCarthy, H.; Mavrogonatou, E.; Kletsas, D.; Garcia, J.; Balain, B.; Richardson, J.; Roberts, S. Mesenchymal stromal cells derived from whole human umbilical cord exhibit similar properties to those derived from wharton’s jelly and bone marrow. FEBS Open Bio 2016, 6, 1054–1066. [Google Scholar] [CrossRef] [Green Version]
- Dabrowska, S.; Andrzejewska, A.; Janowski, M.; Lukomska, B. Immunomodulatory and regenerative effects of mesenchymal stem cells and extracellular vesicles: Therapeutic outlook for inflammatory and degenerative diseases. Front. Immunol. 2021, 11, 3809. [Google Scholar] [CrossRef]
- Mennan, C.; Garcia, J.; Roberts, S.; Hulme, C.; Wright, K. A comprehensive characterisation of large-scale expanded human bone marrow and umbilical cord mesenchymal stem cells. Stem Cell Res. Ther. 2019, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Criado, G.; Simelyte, E.; Inglis, J.J.; Essex, D.; Williams, R.O. Indoleamine 2, 3 Dioxygenase-mediated tryptophan catabolism regulates accumulation of Th1/Th17 cells in the joint in collagen-induced arthritis. Arthritis Rheum. 2009, 60, 1342–1351. [Google Scholar] [CrossRef]
- Perry, J.; McCarthy, H.S.; Bou-Gharios, G.; van ’t Hof, R.; Milner, P.I.; Mennan, C.; Roberts, S. Injected human umbilical cord-derived mesenchymal stromal cells do not appear to elicit an inflammatory response in a murine model of osteoarthritis. Osteoarthr. Cartil. Open 2020, 2, 100044. [Google Scholar] [CrossRef] [PubMed]
- Eltawil, N.M.; De Bari, C.; Achan, P.; Pitzalis, C.; Dell’Accio, F. A novel in vivo murine model of cartilage regeneration. Age and strain-dependent outcome after joint surface injury. Osteoarthr. Cartil. 2009, 17, 695–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roelofs, A.J.; Zupan, J.; Riemen, A.H.K.; Kania, K.; Ansboro, S.; White, N.; Clark, S.M.; De Bari, C. Joint morphogenetic cells in the adult mammalian synovium. Nat. Commun. 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The international society for cellular therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Rountree, R.B.; Schoor, M.; Chen, H.; Marks, M.E.; Harley, V.; Mishina, Y.; Kingsley, D.M. BMP receptor Signaling is required for postnatal maintenance of articular cartilage. PLoS Biol. 2004, 2, e355. [Google Scholar] [CrossRef] [Green Version]
- Madisen, L.; Zwingman, T.A.; Sunkin, S.M.; Oh, S.W.; Zariwala, H.A.; Gu, H.; Ng, L.L.; Palmiter, R.D.; Hawrylycz, M.J.; Jones, A.R.; et al. A robust and high-throughput Cre reporting and characterization system for the whole mouse brain. Nat. Neurosci. 2010, 13, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Wakitani, S.; Goto, T.; Pineda, S.J.; Young, R.G.; Mansour, J.M.; Caplan, A.I.; Goldberg, V.M. Mesenchymal cell-based repair of large, full-thickness defects of articular cartilage. J. Bone Jt. Surg. Ser. A 1994, 76, 579–592. [Google Scholar] [CrossRef]
- Kurth, T.B.; Dell’Accio, F.; Crouch, V.; Augello, A.; Sharpe, P.T.; De Bari, C. Functional mesenchymal stem cell niches in adult mouse knee joint synovium in vivo. Arthritis Rheumatol. 2011, 63, 1289–1300. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.T.; Moradi, B.; Zaki, S.; Smith, M.M.; McCracken, S.; Smith, S.M.; Jackson, C.J.; Little, C.B. Depletion of protease-activated receptor 2 but not protease-activated receptor 1 may confer protection against osteoarthritis in mice through extracartilaginous mechanisms. Arthritis Rheumatol. 2014, 66, 3337–3348. [Google Scholar] [CrossRef] [PubMed]
- Krenn, V.; Morawietz, L.; Burmester, G.R.; Kinne, R.W.; Mueller-Ladner, U.; Muller, B.; Haupl, T. Synovitis score: Discrimination between chronic low-grade and high-grade synovitis. Histopathology 2006, 49, 358–364. [Google Scholar] [CrossRef]
- Kurth, T.B.; De Bari, C. Immunostaining of skeletal tissues. Methods Mol. Biol. 2012, 816, 321–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manilal, S.; Nguyen, T.M.; Sewry, C.A.; Morris, G.E. The Emery-Dreifuss muscular dystrophy protein, emerin, is a nuclear membrane protein. Hum. Mol. Genet. 1996, 5, 801–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watt, F.E.; Paterson, E.; Freidin, A.; Kenny, M.; Judge, A.; Saklatvala, J.; Williams, A.; Vincent, T.L. Acute molecular changes in synovial fluid following human knee injury: Association with early clinical outcomes. Arthritis Rheumatol. 2016, 68, 2129–2140. [Google Scholar] [CrossRef]
- McGraw, K.O.; Wong, S.P. Forming inferences about some intraclass correlation coefficients. Psychol. Methods 1996, 1, 30–46. [Google Scholar] [CrossRef]
- Eliasziw, M.; Young, S.L.; Woodbury, M.G.; Fryday-Field, K. Statistical methodology for the concurrent assessment of interrater and intrarater reliability: Using goniometric measurements as an example. Phys. Ther. 1994, 74, 777–788. [Google Scholar] [CrossRef]
- Terpstra, T.J. The asymptotic normality and consistency of Kendall’s test against trend, when ties are present in one ranking. Proc. KNAW Ser. A 1952, 55, 327–333. [Google Scholar] [CrossRef] [Green Version]
- Jonckheere, A.R. A distribution-free k-sample test against ordered alternatives. Biometrika 1954, 41, 133–145. [Google Scholar] [CrossRef]
- Malik, N. Allogeneic versus autologous stem-cell therapy: A comparison of manufacturing costs and commercialization challenges. BioPharm Int. 2012, 25, 36–40. [Google Scholar]
- Chen, H.; Capellini, T.D.; Schoor, M.; Mortlock, D.P.; Reddi, A.H.; Kingsley, D.M. Heads, shoulders, elbows, knees, and toes: Modular Gdf5 enhancers control different joints in the vertebrate skeleton. PLoS Genet. 2016, 12, e1006454. [Google Scholar] [CrossRef] [Green Version]
- Pregizer, S.K.; Kiapour, A.M.; Young, M.; Chen, H.; Schoor, M.; Liu, Z.; Cao, J.; Rosen, V.; Capellini, T.D. Impact of broad regulatory regions on gdf5 expression and function in knee development and susceptibility to osteoarthritis. Ann. Rheum. Dis. 2018, 77, 450. [Google Scholar] [CrossRef] [PubMed]
- Morales, T.I. Chondrocyte moves: Clever strategies? Osteoarthr. Cartil. 2007, 15, 861–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowthwaite, G.P.; Bishop, J.C.; Redman, S.N.; Khan, I.M.; Rooney, P.; Evans, D.J.R.; Haughton, L.; Bayram, Z.; Boyer, S.; Thomson, B.; et al. The surface of articular cartilage contains a progenitor cell populations. J. Cell Sci. 2004, 117, 889–897. [Google Scholar] [CrossRef] [Green Version]
- Williams, R.; Khan, I.M.; Richardson, K.; Nelson, L.; McCarthy, H.E.; Analbelsi, T.; Singhrao, S.K.; Dowthwaite, G.P.; Jones, R.E.; Baird, D.M.; et al. Identification and clonal characterisation of a progenitor cell sub-population in normal human articular cartilage. PLoS ONE 2010, 5, e13246. [Google Scholar] [CrossRef] [Green Version]
- Kozhemyakina, E.; Zhang, M.; Ionescu, A.; Ayturk, U.M.; Ono, N.; Kobayashi, A.; Kronenberg, H.; Warman, M.L.; Lassar, A.B. Identification of a Prg4-expressing articular cartilage progenitor cell population in mice. Arthritis Rheumatol. 2015, 67, 1261–1273. [Google Scholar] [CrossRef] [Green Version]
- De Bari, C.; Dell’Accio, F.; Tylzanowski, P.; Luyten, F.P. Multipotent mesenchymal stem cells from adult human synovial membrane. Arthritis Rheum. 2001, 44, 1928–1942. [Google Scholar] [CrossRef]
- Decker, R.S.; Um, H.-B.; Dyment, N.A.; Cottingham, N.; Usami, Y.; Enomoto-Iwamoto, M.; Kronenberg, M.S.; Maye, P.; Rowe, D.W.; Koyama, E.; et al. Cell origin, volume and arrangement are drivers of articular cartilage formation, morphogenesis and response to injury in mouse limbs. Dev. Biol. 2017, 426, 56–68. [Google Scholar] [CrossRef]
- Garcia, J.; Hulme, C.; Mennan, C.; Roberts, S.; Bastiaansen-Jenniskens, Y.M.; van Osch, G.J.V.M.; Tins, B.; Gallacher, P.; Wright, K. The synovial fluid from patients with focal cartilage defects contains mesenchymal stem/stromal cells and macrophages with pro- and anti-inflammatory phenotypes. Osteoarthr. Cartil. Open 2020, 2, 100039. [Google Scholar] [CrossRef]
- Hollander, A.P.; Dickinson, S.C.; Sims, T.J.; Brun, P.; Cortivo, R.; Kon, E.; Marcacci, M.; Zanasi, S.; Borrione, A.; De Luca, C.; et al. Maturation of tissue engineered cartilage implanted in injured and osteoarthritic human knees. Tissue Eng. 2006, 12, 1787–1798. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.; Menage, J.; Sandell, L.J.; Evans, E.H.; Richardson, J.B. Immunohistochemical study of collagen types I and II and procollagen IIA in human cartilage repair tissue following autologous chondrocyte implantation. Knee 2009, 16, 398–404. [Google Scholar] [CrossRef] [Green Version]
- Ozeki, N.; Muneta, T.; Koga, H.; Nakagawa, Y.; Mizuno, M.; Tsuji, K.; Mabuchi, Y.; Akazawa, C.; Kobayashi, E.; Matsumoto, K.; et al. Not single but periodic injections of synovial mesenchymal stem cells maintain viable cells in knees and inhibit osteoarthritis progression in rats. Osteoarthr. Cartil. 2016, 24, 1061–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancuso, P.; Raman, S.; Glynn, A.; Barry, F.; Murphy, J.M. Mesenchymal stem cell therapy for osteoarthritis: The critical role of the cell secretome. Front. Bioeng. Biotechnol. 2019, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Caplan, A.I. Mesenchymal stem cells: Time to change the name! Stem Cells Transl. Med. 2017, 6, 1445–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, A.; Hansen, A.K.; Mennan, C.; Martinez-Zubiaurre, I. Mesenchymal stromal cells from human umbilical cords display poor chondrogenic potential in scaffold-free three dimensional cultures. Eur. Cells Mater. 2016, 31, 407–424. [Google Scholar] [CrossRef]
- Niemeyer, P.; Schönberger, T.S.; Hahn, J.; Kasten, P.; Fellenberg, J.; Suedkamp, N.; Mehlhorn, A.T.; Milz, S.; Pearce, S. Xenogenic Transplantation of human mesenchymal stem cells in a critical size defect of the sheep tibia for bone regeneration. Tissue Eng. Part A 2010, 16, 33–43. [Google Scholar] [CrossRef]
- Saris, T.F.F.; de Windt, T.S.; Kester, E.C.; Vonk, L.A.; Custers, R.J.H.; Saris, D.B.F. Five-year outcome of 1-stage cell-based cartilage repair using recycled autologous chondrons and allogenic mesenchymal stromal cells: A first-in-human clinical trial. Am. J. Sports Med. 2021, 49, 941–947. [Google Scholar] [CrossRef]
- Chung, Y.W.; Yang, H.Y.; Kang, S.J.; Song, E.K.; Seon, J.K. Allogeneic umbilical cord blood-derived mesenchymal stem cells combined with high tibial osteotomy: A retrospective study on safety and early results. Int. Orthop. 2021, 45, 481–488. [Google Scholar] [CrossRef]
- Sotiropoulou, P.A.; Perez, S.A.; Gritzapis, A.D.; Baxevanis, C.N.; Papamichail, M. Interactions between human mesenchymal stem cells and natural killer cells. Stem Cells 2006, 24, 74–85. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perry, J.; Roelofs, A.J.; Mennan, C.; McCarthy, H.S.; Richmond, A.; Clark, S.M.; Riemen, A.H.K.; Wright, K.; De Bari, C.; Roberts, S. Human Mesenchymal Stromal Cells Enhance Cartilage Healing in a Murine Joint Surface Injury Model. Cells 2021, 10, 1999. https://doi.org/10.3390/cells10081999
Perry J, Roelofs AJ, Mennan C, McCarthy HS, Richmond A, Clark SM, Riemen AHK, Wright K, De Bari C, Roberts S. Human Mesenchymal Stromal Cells Enhance Cartilage Healing in a Murine Joint Surface Injury Model. Cells. 2021; 10(8):1999. https://doi.org/10.3390/cells10081999
Chicago/Turabian StylePerry, Jade, Anke J. Roelofs, Claire Mennan, Helen S. McCarthy, Alison Richmond, Susan M. Clark, Anna H. K. Riemen, Karina Wright, Cosimo De Bari, and Sally Roberts. 2021. "Human Mesenchymal Stromal Cells Enhance Cartilage Healing in a Murine Joint Surface Injury Model" Cells 10, no. 8: 1999. https://doi.org/10.3390/cells10081999