
Early Drosophila Oogenesis: A Tale of Centriolar Asymmetry

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Drosophila Strains
2.2. Transmission Electron Microscopy
3. Results
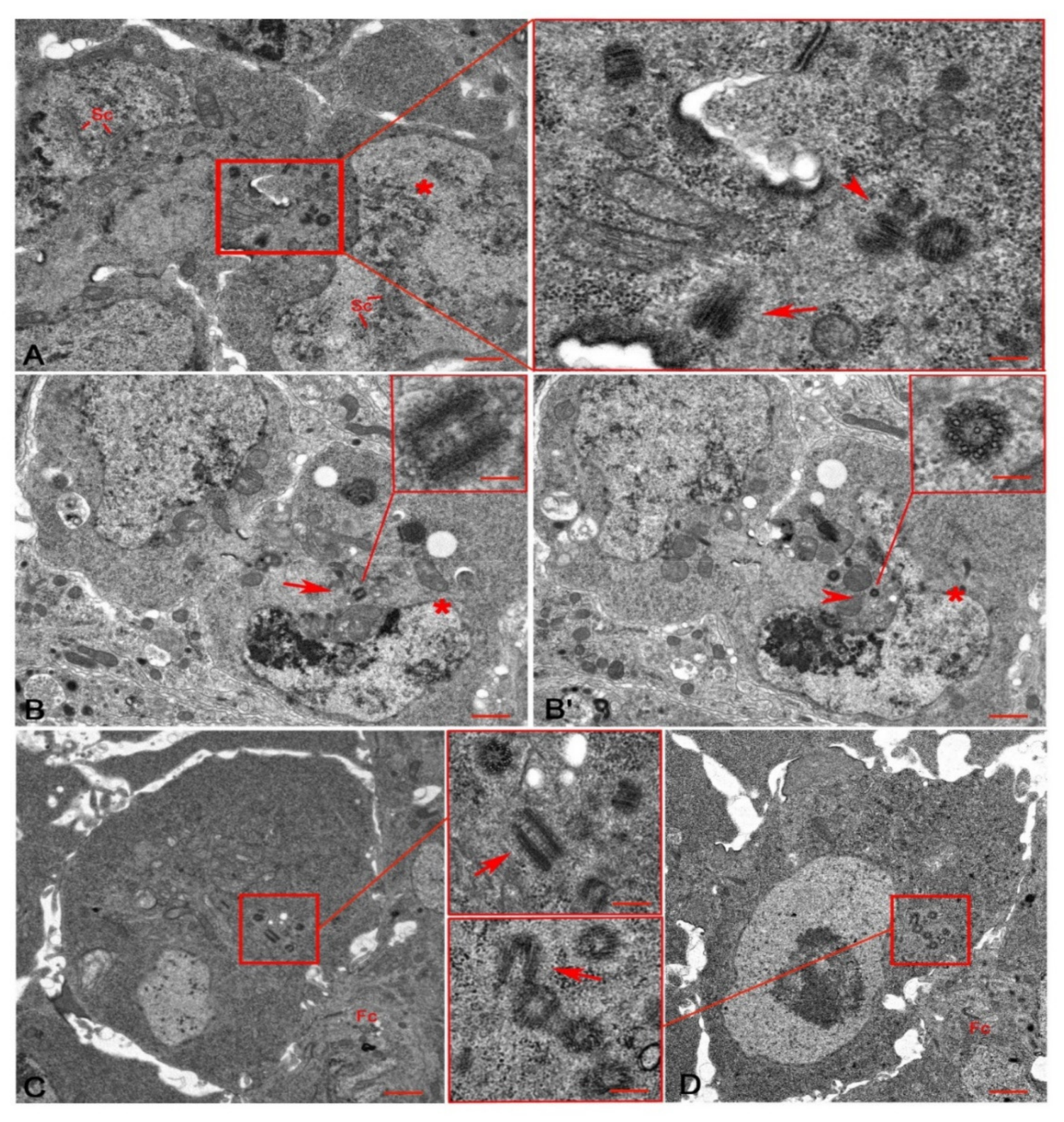
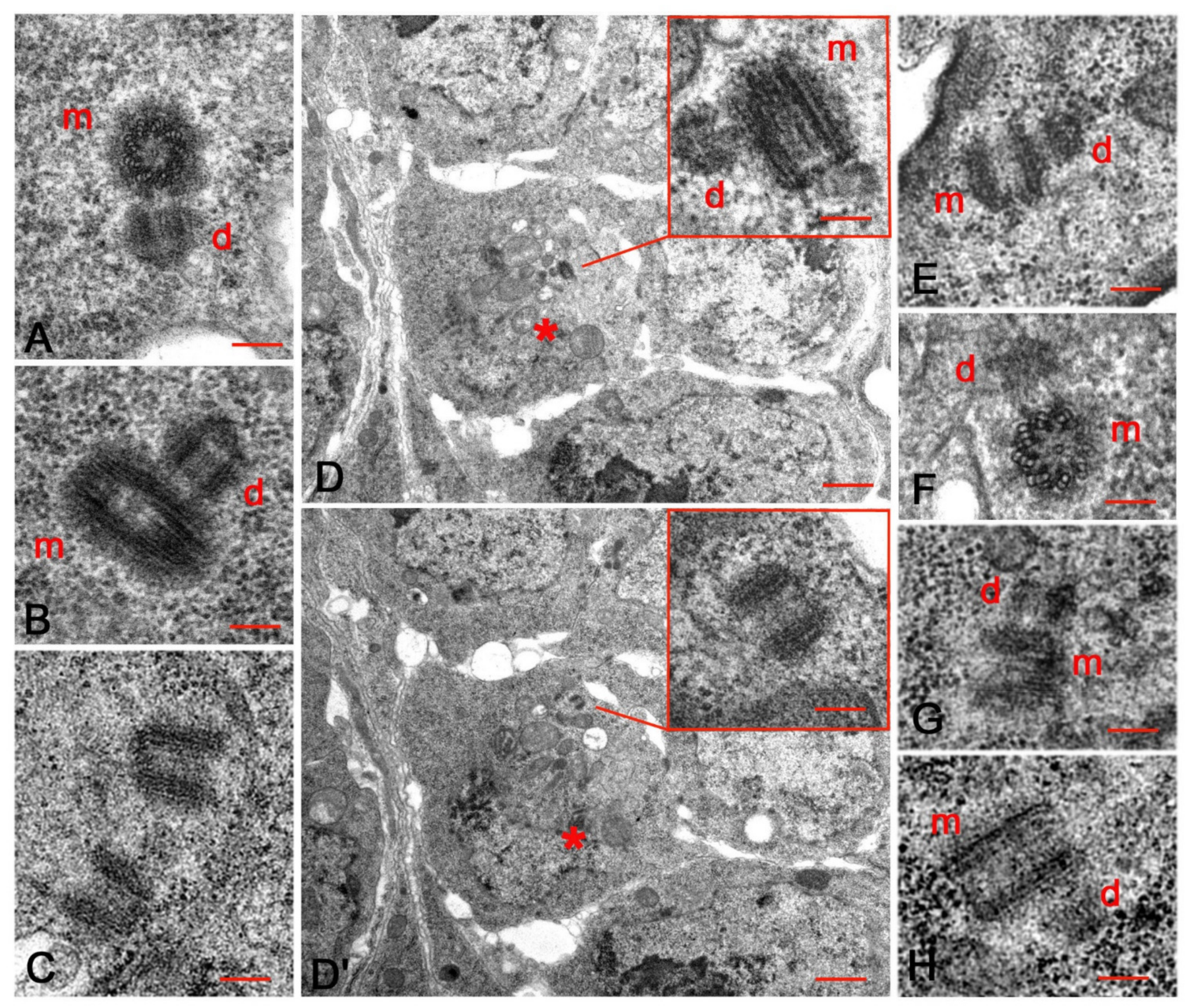
3.1. An Unusual Centriole in the Oocyte Cytoplasm
3.2. Centriole Duplication during Early Oogenesis
3.3. The Centriole Structure in the Drosophila Ovariole
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Spradling, A. Developmental genetics of oogenesis. In The Development of Drosophila Melanogaster, 1st ed.; Bate, M., Martinez-Arias, A., Eds.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1993; Volume 1, pp. 1–70. [Google Scholar]
- Roth, S.; Lynch, J.A. Symmetry breaking during Drosophila oogenesis. Cold Spring Harb. Perspect. Biol. 2009, 1, a001891. [Google Scholar] [CrossRef] [Green Version]
- Hinnant, T.D.; Merkle, J.A.; Ables, E.T. Coordinating Proliferation, Polarity, and Cell Fate in the Drosophila Female Germline. Front. Front. Cell Dev. Biol. 2020, 8, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huynh, J.R.; St Johnston, D. The role of BicD, Egl, Orb and the microtubules in the restriction of meiosis to the Drosophila oocyte. Development 2000, 127, 2785–2794. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, A.T. Electron microscopy of meiosis in Drosophila melanogaster females. I. Structure, arrangement, and temporal change of the synaptonemal complex in wild-type. Chromosoma 1975, 51, 157–182. [Google Scholar] [CrossRef] [PubMed]
- Huynh, J.-R.; St Johnston, D. The Origin of Asymmetry: Early Polarisation of the Drosophila Germline Cyst and Oocyte. Curr. Biol. 2004, 14, R438–R449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grieder, N.C.; de Cuevas, M.; Spradling, A.C. The fusome organizes the microtubule network during oocyte differentiation in Drosophila. Development 2000, 127, 4253–4264. [Google Scholar] [CrossRef]
- Megraw, T.L.; Kaufman, T.C. The centrosome in Drosophila oocyte development. Curr. Top. Dev. Biol. 2000, 49, 385–407. [Google Scholar]
- Bolivar, J.; Huynh, J.R.; Lopez-Schier, H.; González, C.; St Johnston, D.; González-Reyes, A. Centrosome migration into the Drosophila oocyte is independent of BicD, egl and of the organisation of the microtubule cytoskeleton. Development 2001, 128, 1889–1897. [Google Scholar] [CrossRef] [PubMed]
- Theurkauf, W.; Alberts, M.; Jan, Y.; Jongens, T. A central role for microtubules in the differentiation of Drosophila oocytes. Development 1993, 118, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Koch, E.; Spitzer, R. Multiple effects of colchicine on oogenesis in Drosophila; Induced sterility and switch of potencial oocyte to nurse-cell developmental pathway. Cell Tissue Res. 1983, 228, 21–32. [Google Scholar] [CrossRef]
- Mahowald, A.P.; Strassheim, J.M. Intercellular migration of centrioles in the germarium of Drosophila melanogaster: An electron microscopic study. J. Cell Biol. 1970, 45, 306–320. [Google Scholar] [CrossRef] [Green Version]
- Januschke, J.; Gervais, L.; Gillet, L.; Keryer, G.; Bornens, M.; Guichet, A. The centrosome-nucleus complex and microtubule organization in the Drosophila oocyte. Development 2006, 133, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Pimenta-Marques, A.; Bento, I.; Lopes, C.A.; Duarte, P.; Jana, S.C.; Bettencourt-Dias, M. A mechanism for the elimination of the female gamete centrosome in Drosophila melanogaster. Science 2016, 353, aaf4866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roper, K.; Brown, N.H. A Spectraplakin Is Enriched on the Fusome and Organizes Microtubules during Oocyte Specification in Drosophila. Curr. Biol. 2004, 14, 99–110. [Google Scholar] [CrossRef]
- Lin, H.; Spradling, A.C. Fusome asymmetry and oocyte determination in Drosophila. Dev. Genet. 1995, 16, 6–12. [Google Scholar] [CrossRef]
- Koch, E.A.; King, R.C. The origin and early differentiation of the egg chamber of Drosophila melanogaster. J. Morphol. 1966, 119, 283–303. [Google Scholar] [CrossRef]
- de Cuevas, M.; Spradling, A.C. Morphogenesis of the Drosophila fusome and its implications for oocyte specification. Development 1998, 125, 2781–2789. [Google Scholar] [CrossRef] [PubMed]
- Gottardo, M.; Callaini, G.; Riparbelli, M.G. The Drosophila centriole—conversion of doublets into triplets within the stem cell niche. J. Cell Sci. 2015, 128, 2437–2442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galletta, B.J.; Jacobs, K.C.; Fagerstrom, C.J.; Rusan, N.M. Asterless is required for centriole length control and sperm development. J. Cell Biol. 2016, 213, 435–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Inaba, M.; Venkei, Z.G.; Yamashita, Y.M. Klp10A, a stem cell centrosome-enriched kinesin, balances asymmetries in Drosophila male germline stem cell division. Elife 2016, 5, e20977. [Google Scholar] [CrossRef]
- Gottardo, M.; Callaini, G.; Riparbelli, M.G. Klp10A modulates the localization of centriole-associated proteins during Drosophila male gametogenesis. Cell Cycle 2016, 15, 3432–3441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, Y.M.; Mahowald, A.P.; Perlin, J.R.; Fuller, M.T. Asymmetric inheritance of mother versus daughter centrosome in stem cell division. Science 2007, 315, 518–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salzmann, V.; Chen, C.; Chiang, C.Y.; Tiyaboonchai, A.; Mayer, M.; Yamashita, Y.M. Centrosome-dependent asymmetric inheritance of the midbody ring in Drosophila germline stem cell division. Mol. Biol. Cell 2014, 25, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Theurkauf, W.E. Microtubules and cytoplasm organization during Drosophila oogenesis. Dev. Biol. 1994, 165, 352–360. [Google Scholar] [CrossRef]
- Basto, R.; Lau, J.; Vinogradova, T.; Gardiol, A.; Woods, C.G.; Khodjakov, A.; Raff, J.W. Flies without centrioles. Cell 2006, 125, 1375–1386. [Google Scholar] [CrossRef] [Green Version]
- Stevens, N.R.; Raposo, A.A.; Basto, R.; St Johnston, D.; Raff, J.W. From stem cell to embryo without centrioles. Curr. Biol. 2007, 17, 1498–1503. [Google Scholar] [CrossRef] [Green Version]
- Uzbekov, R.E.; Avidor-Reiss, T. Principal Postulates of Centrosomal Biology. Version 2020. Cells 2020, 9, 2156. [Google Scholar] [CrossRef]
- Riparbelli, M.G.; Colozza, G.; Callaini, G. Procentriole elongation and recruitment of pericentriolar material are downregulated in cyst cells as they enter quiescence. J. Cell Sci. 2009, 122, 3613–3618. [Google Scholar] [CrossRef] [Green Version]
- Riparbelli, M.G.; Persico, V.; Gottardo, M.; Callaini, G. The developing Drosophila eye—a new model to study centriole reduction. J. Cell Sci. 2018, 131, jcs211441. [Google Scholar] [CrossRef] [Green Version]
- Riparbelli, M.G.; Whitfield, W.G.; Dallai, R.; Callaini, G. Assembly of the zygotic centrosome in the fertilized Drosophila egg. Mech. Dev. 1997, 65, 135–144. [Google Scholar] [CrossRef]
- Gottardo, M.; Persico, V.; Callaini, G.; Riparbelli, M.G. The “transition zone” of the cilium-like regions in the Drosophila spermatocytes and the role of the C-tubule in axoneme assembly. Exp. Cell Res. 2018, 371, 262–268. [Google Scholar] [CrossRef]
- Huettner, A.F. Maturation and fertilization in Drosophila melanogaster. J. Morphol. 1924, 39, 249–265. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riparbelli, M.G.; Persico, V.; Callaini, G. Early Drosophila Oogenesis: A Tale of Centriolar Asymmetry. Cells 2021, 10, 1997. https://doi.org/10.3390/cells10081997
Riparbelli MG, Persico V, Callaini G. Early Drosophila Oogenesis: A Tale of Centriolar Asymmetry. Cells. 2021; 10(8):1997. https://doi.org/10.3390/cells10081997
Chicago/Turabian StyleRiparbelli, Maria Giovanna, Veronica Persico, and Giuliano Callaini. 2021. "Early Drosophila Oogenesis: A Tale of Centriolar Asymmetry" Cells 10, no. 8: 1997. https://doi.org/10.3390/cells10081997