
Germ Cell Derivation from Pluripotent Stem Cells for Understanding In Vitro Gametogenesis



{kind=link}
{kind=link}
Abstract
:1. Introduction
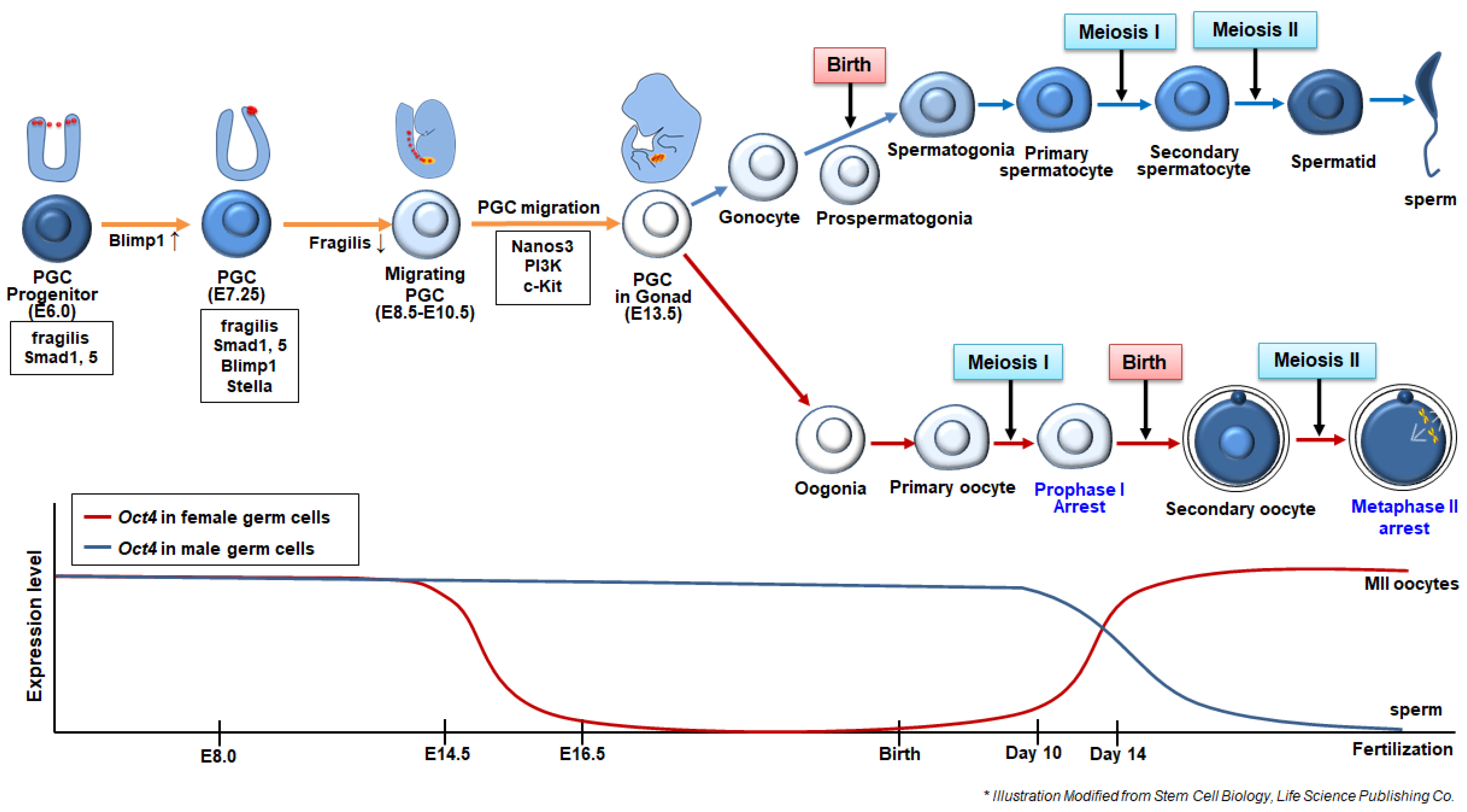
2. Germ Cell Differentiation Pathway in Mammals
2.1. Specification and Migration of Primordial Germ Cells (PGCs)
2.2. Sex Determination and Maturation of Germ Cells
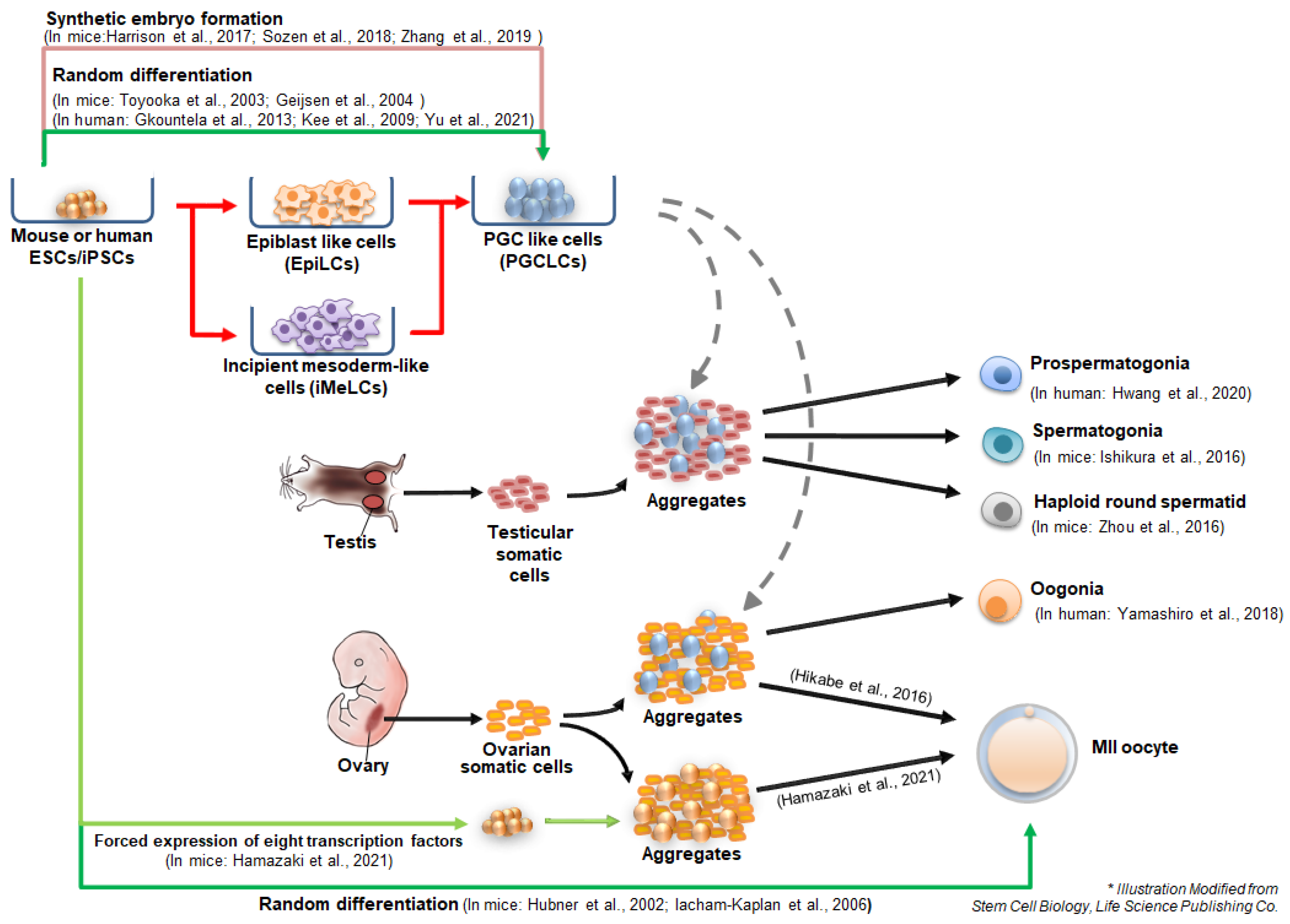
3. Germ Cell Differentiation from Pluripotent Stem Cells In Vitro
3.1. In Vitro Derivation of PGC-Like Cells (PGCLCs) from Pluripotent Stem Cells
3.2. In Vitro Derivation of Spermatogonia and Oogonia from Pluripotent Stem Cells
3.3. In Vitro Derivation of Haploid Spermatid-Like Cells from Pluripotent Stem Cells
3.4. In Vitro Oogenesis from Pluripotent Stem Cells
4. Germ Cell Derivation via Synthetic Embryo Formation
5. In Vitro Gametogenesis for Clinical Applications
6. Safety and Ethical Concerns
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hübner, K.; Fuhrmann, G.; Christenson, L.K.; Kehler, J.; Reinbold, R.; De La Fuente, R.; Wood, J.; Iii, J.F.S.; Boiani, M.; Schöler, H. Derivation of Oocytes from Mouse Embryonic Stem Cells. Science 2003, 300, 1251–1256. [Google Scholar] [CrossRef] [Green Version]
- Marques-Mari, A.; Lacham-Kaplan, O.; Medrano, J.; Pellicer, A.; Simon, C. Differentiation of germ cells and gametes from stem cells. Hum. Reprod. Updat. 2009, 15, 379–390. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, K.; Ohta, H.; Kurimoto, K.; Aramaki, S.; Saitou, M. Reconstitution of the Mouse Germ Cell Specification Pathway in Culture by Pluripotent Stem Cells. Cell 2011, 146, 519–532. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, K.; Ogushi, S.; Kurimoto, K.; Shimamoto, S.; Ohta, H.; Saitou, M. Offspring from Oocytes Derived from in Vitro Primordial Germ Cell-like Cells in Mice. Sciene 2012, 338, 971–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Fertility Rate 1950–2021. Available online: https://www.macrotrends.net/countries/WLD/world/fertility-rate (accessed on 28 January 2021).
- Pera, R.A.R.; Simon, C.; Medrano, J.V. Germ Cell Differentiation from Pluripotent Cells. Semin. Reprod. Med. 2013, 31, 014–023. [Google Scholar] [CrossRef] [Green Version]
- Lawson, K.A.; Hage, W.J. Clonal Analysis of the Origin of Primordial Germ Cells in the Mouse. Novartis Found. Symp. 2007, 182, 68–91. [Google Scholar] [CrossRef]
- Ohinata, Y.; Payer, B.; O’Carroll, D.; Ancelin, K.; Ono, Y.; Sano, M.; Barton, S.C.; Obukhanych, T.; Nussenzweig, M.C.; Tarakhovsky, A.; et al. Blimp1 is a critical determinant of the germ cell lineage in mice. Nat. Cell Biol. 2005, 436, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, M.; Sasaoka, Y.; Kiso, M.; Abe, K.; Haraguchi, S.; Kobayashi, S.; Saga, Y. Conserved Role of nanos Proteins in Germ Cell Development. Science 2003, 301, 1239–1241. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Lopes, S.M.C.D.S.; Surani, A.; Chung, H.-Y.; Weinberger, M.B.; Levine, J.B.; Kavner, A.; Yang, J.-M.; Tolbert, S.H.; Kaner, R.B. Germ Cell Specification in Mice. Science 2007, 316, 394–396. [Google Scholar] [CrossRef]
- Matsui, Y.; Zsebo, K.M.; Hogan, B.L.M. Embryonic expression of a haematopoietic growth factor encoded by the SI locus and the ligand for c-kit. Nat. Cell Biol. 1990, 347, 667–669. [Google Scholar] [CrossRef]
- Surani, M.A. Reprogramming of genome function through epigenetic inheritance. Nat. Cell Biol. 2001, 414, 122–128. [Google Scholar] [CrossRef]
- von Meyenn, F.; Berrens, R.V.; Andrews, S.; Santos, F.; Collier, A.; Krueger, F.; Osorno, R.; Dean, W.; Rugg-Gunn, P.; Reik, W. Comparative Principles of DNA Methylation Reprogramming during Human and Mouse In Vitro Primordial Germ Cell Specification. Dev. Cell 2016, 39, 104–115. [Google Scholar] [CrossRef] [Green Version]
- Tam, P.P.L.; Snow, M.H.L. Proliferation and migration of primordial germ cells during compensatory growth in mouse embryos. J. Embryol Exp. Morphol. 1981, 64, 133–147. [Google Scholar] [CrossRef]
- Hamer, G.; De Rooij, D.G. Mutations causing specific arrests in the development of mouse primordial germ cells and gonocytes. Biol. Reprod. 2018, 99, 75–86. [Google Scholar] [CrossRef]
- Donovan, P. Migratory and postmigratory mouse primordial germ cells behave differently in culture. Cell 1986, 44, 831–838. [Google Scholar] [CrossRef]
- Culty, M. Gonocytes, from the Fifties to the Present: Is There a Reason to Change the Name? Biol. Reprod. 2013, 89, 46. [Google Scholar] [CrossRef]
- Schöler, H.; Ruppert, S.; Suzuki, N.; Chowdhury, K.; Gruss, P. New type of POU domain in germ line-specific protein Oct-4. Nat. Cell Biol. 1990, 344, 435–439. [Google Scholar] [CrossRef]
- Pesce, M.; Wang, X.; Wolgemuth, D.J.; Schöler, H. Differential expression of the Oct-4 transcription factor during mouse germ cell differentiation. Mech. Dev. 1998, 71, 89–98. [Google Scholar] [CrossRef]
- Kehler, J.; Tolkunova, E.; Koschorz, B.; Pesce, M.; Gentile, L.; Boiani, M.; Lomelí, H.; Nagy, A.; McLaughlin, K.J.; Schöler, H.; et al. Oct4 is required for primordial germ cell survival. EMBO Rep. 2004, 5, 1078–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.W.; Joo, J.Y.; Hong, Y.J.; Kim, J.S.; Song, H.; Lee, J.W.; Wu, G.; Schöler, H.R.; Do, J.T. Distinct Enhancer Activity of Oct4 in Naive and Primed Mouse Pluripotency. Stem Cell Rep. 2016, 7, 911–926. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Oron, E.; Nelson, B.; Razis, S.; Ivanova, N. Distinct Lineage Specification Roles for NANOG, OCT4, and SOX2 in Human Embryonic Stem Cells. Cell Stem Cell 2012, 10, 440–454. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, S.; Kimura, H.; Tada, M.; Nakatsuji, N.; Tada, T. Nanog expression in mouse germ cell development. Gene Expr. Patterns 2005, 5, 639–646. [Google Scholar] [CrossRef]
- Wen, L.; Tang, F. Human Germline Cell Development: From the Perspective of Single-Cell Sequencing. Mol. Cell 2019, 76, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Witschi, E. Migration of the Germ Cells of Human Embryos from the Yolk Sac to the Primitive Gonadal Folds; Carnegie Institution of Washington: Washington, DC, USA, 1948. [Google Scholar]
- Tang, W.W.; Kobayashi, T.; Irie, N.; Dietmann, S.; Surani, M.A. Specification and epigenetic programming of the human germ line. Nat. Rev. Genet. 2016, 17, 585–600. [Google Scholar] [CrossRef] [PubMed]
- Do, J.T. Stem Cell Biology; Life Science Publishing Co.: Seoul, Korea, 2019. [Google Scholar]
- Koopman, P.; Gubbay, J.; Vivian, N.; Goodfellow, P.; Lovell-Badge, R. Male development of chromosomally female mice transgenic for Sry. Nat. Cell Biol. 1991, 351, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Burgoyne, P.S. Role of mammalian Y chromosome in sex determination. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1988, 322, 63–72. [Google Scholar] [CrossRef]
- Swain, A.; Lovell-Badge, R. A molecular approach to sex determination in mammals. Acta. Paediatr. 1997, 86, 46–49. [Google Scholar] [CrossRef]
- Maatouk, D.M.; DiNapoli, L.; Alvers, A.L.; Parker, K.L.; Taketo, M.M.; Capel, B. Stabilization of β-catenin in XY gonads causes male-to-female sex-reversal. Hum. Mol. Genet. 2008, 17, 2949–2955. [Google Scholar] [CrossRef] [Green Version]
- Uhlenhaut, H.; Jakob, S.; Anlag, K.; Eisenberger, T.; Sekido, R.; Kress, J.; Treier, A.-C.; Klugmann, C.; Klasen, C.; Holter, N.I.; et al. Somatic Sex Reprogramming of Adult Ovaries to Testes by FOXL2 Ablation. Cell 2009, 139, 1130–1142. [Google Scholar] [CrossRef] [Green Version]
- Pailhoux, E.; Parma, P.; Sundström, J.; Vigier, B.; Servel, N.; Kuopio, T.; Locatelli, A.; Pelliniemi, L.J.; Cotinot, C. Time course of female-to-male sex reversal in 38,XX fetal and postnatal pigs. Dev. Dyn. 2001, 222, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Matzuk, M.M.; Lamb, D.J. The biology of infertility: Research advances and clinical challenges. Nat. Med. 2008, 14, 1197–1213. [Google Scholar] [CrossRef]
- Schmidt, D.; Ovitt, C.E.; Anlag, K.; Fehsenfeld, S.; Gredsted, L.; Treier, A.-C.; Treier, M. The murine winged-helix transcription factor Foxl2 is required for granulosa cell differentiation and ovary maintenance. Development 2004, 131, 933–942. [Google Scholar] [CrossRef] [Green Version]
- Uda, M.; Ottolenghi, C.; Crisponi, L.; Garcia, J.E.; Deiana, M.; Kimber, W.; Forabosco, A.; Cao, A.; Schlessinger, D.; Pilia, G. Foxl2 disruption causes mouse ovarian failure by pervasive blockage of follicle development. Hum. Mol. Genet. 2004, 13, 1171–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowles, J.; Knight, D.; Smith, C.; Wilhelm, D.; Richman, J.; Mamiya, S.; Yashiro, K.; Chawengsaksophak, K.; Wilson, M.; Rossant, J.; et al. Retinoid Signaling Determines Germ Cell Fate in Mice. Science 2006, 312, 596–600. [Google Scholar] [CrossRef] [Green Version]
- Anderson, E.L.; Baltus, A.E.; Roepers-Gajadien, H.L.; Hassold, T.J.; de Rooij, D.; van Pelt, A.; Page, D.C. Stra8 and its inducer, retinoic acid, regulate meiotic initiation in both spermatogenesis and oogenesis in mice. Proc. Natl. Acad. Sci. USA 2008, 105, 14976–14980. [Google Scholar] [CrossRef] [Green Version]
- De Rooij, D.G.; Russell, L.D. All you wanted to know about spermatogonia but were afraid to ask. J. Androl. 2000, 21, 776–798. [Google Scholar]
- Goto, T.; Adjaye, J.; Rodeck, C.H.; Monk, M. Identification of genes expressed in human primordial germ cells at the time of entry of the female germ line into meiosis. Mol. Hum. Reprod. 1999, 5, 851–860. [Google Scholar] [CrossRef]
- McLaren, A. Meiosis and differentiation of mouse germ cells. Symp. Soc. Exp. Biol. 1984, 38, 7–23. [Google Scholar]
- MacLennan, M.; Crichton, J.H.; Playfoot, C.J.; Adams, I.R. Oocyte development, meiosis and aneuploidy. Semin. Cell Dev. Biol. 2015, 45, 68–76. [Google Scholar] [CrossRef] [Green Version]
- Gu, L.; Liu, H.; Gu, X.; Boots, C.; Moley, K.H.; Wang, Q. Metabolic control of oocyte development: Linking maternal nutrition and reproductive outcomes. Cell. Mol. Life Sci. 2015, 72, 251–271. [Google Scholar] [CrossRef] [PubMed]
- Ohinata, Y.; Ohta, H.; Shigeta, M.; Yamanaka, K.; Wakayama, T.; Saitou, M. A Signaling Principle for the Specification of the Germ Cell Lineage in Mice. Cell 2009, 137, 571–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eguizabal, C.; Shovlin, T.C.; Durcova-Hills, G.; Surani, A.; McLaren, A. Generation of primordial germ cells from pluripotent stem cells. Difference 2009, 78, 116–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toyooka, Y.; Tsunekawa, N.; Akasu, R.; Noce, T. Embryonic stem cells can form germ cells in vitro. Proc. Natl. Acad. Sci. USA 2003, 100, 11457–11462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kee, K.; Gonsalves, J.M.; Clark, A.T.; Pera, R.R. Bone Morphogenetic Proteins Induce Germ Cell Differentiation from Human Embryonic Stem Cells. Stem Cells Dev. 2006, 15, 831–837. [Google Scholar] [CrossRef]
- Geijsen, N.; Horoschak, M.; Kim, K.; Gribnau, J.; Eggan, K.; Daley, G.Q. Derivation of embryonic germ cells and male gametes from embryonic stem cells. Nat. Cell Biol. 2003, 427, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Ying, Q.-L.; Wray, J.; Nichols, J.; Batlle-Morera, L.; Doble, B.; Woodgett, J.; Cohen, P.; Smith, A. The ground state of embryonic stem cell self-renewal. Nat. Cell Biol. 2008, 453, 519–523. [Google Scholar] [CrossRef] [Green Version]
- Nichols, J.; Jones, K.; Phillips, J.M.; Newland, S.A.; Roode, M.; Mansfield, W.; Smith, A.; Cooke, A. Validated germline-competent embryonic stem cell lines from nonobese diabetic mice. Nat. Med. 2009, 15, 814–818. [Google Scholar] [CrossRef]
- Marks, H.; Kalkan, T.; Menafra, R.; Denissov, S.; Jones, K.; Hofemeister, H.; Nichols, J.; Kranz, A.; Stewart, A.F.; Smith, A.; et al. The Transcriptional and Epigenomic Foundations of Ground State Pluripotency. Cell 2012, 149, 590–604. [Google Scholar] [CrossRef] [Green Version]
- Chuma, S.; Kanatsu-Shinohara, M.; Inoue, K.; Ogonuki, N.; Miki, H.; Toyokuni, S.; Hosokawa, M.; Nakatsuji, N.; Ogura, A.; Shinohara, T. Spermatogenesis from epiblast and primordial germ cells following transplantation into postnatal mouse testis. Development 2005, 132, 117–122. [Google Scholar] [CrossRef] [Green Version]
- Gkountela, S.; Li, Z.; Vincent, J.J.; Zhang, K.X.; Chen, A.; Pellegrini, M.; Clark, A.T. The ontogeny of cKIT+ human primordial germ cells proves to be a resource for human germ line reprogramming, imprint erasure and in vitro differentiation. Nat. Cell Biol. 2013, 15, 113–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kee, K.; Angeles, V.T.; Flores, M.; Nguyen, H.N.; Pera, R.R. Human DAZL, DAZ and BOULE genes modulate primordial germ-cell and haploid gamete formation. Nat. Cell Biol. 2009, 462, 222–225. [Google Scholar] [CrossRef] [PubMed]
- Irie, N.; Weinberger, L.; Tang, W.W.; Kobayashi, T.; Viukov, S.; Manor, Y.S.; Dietmann, S.; Hanna, J.H.; Surani, M.A. SOX17 Is a Critical Specifier of Human Primordial Germ Cell Fate. Cell 2015, 160, 253–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, K.; Yokobayashi, S.; Nakamura, T.; Okamoto, I.; Yabuta, Y.; Kurimoto, K.; Ohta, H.; Moritoki, Y.; Iwatani, C.; Tsuchiya, H.; et al. Robust In Vitro Induction of Human Germ Cell Fate from Pluripotent Stem Cells. Cell Stem Cell 2015, 17, 178–194. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.C.; Wu, F.-C.; Wu, C.-E.; Chow, L.-P.; Ho, H.-N.; Chen, H.-F. Human pluripotent stem cell-derived DDX4 and KRT-8 positive cells participate in ovarian follicle-like structure formation. iScience 2021, 24, 102003. [Google Scholar] [CrossRef] [PubMed]
- Eckert, D.; Biermann, K.; Nettersheim, D.; Gillis, A.J.; Steger, K.; Jäck, H.-M.; Müller, A.M.; Looijenga, L.H.; Schorle, H. Expression of BLIMP1/PRMT5 and concurrent histone H2A/H4 arginine 3 dimethylation in fetal germ cells, CIS/IGCNU and germ cell tumors. BMC Dev. Biol. 2008, 8, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauls, K.; Jäger, R.; Weber, S.; Wardelmann, E.; Koch, A.; Büttner, R.; Schorle, H. Transcription factor AP-2γ, a novel marker of gonocytes and seminomatous germ cell tumors. Int. J. Cancer 2005, 115, 470–477. [Google Scholar] [CrossRef]
- Mishra, S.; Taelman, J.; Popovic, M.; Tilleman, L.; Duthoo, E.; van der Jeught, M.; Deforce, D.; van Nieuwerburgh, F.; Menten, B.; de Sutter, P.; et al. Activin A-derived human embryonic stem cells show increased competence to differentiate into primordial germ cell-like cells. Stem Cells 2021, 39, 551–563. [Google Scholar] [CrossRef]
- Li, Z.; Fang, F.; Zhao, Q.; Li, H.; Xiong, C. Supplementation of vitamin C promotes early germ cell specification from human embryonic stem cells. Stem Cell Res. Ther. 2019, 10, 324–329. [Google Scholar] [CrossRef]
- Wang, J.; Zhai, Q.; Zhang, R.; Ge, W.; Liu, J.; Li, L.; Sun, Z.; De Felici, M.; Shen, W. Effects of activin A on the transcriptome of mouse oogenesis in vitro. J. Cell. Physiol. 2019, 234, 14339–14350. [Google Scholar] [CrossRef]
- Kanatsu-Shinohara, M.; Ogonuki, N.; Inoue, K.; Miki, H.; Ogura, A.; Toyokuni, S.; Shinohara, T. Long-Term Proliferation in Culture and Germline Transmission of Mouse Male Germline Stem Cells. Biol. Reprod. 2003, 69, 612–616. [Google Scholar] [CrossRef] [Green Version]
- Kubota, H.; Avarbock, M.R.; Brinster, R.L. Growth factors essential for self-renewal and expansion of mouse spermatogonial stem cells. Proc. Natl. Acad. Sci. USA 2004, 101, 16489–16494. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.S.; Ryu, H.-S.; Jung, S.-E.; Shin, B.-J.; Won, J.-H.; Um, T.G.; Oh, H.; Kim, S.-H.; Ryu, S.-H.K.A.B.-Y. Expression profile of spermatogenesis associated genes in male germ cells during postnatal development in mice. J. Anim. Reprod. Biotechnol. 2020, 35, 289–296. [Google Scholar] [CrossRef]
- Western, P.S.; Miles, D.C.; Bergen, J.A.V.D.; Burton, M.; Sinclair, A. Dynamic Regulation of Mitotic Arrest in Fetal Male Germ Cells. Stem Cells 2008, 26, 339–347. [Google Scholar] [CrossRef]
- Nayernia, K.; Nolte, J.; Michelmann, H.W.; Lee, J.H.; Rathsack, K.; Drusenheimer, N.; Dev, A.; Wulf, G.; Ehrmann, I.E.; Elliott, D.; et al. In Vitro-Differentiated Embryonic Stem Cells Give Rise to Male Gametes that Can Generate Offspring Mice. Dev. Cell 2006, 11, 125–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikura, Y.; Yabuta, Y.; Ohta, H.; Hayashi, K.; Nakamura, T.; Okamoto, I.; Yamamoto, T.; Kurimoto, K.; Shirane, K.; Sasaki, H.; et al. In Vitro Derivation and Propagation of Spermatogonial Stem Cell Activity from Mouse Pluripotent Stem Cells. Cell Rep. 2016, 17, 2789–2804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, Y.S.; Suzuki, S.; Seita, Y.; Ito, J.; Sakata, Y.; Aso, H.; Sato, K.; Hermann, B.P.; Sasaki, K. Reconstitution of prospermatogonial specification in vitro from human induced pluripotent stem cells. Nat. Commun. 2020, 11, 1–17. [Google Scholar] [CrossRef]
- Yamashiro, C.; Sasaki, K.; Yabuta, Y.; Kojima, Y.; Nakamura, T.; Okamoto, I.; Yokobayashi, S.; Murase, Y.; Ishikura, Y.; Shirane, K.; et al. Generation of human oogonia from induced pluripotent stem cells in vitro. Science 2018, 362, 356–360. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.W.; Dietmann, S.; Irie, N.; Leitch, H.; Floros, V.I.; Bradshaw, C.; Hackett, J.; Chinnery, P.F.; Surani, M.A. A Unique Gene Regulatory Network Resets the Human Germline Epigenome for Development. Cell 2015, 161, 1453–1467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.; Wang, M.; Yuan, Y.; Wang, X.; Fu, R.; Wan, H.; Xie, M.; Liu, M.; Guo, X.; Zheng, Y.; et al. Complete Meiosis from Embryonic Stem Cell-Derived Germ Cells In Vitro. Cell Stem Cell 2016, 18, 330–340. [Google Scholar] [CrossRef] [Green Version]
- Pepling, M.E. From primordial germ cell to primordial follicle: Mammalian female germ cell development. Genesis 2006, 44, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Eppig, J.J. Development in vitro of mouse oocytes from primordial follicles. Biol. Reprod. 1996, 54, 197–207. [Google Scholar] [CrossRef]
- Gilchrist, R.B.; Luciano, A.M.; Richani, D.; Zeng, H.T.; Wang, X.; De Vos, M.; Sugimura, S.; Smitz, J.; Richard, F.J.; Thompson, J. Oocyte maturation and quality: Role of cyclic nucleotides. Reproduction. 2016, 152, R143–R157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirao, Y. Isolation of Ovarian Components Essential for Growth and Development of Mammalian Oocytes in Vitro. J. Reprod. Dev. 2012, 58, 167–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morohaku, K.; Tanimoto, R.; Sasaki, K.; Miki, R.; Kono, T.; Hayashi, K.; Hirao, Y.; Obata, Y. Complete in vitro generation of fertile oocytes from mouse primordial germ cells. Proc. Natl. Acad. Sci. USA 2016, 113, 9021–9026. [Google Scholar] [CrossRef] [Green Version]
- Lacham-Kaplan, O.; Chy, H.; Trounson, A. Testicular Cell Conditioned Medium Supports Differentiation of Embryonic Stem Cells into Ovarian Structures Containing Oocytes. Stem Cells 2006, 24, 266–273. [Google Scholar] [CrossRef]
- Hikabe, O.; Hamazaki, N.; Nagamatsu, G.; Obata, Y.; Hirao, Y.; Hamada, N.; Shimamoto, S.; Imamura, T.; Nakashima, K.; Saitou, M.; et al. Reconstitution in vitro of the entire cycle of the mouse female germ line. Nat. Cell Biol. 2016, 539, 299–303. [Google Scholar] [CrossRef]
- Hayashi, K.; Hikabe, O.; Obata, Y.; Hirao, Y. Reconstitution of mouse oogenesis in a dish from pluripotent stem cells. Nat. Protoc. 2017, 12, 1733–1744. [Google Scholar] [CrossRef]
- Ohta, H.; Kurimoto, K.; Okamoto, I.; Nakamura, T.; Yabuta, Y.; Miyauchi, H.; Yamamoto, T.; Okuno, Y.; Hagiwara, M.; Shirane, K.; et al. In vitro expansion of mouse primordial germ cell-like cells recapitulates an epigenetic blank slate. EMBO J. 2017, 36, 1888–1907. [Google Scholar] [CrossRef] [PubMed]
- Miyauchi, H.; Ohta, H.; Nagaoka, S.; Nakaki, F.; Sasaki, K.; Hayashi, K.; Yabuta, Y.; Nakamura, T.; Yamamoto, T.; Saitou, M. Bone morphogenetic protein and retinoic acid synergistically specify female germ-cell fate in mice. EMBO J. 2017, 36, 3100–3119. [Google Scholar] [CrossRef]
- Hamazaki, N.; Kyogoku, H.; Araki, H.; Miura, F.; Horikawa, C.; Hamada, N.; Shimamoto, S.; Hikabe, O.; Nakashima, K.; Kitajima, T.S.; et al. Reconstitution of the oocyte transcriptional network with transcription factors. Nat. Cell Biol. 2021, 589, 264–269. [Google Scholar] [CrossRef]
- Jung, D.; Xiong, J.; Ye, M.; Qin, X.; Li, L.; Cheng, S.; Luo, M.; Peng, J.; Dong, J.; Tang, F.; et al. In vitro differentiation of human embryonic stem cells into ovarian follicle-like cells. Nat. Commun. 2017, 8, 15680. [Google Scholar] [CrossRef]
- Harrison, S.E.; Sozen, B.; Christodoulou, N.; Kyprianou, C.; Zernicka-Goetz, M. Assembly of embryonic and extraembryonic stem cells to mimic embryogenesis in vitro. Science 2017, 356, eaal1810. [Google Scholar] [CrossRef] [Green Version]
- Sozen, B.; Amadei, G.; Cox, A.; Wang, R.; Na, E.; Czukiewska, S.; Chappell, L.; Voet, T.; Michel, G.; Jing, N.; et al. Self-assembly of embryonic and two extra-embryonic stem cell types into gastrulating embryo-like structures. Nat. Cell Biol. 2018, 20, 979–989. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, T.; Chen, N.; Gao, D.; Shi, B.; Kong, S.; West, R.; Yuan, Y.; Zhi, M.; Wei, Q.; et al. Implantation initiation of self-assembled embryo-like structures generated using three types of mouse blastocyst-derived stem cells. Nat. Commun. 2019, 10, 1–17. [Google Scholar] [CrossRef]
- Amadei, G.; Lau, K.Y.; De Jonghe, J.; Gantner, C.W.; Sozen, B.; Chan, C.; Zhu, M.; Kyprianou, C.; Hollfelder, F.; Zernicka-Goetz, M. Inducible Stem-Cell-Derived Embryos Capture Mouse Morphogenetic Events in Vitro. Dev. Cell 2021, 56, 366–382. [Google Scholar] [CrossRef]
- Liu, X.; Tan, J.P.; Schröder, J.; Aberkane, A.; Ouyang, J.F.; Mohenska, M.; Lim, S.M.; Sun, Y.B.Y.; Chen, J.; Sun, G.; et al. Modelling human blastocysts by reprogramming fibroblasts into iBlastoids. Nat. Cell Biol. 2021, 591, 627–632. [Google Scholar] [CrossRef]
- Brugo-Olmedo, S.; Chillik, C.; Kopelman, S. Definition and causes of infertility. Reprod. Biomed. Online 2001, 2, 41–53. [Google Scholar] [CrossRef]
- Zorrilla, M.; Yatsenko, A.N. The Genetics of Infertility: Current Status of the Field. Curr. Genet. Med. Rep. 2013, 1, 247–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, Y.; Saitou, M.; Yamanaka, S. Germline development from human pluripotent stem cells toward disease modeling of infertility. Fertil. Steril. 2012, 97, 1250–1259. [Google Scholar] [CrossRef] [PubMed]
- Chiba, K.; Enatsu, N.; Fujisawa, M. Management of non-obstructive azoospermia. Reprod. Med. Biol. 2016, 15, 165–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Ye, S.; Liang, D.; Wang, P.; Fu, J.; Ma, Q.; Kong, R.; Shi, L.; Gong, X.; Chen, W.; et al. In Vitro Modeling of Human Germ Cell Development Using Pluripotent Stem Cells. Stem Cell Rep. 2018, 10, 509–523. [Google Scholar] [CrossRef] [Green Version]
- Fang, F.; Li, Z.; Zhao, Q.; Ye, Z.; Gu, X.; Pan, F.; Li, H.; Xiang, W.; Xiong, C. Induced Pluripotent Stem Cells Derived From Two Idiopathic Azoospermia Patients Display Compromised Differentiation Potential for Primordial Germ Cell Fate. Front. Cell Dev. Biol. 2020, 8, 432. [Google Scholar] [CrossRef]
- Botman, O.; Hibaoui, Y.; Giudice, M.G.; Ambroise, J.; Creppe, C.; Feki, A.; Wyns, C. Modeling Klinefelter Syndrome Using Induced Pluripotent Stem Cells Reveals Impaired Germ Cell Differentiation. Front. Cell Dev. Biol. 2020, 8, 567454. [Google Scholar] [CrossRef] [PubMed]
- Van Assche, E.; Bonduelle, M.; Tournaye, H.; Joris, H.; Verheyen, G.; Devroey, P.; Van Steirteghem, A.; Liebaers, I. Cytogenetics of infertile men. Hum. Reprod. 1996, 11, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Leng, L.; Tan, Y.; Gong, F.; Hu, L.; Ouyang, Q.; Zhao, Y.; Lu, G.; Lin, G. Differentiation of primordial germ cells from induced pluripotent stem cells of primary ovarian insufficiency. Hum. Reprod. 2015, 30, 737–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilgner, K.; Atkinson, S.P.; Yung, S.; Golebiewska, A.; Stojkovic, M.; Moreno, R.; Lako, M.; Armstrong, L. Expression of GFP Under the Control of the RNA HelicaseVASAPermits FACS Isolation of Human Primordial Germ Cells. Stem Cells 2009, 28, 84–92. [Google Scholar] [CrossRef]
- Rao, M. Scalable human ES culture for therapeutic use: Propagation, differentiation, genetic modification and regulatory issues. Gene Ther. 2007, 15, 82–88. [Google Scholar] [CrossRef]
- Vitale, I.; Manic, G.; De Maria, R.; Kroemer, G.; Galluzzi, L. DNA Damage in Stem Cells. Mol. Cell 2017, 66, 306–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simara, P.; Tesarova, L.; Rehakova, D.; Matula, P.; Stejskal, S.; Hampl, A.; Koutna, I. DNA double-strand breaks in human induced pluripotent stem cell reprogramming and long-term in vitro culturing. Stem Cell Res. Ther. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Stefano, B.; Ueda, M.; Sabri, S.; Brumbaugh, J.; Huebner, A.J.; Sahakyan, A.; Clement, K.; Clowers, K.J.; Erickson, A.R.; Shioda, K.; et al. Reduced MEK inhibition preserves genomic stability in naive human embryonic stem cells. Nat. Methods 2018, 15, 732–740. [Google Scholar] [CrossRef]
- García-Rodríguez, A.; Gosálvez, J.; Agarwal, A.; Roy, R.; Johnston, S. DNA Damage and Repair in Human Reproductive Cells. Int. J. Mol. Sci. 2018, 20, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reik, W.; Surani, M.A. Germline and Pluripotent Stem Cells. Cold Spring Harb. Perspect. Biol. 2015, 7, a019422. [Google Scholar] [CrossRef] [Green Version]
- DeBaun, M.R.; Niemitz, E.L.; Feinberg, A.P. Association of In Vitro Fertilization with Beckwith-Wiedemann Syndrome and Epigenetic Alterations of LIT1 and H19. Am. J. Hum. Genet. 2003, 72, 156–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soejima, H.; Higashimoto, K. Epigenetic and genetic alterations of the imprinting disorder Beckwith–Wiedemann syndrome and related disorders. J. Hum. Genet. 2013, 58, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Meyer, E.; Lim, D.; Pasha, S.; Tee, L.J.; Rahman, F.; Yates, J.R.W.; Woods, C.G.; Reik, W.; Maher, E.R. Germline Mutation in NLRP2 (NALP2) in a Familial Imprinting Disorder (Beckwith-Wiedemann Syndrome). PLoS Genet. 2009, 5, e1000423. [Google Scholar] [CrossRef] [Green Version]
- Reik, W.; W.Brown, K.; E.Slatter, R.; Sartor, P.; Elliott, M.; R.Maher, E. Allelic methylation of H19 and IGF2 in the Beckwith—Wiedemann syndrome. Hum. Mol. Genet. 1994, 3, 1297–1301. [Google Scholar] [CrossRef]
- Monk, D.; Mackay, D.J.; Eggermann, T.; Maher, E.R.; Riccio, A. Genomic imprinting disorders: Lessons on how genome, epigenome and environment interact. Nat. Rev. Genet. 2019, 20, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Maher, E.R.; Brueton, L.A.; Bowdin, S.C.; Luharia, A.; Cooper, W.; Cole, T.R.; Macdonald, F.; Sampson, J.R.; Barratt, C.; Reik, W.; et al. Beckwith-Wiedemann syndrome and assisted reproduction technology (ART). J. Med Genet. 2003, 40, 62–64. [Google Scholar] [CrossRef] [Green Version]
- Ilic, D.; Ogilvie, C.; Noli, L.; Kolundzic, N.; Khalaf, Y. Human embryos from induced pluripotent stem cell-derived gametes: Ethical and quality considerations. Regen. Med. 2017, 12, 681–691. [Google Scholar] [CrossRef]
- Ishii, T.; Pera, R.A.R.; Greely, H.T. Ethical and Legal Issues Arising in Research on Inducing Human Germ Cells from Pluripotent Stem Cells. Cell Stem Cell 2013, 13, 145–148. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, M.; Kawaguchi, T.; Durcova-Hills, G.; Imai, H. Generation of germ cells from pluripotent stem cells in mammals. Reprod. Med. Biol. 2017, 17, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Horisawa, K.; Suzuki, A. Direct cell-fate conversion of somatic cells: Toward regenerative medicine and industries. Proc. Jpn. Acad. Ser. B 2020, 96, 131–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medrano, J.V.; Martínez-Arroyo, A.M.; Míguez, J.M.; Moreno, I.; Martínez, S.; Quiñonero, A.; Gimeno, P.D.; Marqués-Marí, A.I.; Pellicer, A.; Remohí, J.; et al. Human somatic cells subjected to genetic induction with six germ line-related factors display meiotic germ cell-like features. Sci. Rep. 2016, 6, 24956. [Google Scholar] [CrossRef] [Green Version]
- Aramaki, S.; Kagiwada, S.; Wu, G.; Obridge, D.; Adachi, K.; Kutejova, E.; Lickert, H.; Hübner, K.; Schöler, H.R. Residual pluripotency is required for inductive germ cell segregation. EMBO Rep. 2021, e52553, e52553. [Google Scholar] [CrossRef]
- Yu, L.; Wei, Y.; Sun, H.-X.; Mahdi, A.K.; Arteaga, C.A.P.; Sakurai, M.; Schmitz, D.A.; Zheng, C.; Ballard, E.D.; Li, J.; et al. Derivation of Intermediate Pluripotent Stem Cells Amenable to Primordial Germ Cell Specification. Cell Stem Cell 2021, 28, 550–567.e12. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T. Human iPS Cell-Derived Germ Cells: Current Status and Clinical Potential. J. Clin. Med. 2014, 3, 1064–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa-Borges, N.; Spath, K.; Miguel-Escalada, I.; Mestres, E.; Balmaseda, R.; Serafín, A.; Garcia-Jiménez, M.; Vanrell, I.; González, J.; Rink, K.; et al. Maternal spindle transfer overcomes embryo developmental arrest caused by ooplasmic defects in mice. eLife 2020, 9. [Google Scholar] [CrossRef]
- Labarta, E.; Santos, M.J.D.L.; Escribá, M.J.; Pellicer, A.; Herraiz, S. Mitochondria as a tool for oocyte rejuvenation. Fertil. Steril. 2019, 111, 219–226. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, T.-K.; Song, J.-H.; Lee, S.-B.; Do, J.-T. Germ Cell Derivation from Pluripotent Stem Cells for Understanding In Vitro Gametogenesis. Cells 2021, 10, 1889. https://doi.org/10.3390/cells10081889
Hong T-K, Song J-H, Lee S-B, Do J-T. Germ Cell Derivation from Pluripotent Stem Cells for Understanding In Vitro Gametogenesis. Cells. 2021; 10(8):1889. https://doi.org/10.3390/cells10081889
Chicago/Turabian StyleHong, Tae-Kyung, Jae-Hoon Song, So-Been Lee, and Jeong-Tae Do. 2021. "Germ Cell Derivation from Pluripotent Stem Cells for Understanding In Vitro Gametogenesis" Cells 10, no. 8: 1889. https://doi.org/10.3390/cells10081889