
Genetic Constructs for the Control of Astrocytes’ Activity

 , ,
, ,
Abstract
:1. Introduction
2. Astroglial Molecular Physiology
2.1. Heterogeneity of Astroglia
- (i)
- radial astroglia (GLAST+(–)/GFAP(+)(–)/Pax6+/FABP7+/Nestin+/Vimentin+/Pax6+/Cx43+/Cx30 cells) originating from neuroepithelial cells that are involved in embryonic and adult neurogenesis;
- (ii)
- protoplasmic astrocytes (GFAP++/s100β+/EAAT+/AQP4+/NDRG2+/Cx43/CD38+), which are the principal glial constituents of the neurovascular unit; they stay close to neurons due to direct contacts made by their perisynaptic processes and control neuronal excitability, plasticity, metabolic status, and close to brain microvessel endothelial cells due to their end-feet contacts to adjust local microcirculation to the actual needs of neurons;
- (iii)
- fibrous astrocytes (GFAP+(–)/CD44+ cells) surrounding myelinated fibers and controlling myelinization; iv) reactive astrocytes with upregulated expression of GFAP (GFAP++/EAAT+/Nestin+/Vimentin+/PDGFR+/Musashi+/CD44+/CD38+/Lcn-2+ cells) that take part in the progression of local inflammation and gliosis [11,12,13,14,15].
2.2. Measuring Astroglial Activity
- (i)
- rise in intracellular Ca2+ concentrations due to Ca2+ release from intracellular stores, Ca2+ entry, or connexon-mediated propagation of Ca2+ waves [25];
- (ii)
- metabolic changes in astrocytes (lactate production, K+ uptake, mitochondrial activity, etc.) caused by their activation [26];
- (iii)
- changes in the expression pattern in activated glial cells, i.e., due to conversion of resting astrocytes into reactive ones [27];
- (iv)
- changes in astroglial morphology [28];
- (v)
- and changes in astroglial secretory activity resulting in the release of gliotransmitters, cytokines, and growth factors [29].
2.3. Choice of Genetic Constructs Suitable for Glia Activation
3. Targeted Delivery of Genetic Constructs into Glial Cells
3.1. Is the Optimal AAV Serotype Myth or Reality?
3.2. General Requirements for the AAV Cassette for Efficient Transgene Expression
3.3. GFAP as a Specific Marker of Astroglial Cells
3.4. Efficient and Restricted AAV-Mediated Transgene Expression in Astrocytes: Does the Promoter Really Matter?
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siracusa, R.; Fusco, R.; Cuzzocrea, S. Astrocytes: Role and Functions in Brain Pathologies. Front. Pharmacol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oshio, K.; Watanabe, H.; Song, Y.; Verkman, A.S.; Manley, G.T. Reduced cerebrospinal fluid production and intracranial pressure in mice lacking choroid plexus water channel Aquaporin-1. FASEB J. 2005, 19, 76–78. [Google Scholar] [CrossRef]
- Witthoft, A.; Filosa, J.A.; Karniadakis, G.E. Potassium buffering in the neurovascular unit: Models and sensitivity analysis. Biophys. J. 2013, 105, 2046–2054. [Google Scholar] [CrossRef] [Green Version]
- Verkhratsky, A.; Zorec, R.; Parpura, V. Stratification of astrocytes in healthy and diseased brain. Brain Pathol. 2017, 27, 629–644. [Google Scholar] [CrossRef]
- Cabezas, R.; Avila, M.; Gonzalez, J.; El-Bachá, R.S.; Báez, E.; García-Segura, L.M.; Jurado Coronel, J.C.; Capani, F.; Cardona-Gomez, G.P.; Barreto, G.E. Astrocytic modulation of blood brain barrier: Perspectives on Parkinson’s disease. Front. Cell. Neurosci. 2014, 8, 211. [Google Scholar] [CrossRef] [Green Version]
- Kitchen, P.; Salman, M.M.; Halsey, A.M.; Clarke-Bland, C.; MacDonald, J.A.; Ishida, H.; Vogel, H.J.; Almutiri, S.; Logan, A.; Kreida, S.; et al. Targeting Aquaporin-4 Subcellular Localization to Treat Central Nervous System Edema. Cell 2020, 181, 784–799. [Google Scholar] [CrossRef]
- Salmina, A.B.; Gorina, Y.V.; Erofeev, A.I.; Balaban, P.M.; Bezprozvanny, I.B.; Vlasova, O.L. Optogenetic and chemogenetic modulation of astroglial secretory phenotype. Rev. Neurosci. 2021. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Kang, X.; Hu, J.; Zhang, D.; Liang, Z.; Meng, F.; Zhang, X.; Xue, Y.; Maimon, R.; Dowdy, S.F.; et al. Reversing a model of Parkinson‘s disease with in situ converted nigral neurons. Nature 2020, 582, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Sylvain, N.J.; Salman, M.M.; Pushie, M.J.; Hou, H.; Meher, V.; Herlo, R.; Peeling, L.; Kelly, M.E. The effects of trifluoperazine on brain edema, aquaporin-4 expression and metabolic markers during the acute phase of stroke using photothrombotic mouse model. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183573. [Google Scholar] [CrossRef]
- Westergard, T.; Rothstein, J.D. Astrocyte Diversity: Current Insights and Future Directions. Neurochem. Res. 2020, 45, 1298–1305. [Google Scholar] [CrossRef]
- Zhang, Z.; Ma, Z.; Zou, W.; Guo, H.; Liu, M.; Ma, Y.; Zhang, L. The Appropriate Marker for Astrocytes: Comparing the Distribution and Expression of Three Astrocytic Markers in Different Mouse Cerebral Regions. BioMed Res. Int. 2019, 2019, 9605265. [Google Scholar] [CrossRef]
- Morgun, A.V.; Malinovskaya, N.A.; Komleva, Y.K.; Lopatina, O.L.; Kuvacheva, N.V.; Panina, Y.A.; Taranushenko, T.Y.; Solonchuk, Y.R.; Salmina, A.B. Structural and functional heterogeneity of astrocytes in the brain: Role in neurodegeneration and neuroinflammation. Bull. Sib. Med. 2014, 13, 138–148. [Google Scholar] [CrossRef]
- Escartin, C.; Galea, E.; Lakatos, A.; O’Callaghan, J.P.; Petzold, G.C.; Serrano-Pozo, A.; Steinhäuser, C.; Volterra, A.; Carmignoto, G.; Agarwal, A.; et al. Reactive astrocyte nomenclature, definitions, and future directions. Nat. Neurosci. 2021, 24, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Aboufares El Alaoui, A.; Jackson, M.; Fabri, M.; de Vivo, L.; Bellesi, M. Characterization of Subcellular Organelles in Cortical Perisynaptic Astrocytes. Front. Cell. Neurosci. 2021, 14. [Google Scholar] [CrossRef] [PubMed]
- Girgrah, N.; Letarte, M.; Becker, L.E.; Cruz, T.F.; Theriault, E.; Moscarello, M.A. Localization of the CD44 glycoprotein to fibrous astrocytes in normal white matter and to reactive astrocytes in active lesions in multiple sclerosis. J. Neuropathol. Exp. Neurol. 1991, 50, 779–792. [Google Scholar] [CrossRef]
- Matias, I.; Morgado, J.; Gomes, F.C.A. Astrocyte Heterogeneity: Impact to Brain Aging and Disease. Front. Aging Neurosci. 2019, 11. [Google Scholar] [CrossRef] [Green Version]
- Chai, H.; Diaz-Castro, B.; Shigetomi, E.; Monte, E.; Octeau, J.C.; Yu, X.; Cohn, W.; Rajendran, P.S.; Vondriska, T.M.; Whitelegge, J.P.; et al. Neural Circuit-Specialized Astrocytes: Transcriptomic, Proteomic, Morphological, and Functional Evidence. Neuron 2017, 95, 531–549. [Google Scholar] [CrossRef]
- Vasile, F.; Dossi, E.; Rouach, N. Human astrocytes: Structure and functions in the healthy brain. Brain Struct. Funct. 2017, 222, 2017–2029. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Jakobs, T.C. Structural remodeling of astrocytes in the injured CNS. Neuroscientist 2012, 18, 567–588. [Google Scholar] [CrossRef] [Green Version]
- Gundersen, G.A.; Vindedal, G.F.; Skare, O.; Nagelhus, E.A. Evidence that pericytes regulate aquaporin-4 polarization in mouse cortical astrocytes. Brain Struct. Funct. 2014, 219, 2181–2186. [Google Scholar] [CrossRef] [Green Version]
- Wevers, N.R.; Kasi, D.G.; Gray, T.; Wilschut, K.J.; Smith, B.; van Vught, R.; Shimizu, F.; Sano, Y.; Kanda, T.; Marsh, G.; et al. A perfused human blood-brain barrier on-a-chip for high-throughput assessment of barrier function and antibody transport. Fluids Barriers CNS 2018, 15, 23. [Google Scholar] [CrossRef] [Green Version]
- Salman, M.M.; Marsh, G.; Kusters, I.; Delincé, M.; Di Caprio, G.; Upadhyayula, S.; de Nola, G.; Hunt, R.; Ohashi, K.G.; Gray, T.; et al. Design and Validation of a Human Brain Endothelial Microvessel-on-a-Chip Open Microfluidic Model Enabling Advanced Optical Imaging. Front. Bioeng. Biotechnol. 2020, 8, 573775. [Google Scholar] [CrossRef]
- Wang, S.N.; Wang, Z.; Xu, T.Y.; Cheng, M.H.; Li, W.L.; Miao, C.Y. Cerebral Organoids Repair Ischemic Stroke Brain Injury. Transl. Stroke Res. 2020, 11, 983–1000. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.J. Astrocyte Heterogeneity in the Adult Central Nervous System. Front. Cell. Neurosci. 2018, 12. [Google Scholar] [CrossRef] [Green Version]
- Guerra-Gomes, S.; Sousa, N.; Pinto, L.; Oliveira, J.F. Functional Roles of Astrocyte Calcium Elevations: From Synapses to Behavior. Front. Cell. Neurosci. 2018, 11. [Google Scholar] [CrossRef] [Green Version]
- Vardjan, N.; Chowdhury, H.H.; Horvat, A.; Velebit, J.; Malnar, M.; Muhič, M.; Kreft, M.; Krivec, Š.G.; Bobnar, S.T.; Miš, K.; et al. Enhancement of Astroglial Aerobic Glycolysis by Extracellular Lactate-Mediated Increase in cAMP. Front. Mol. Neurosci. 2018, 11. [Google Scholar] [CrossRef]
- Hyvärinen, T.; Hagman, S.; Ristola, M.; Sukki, L.; Veijula, K.; Kreutzer, J.; Kallio, P.; Narkilahti, S. Co-stimulation with IL-1β and TNF-α induces an inflammatory reactive astrocyte phenotype with neurosupportive characteristics in a human pluripotent stem cell model system. Sci. Rep. 2019, 9, 16944. [Google Scholar] [CrossRef]
- Acaz-Fonseca, E.; Ortiz-Rodriguez, A.; Azcoitia, I.; Garcia-Segura, L.M.; Arevalo, M.-A. Notch signaling in astrocytes mediates their morphological response to an inflammatory challenge. Cell Death Dis. 2019, 5, 85. [Google Scholar] [CrossRef]
- Osipova, E.D.; Semyachkina-Glushkovskaya, O.V.; Morgun, A.V.; Pisareva, N.V.; Malinovskaya, N.A.; Boitsova, E.B.; Pozhilenkova, E.A.; Belova, O.A.; Salmin, V.V.; Taranushenko, T.E.; et al. Gliotransmitters and cytokines in the control of blood-brain barrier permeability. Rev. Neurosci. 2018, 29, 567–591. [Google Scholar] [CrossRef] [Green Version]
- Bordone, M.P.; Salman, M.M.; Titus, H.E.; Amini, E.; Andersen, J.V.; Chakraborti, B.; Diuba, A.V.; Dubouskaya, T.G.; Ehrke, E.; Espindola de Freitas, A.; et al. The energetic brain—A review from students to students. J. Neurochem. 2019, 151, 139–165. [Google Scholar] [CrossRef]
- Pellerin, L.; Magistretti, P.J. Glutamate uptake into astrocytes stimulates aerobic glycolysis: A mechanism coupling neuronal activity to glucose utilization. Proc. Natl. Acad. Sci. USA 1994, 91, 10625–10629. [Google Scholar] [CrossRef] [Green Version]
- Salmina, A.B.; Kuvacheva, N.V.; Morgun, A.V.; Komleva, Y.K.; Pozhilenkova, E.A.; Lopatina, O.L.; Gorina, Y.V.; Taranushenko, T.E.; Petrova, L.L. Glycolysis-mediated control of blood-brain barrier development and function. Int. J. Biochem. Cell Biol. 2015, 64, 174–184. [Google Scholar] [CrossRef]
- Bak, L.K.; Walls, A.B.; Schousboe, A.; Ring, A.; Sonnewald, U.; Waagepetersen, H.S. Neuronal glucose but not lactate utilization is positively correlated with NMDA-induced neurotransmission and fluctuations in cytosolic Ca2+ levels. J. Neurochem. 2009, 109, 87–93. [Google Scholar] [CrossRef]
- Toth, A.B.; Hori, K.; Novakovic, M.M.; Bernstein, N.G.; Lambot, L.; Prakriya, M. CRAC channels regulate astrocyte Ca2+ signaling and gliotransmitter release to modulate hippocampal GABAergic transmission. Sci. Signal. 2019, 12, eaaw5450. [Google Scholar] [CrossRef]
- Parpura, V.; Grubišić, V.; Verkhratsky, A. Ca2+ sources for the exocytotic release of glutamate from astrocytes. Biochim. Biophys. Acta Mol. Cell Res. 2011, 1813, 984–991. [Google Scholar] [CrossRef]
- Shibasaki, K.; Ikenaka, K.; Tamalu, F.; Tominaga, M.; Ishizaki, Y. A Novel Subtype of Astrocytes Expressing TRPV4 (Transient Receptor Potential Vanilloid 4) Regulates Neuronal Excitability via Release of Gliotransmitters. J. Biol. Chem. 2014, 289, 14470–14480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiacco, T.A.; McCarthy, K.D. Multiple Lines of Evidence Indicate That Gliotransmission Does Not Occur under Physiological Conditions. J. Neurosci. 2018, 38, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Edling, Y.; Ingelman-Sundberg, M.; Simi, A. Glutamate activates c-fos in glial cells via a novel mechanism involving the glutamate receptor subtype mGlu5 and the transcriptional repressor DREAM. Glia 2007, 55, 328–340. [Google Scholar] [CrossRef]
- Rodríguez, J.J.; Davies, H.A.; Errington, M.L.; Verkhratsky, A.; Bliss, T.V.P.; Stewart, M.G. ARG3.1/ARC expression in hippocampal dentate gyrus astrocytes: Ultrastructural evidence and co-localization with glial fibrillary acidic protein. J. Cell. Mol. Med. 2008, 12, 671–678. [Google Scholar] [CrossRef] [Green Version]
- Franceschini, A.; Costantini, I.; Pavone, F.S.; Silvestri, L. Dissecting Neuronal Activation on a Brain-Wide Scale with Immediate Early Genes. Front. Neurosci. 2020, 14. [Google Scholar] [CrossRef] [PubMed]
- Groves, A.; Kihara, Y.; Jonnalagadda, D.; Rivera, R.; Kennedy, G.; Mayford, M.; Chun, J.A. Functionally defined in vivo astrocyte population identified by c-Fos activation in a mouse model of multiple sclerosis modulated by S1P signaling: Immediate-early astrocytes (ieAstrocytes). eNeuro 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Ma, K.; Ding, X.; Song, Q.; Han, Z.; Yao, H.; Ding, J.; Hu, G. Lactate enhances Arc/arg3.1 expression through hydroxycarboxylic acid receptor 1-β-arrestin2 pathway in astrocytes. Neuropharmacology 2020, 171, 108084. [Google Scholar] [CrossRef]
- Descalzi, G.; Gao, V.; Steinman, M.Q.; Suzuki, A.; Alberini, C.M. Lactate from astrocytes fuels learning-induced mRNA translation in excitatory and inhibitory neurons. Commun. Biol. 2019, 2, 247. [Google Scholar] [CrossRef]
- Jiang, Y.; VanDongen, A.M.J. Neuronal Activity-Dependent Accumulation of Arc in Astrocytes. bioRxiv 2020. [Google Scholar] [CrossRef]
- Adamsky, A.; Kol, A.; Kreisel, T.; Doron, A.; Ozeri-Engelhard, N.; Melcer, T.; Refaeli, R.; Horn, H.; Regev, L.; Groysman, M.; et al. Astrocytic activation generates de novo neuronal potentiation and memory enhancement. Cell 2018, 174, 59–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, L.; Chen, C.; Zheng, H.; Jin, L. The evolutionary relationship between microbial rhodopsins and metazoan rhodopsins. Sci. World J. 2013, 2013, 435651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Vierock, J.; Yizhar, O.; Fenno, L.E.; Tsunoda, S.; Kianianmomeni, A.; Prigge, M.; Berndt, A.; Cushman, J.; Polle, J.; et al. The microbial opsin family of optogenetic tools. Cell 2011, 147, 1446–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porter, M.L.; Blasic, J.R.; Bok, M.J.; Cameron, E.G.; Pringle, T.; Cronin, T.W.; Robinson, P.R. Shedding new light on opsin evolution. Proc. Biol. Sci. 2012, 279, 3–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Do, M.T.H. Melanopsin and the intrinsically photosensitive retinal ganglion cells: Biophysics to behavior. Neuron 2019, 104, 205–226. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, L.P.; Boyden, E.S.; Deisseroth, K. Channelrhodopsin-2 and optical control of excitable cells. Nat. Methods 2006, 3, 785–792. [Google Scholar] [CrossRef]
- Malyshev, A.Y.; Roshchin, M.V.; Smirnova, G.R.; Dolgikh, D.A.; Balaban, P.M.; Ostrovsky, M.A. Chloride conducting light activated channel GtACR2 can produce both cessation of firing and generation of action potentials in cortical neurons in response to light. Neurosci. Lett. 2017, 640, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Airan, R.D.; Thompson, K.R.; Fenno, L.E.; Bernstein, H.; Deisseroth, K. Temporally precise in vivo control of intracellular signalling. Nature 2009, 458, 1025–1029. [Google Scholar] [CrossRef] [PubMed]
- Covelo, A.; Araque, A. Neuronal activity determines distinct gliotransmitter release from a single astrocyte. Elife 2018, 7, e32237. [Google Scholar] [CrossRef]
- Mederos, S.; Hernández-Vivanco, A.; Ramírez-Franco, J.; Martín-Fernández, M.; Navarrete, M.; Yang, A.; Boyden, E.S.; Perea, G. Melanopsin for precise optogenetic activation of astrocyte-neuron networks. Glia 2019, 67, 915–934. [Google Scholar] [CrossRef]
- Caudal, L.C.; Gobbo, D.; Scheller, A.; Kirchhoff, F. The paradox of astroglial Ca2+ signals at the interface of excitation and inhibition. Front. Cell. Neurosci. 2020, 1, 609947. [Google Scholar] [CrossRef]
- Tan, Z.; Liu, Y.; Xi, W.; Lou, H.F.; Zhu, L.; Guo, Z.; Mei, L.; Duan, S. Glia-derived ATP inversely regulates excitability of pyramidal and CCK-positive neurons. Nat. Commun. 2017, 8, 13772. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Tan, Z.; Zeng, L.; Zhang, X.; He, Y.; Gao, W.; Wu, X.; Li, Y.; Bu, B.; Wang, W.; et al. Heterosynaptic long-term depression mediated by ATP released from astrocytes. Glia 2013, 61, 178–191. [Google Scholar] [CrossRef]
- Letellier, M.; Park, Y.K.; Chater, T.E.; Chipman, P.H.; Gautam, S.G.; Oshima-Takago, T.; Goda, Y. Astrocytes regulate heterogeneity of presynaptic strengths in hippocampal networks. Proc. Natl. Acad. Sci. USA 2016, 113, E2685–E2694. [Google Scholar] [CrossRef] [Green Version]
- Samulski, R.J.; Chang, L.S.; Shenk, T. Helper-free stocks of recombinant adeno-associated viruses: Normal integration does not require viral gene expression. J. Virol. 1989, 63, 3822–3828. [Google Scholar] [CrossRef] [Green Version]
- Ortinski, P.I.; Dong, J.; Mungenast, A.; Yue, C.; Takano, H.; Watson, D.J.; Haydon, P.G.; Coulter, D.A. Selective induction of astrocytic gliosis generates deficits in neuronal inhibition. Nat. Neurosci. 2010, 13, 584–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aschauer, D.F.; Kreuz, S.; Rumpel, S. Analysis of transduction efficiency, tropism and axonal transport of AAV serotypes 1, 2, 5, 6, 8 and 9 in the mouse brain. PLoS ONE 2013, 8, e76310. [Google Scholar] [CrossRef] [Green Version]
- Watakabe, A.; Ohtsuka, M.; Kinoshita, M.; Takaji, M.; Isa, K.; Mizukami, H.; Ozawa, K.; Isa, T.; Yamamori, T. Comparative analyses of adeno-associated viral vector serotypes 1, 2, 5, 8 and 9 in marmoset, mouse and macaque cerebral cortex. Neurosci. Res. 2015, 93, 144–157. [Google Scholar] [CrossRef] [Green Version]
- Schober, A.L.; Gagarkin, D.A.; Chen, Y.; Gao, G.; Jacobson, L.; Mongin, A.A. Recombinant adeno-associated virus serotype 6 (rAAV6) potently and preferentially transduces rat astrocytes in vitro and in vivo. Front. Cell. Neurosci. 2016, 10, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, R.; Janson, C.G.; Mastakov, M.; Lawlor, P.; Young, D.; Mouravlev, A.; Fitzsimons, H.; Choi, K.L.; Ma, H.; Dragunow, M.; et al. Quantitative comparison of expression with adeno-associated virus (AAV-2) brain-specific gene cassettes. Gene Ther. 2001, 8, 1323–1332. [Google Scholar] [CrossRef] [Green Version]
- Powell, S.K.; Samulski, R.J.; McCown, T.J. AAV Capsid-Promoter Interactions Determine CNS Cell-Selective Gene Expression In Vivo. Mol. Ther. 2020, 28, 1373–1380. [Google Scholar] [CrossRef] [PubMed]
- Lawlor, P.A.; Bland, R.J.; Mouravlev, A.; Young, D.; During, M.J. Efficient gene delivery and selective transduction of glial cells in the mammalian brain by AAV serotypes isolated from nonhuman primates. Mol. Ther. 2009, 17, 1692–1702. [Google Scholar] [CrossRef]
- Koerber, J.T.; Klimczak, R.; Jang, J.H.; Dalkara, D.; Flannery, J.G.; Schaffer, D.V. Molecular evolution of adeno-associated virus for enhanced glial gene delivery. Mol. Ther. 2009, 17, 2088–2095. [Google Scholar] [CrossRef]
- Howard, D.B.; Powers, K.; Wang, Y.; Harvey, B.K. Tropism and toxicity of adeno-associated viral vector serotypes 1, 2, 5, 6, 7, 8, and 9 in rat neurons and glia in vitro. Virology 2008, 372, 24–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foust, K.D.; Nurre, E.; Montgomery, C.L.; Hernandez, A.; Chan, C.M.; Kaspar, B.K. Intravascular AAV9 preferentially targets neonatal neurons and adult astrocytes. Nat. Biotechnol. 2009, 27, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakrabarty, P.; Rosario, A.; Cruz, P.; Siemienski, Z.; Ceballos-Diaz, C.; Crosby, K.; Jansen, K.; Borchelt, D.R.; Kim, J.Y.; Jankowsky, J.L.; et al. Capsid serotype and timing of injection determines AAV transduction in the neonatal mice brain. PLoS ONE 2013, 8, e67680. [Google Scholar] [CrossRef]
- Gray, S.J.; Blake, B.L.; Criswell, H.E.; Nicolson, S.C.; Samulski, R.J.; McCown, T.J.; Li, W. Directed evolution of a novel adeno-associated virus (AAV) vector that crosses the seizure-compromised blood-brain barrier (BBB). Mol. Ther. 2010, 18, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.Y.; Jang, M.J.; Yoo, B.B.; Greenbaum, A.; Ravi, N.; Wu, W.L.; Sánchez-Guardado, L.; Lois, C.; Mazmanian, S.K.; Deverman, B.E.; et al. Engineered AAVs for efficient noninvasive gene delivery to the central and peripheral nervous systems. Nat. Neurosci. 2017, 20, 1172–1179. [Google Scholar] [CrossRef]
- Bedbrook, C.N.; Deverman, B.E.; Gradinaru, V. Viral Strategies for Targeting the Central and Peripheral Nervous Systems. Annu. Rev. Neurosci. 2018, 41, 323–348. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhu, M.; Zhang, Y.; Diao, Y. Crossing the blood-brain barrier with AAV vectors. Metab. Brain Dis. 2021, 36, 45–52. [Google Scholar] [CrossRef]
- Huang, Q.; Chan, K.Y.; Tobey, I.G.; Chan, Y.A.; Poterba, T.; Boutros, C.L.; Balazs, A.B.; Daneman, R.; Bloom, J.M.; Seed, C.; et al. Delivering genes across the blood-brain barrier: LY6A, a novel cellular receptor for AAV-PHP.B capsids. PLoS ONE 2019, 14, e0225206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, B.S.; Wang, X.; Pan, Y.H.; Jiang, G.; Feng, J.F.; Lin, Y. Apolipoprotein E, low-density lipoprotein receptor, and immune cells control blood-brain barrier penetration by AAV-PHP.eB in mice. Theranostics 2021, 11, 1177–1191. [Google Scholar] [CrossRef]
- Loughner, C.L.; Bruford, E.A.; McAndrews, M.S.; Delp, E.E.; Swamynathan, S.; Swamynathan, S.K. Organization, evolution and functions of the human and mouse Ly6/uPAR family genes. Hum. Genom. 2016, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Gong, L.; Liu, F.; Mu, J. Stem cells and vascular dementia: From basic science to the clinic. Cell Tissue Banking 2020, 21, 349–360. [Google Scholar] [CrossRef]
- LaDu, M.J.; Shah, J.A.; Reardon, C.A.; Getz, G.S.; Bu, G.; Hu, J.; Guo, L.; van Eldik, L.J. Apolipoprotein E receptors mediate the effects of beta-amyloid on astrocyte cultures. J. Biol. Chem. 2000, 275, 33974–33980. [Google Scholar] [CrossRef] [Green Version]
- Klein, R.L.; Dayton, R.D.; Tatom, J.B.; Henderson, K.M.; Henning, P.P. AAV8, 9, Rh10, Rh43 vector gene transfer in the rat brain: Effects of serotype, promoter and purification method. Mol. Ther. 2008, 16, 89–96. [Google Scholar] [CrossRef]
- Dong, J.Y.; Fan, P.D.; Frizzell, R.A. Quantitative analysis of the packaging capacity of recombinant adeno-associated virus. Hum. Gene Ther. 1996, 7, 2101–2112. [Google Scholar] [CrossRef]
- Gray, S.J.; Samulski, R.J. Vector Design and Considerations for CNS Applications; University of North Carolina: Chapel Hill, NC, USA, 2011; pp. 9–15. [Google Scholar]
- Pfarr, D.S.; Rieser, L.A.; Woychik, R.P.; Rottman, F.M.; Rosenberg, M.; Reff, M.E. Differential effects of polyadenylation regions on gene expression in mammalian cells. DNA 1986, 5, 115–122. [Google Scholar] [CrossRef]
- Carswell, S.; Alwine, J.C. Efficiency of utilization of the simian virus 40 late polyadenylation site: Effects of upstream sequences. Mol. Cell. Biol. 1989, 9, 4248–4258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.M.; Yen, T.S. Hepatitis B virus RNA element that facilitates accumulation of surface gene transcripts in the cytoplasm. J. Virol. 1994, 68, 3193–3199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donello, J.E.; Loeb, J.E.; Hope, T.J. Woodchuck hepatitis virus contains a tripartite posttranscriptional regulatory element. J. Virol. 1998, 72, 5085–5092. [Google Scholar] [CrossRef] [Green Version]
- Loeb, J.E.; Cordier, W.S.; Harris, M.E.; Weitzman, M.D.; Hope, T.J. Enhanced expression of transgenes from adeno-associated virus vectors with the woodchuck hepatitis virus posttranscriptional regulatory element: Implications for gene therapy. Hum. Gene Ther. 1999, 10, 2295–2305. [Google Scholar] [CrossRef] [Green Version]
- Powell, S.K.; Rivera-Soto, R.; Gray, S.J. Viral expression cassette elements to enhance transgene target specificity and expression in gene therapy. Discov. Med. 2015, 19, 49–57. [Google Scholar]
- Paterna, J.C.; Moccetti, T.; Mura, A.; Feldon, J.; Büeler, H. Influence of promoter and WHV post-transcriptional regulatory element on AAV-mediated transgene expression in the rat brain. Gene Ther. 2000, 7, 1304–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.H.; Yu, N.K.; Baek, G.C.; Bakes, J.; Seo, D.; Nam, H.J.; Baek, S.H.; Lim, C.S.; Lee, Y.S.; Kaang, B.K. Optimization of AAV expression cassettes to improve packaging capacity and transgene expression in neurons. Mol. Brain 2014, 7, 17. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Wang, K.K. Glial fibrillary acidic protein: From intermediate filament assembly and gliosis to neurobiomarker. Trends Neurosci. 2015, 38, 364–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomsen, R.; Daugaard, T.F.; Holm, I.E.; Nielsen, A.L. Alternative mRNA Splicing from the Glial Fibrillary Acidic Protein (GFAP) Gene Generates Isoforms with Distinct Subcellular mRNA Localization Patterns in Astrocytes. PLoS ONE 2013, 8, e72110. [Google Scholar] [CrossRef]
- Hol, E.M.; Pekny, M. Glial fibrillary acidic protein (GFAP) and the astrocyte intermediate filament system in diseases of the central nervous system. Curr. Opin. Cell Biol. 2015, 32, 121–130. [Google Scholar] [CrossRef]
- Pekny, M.; Nilsson, M. Astrocyte activation and reactive gliosis. Glia 2005, 50, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Guo, Y.; Teng, J.; Ding, M.; Yu, A.C.H.; Chen, J. 14-3-3γ affects dynamics and integrity of glial filaments by binding to phosphorylated GFAP. J. Cell Sci. 2006, 119, 4452–4461. [Google Scholar] [CrossRef] [Green Version]
- Viedma-Poyatos, Á.; de Pablo, Y.; Pekny, M.; Pérez-Sala, D. The cysteine residue of glial fibrillary acidic protein is a critical target for lipoxidation and required for efficient network organization. Free Radic. Biol. Med. 2018, 120, 380–394. [Google Scholar] [CrossRef] [PubMed]
- Rodnight, R.; Goncalves, C.A.; Wofchuk, S.T.; Leal, R. Control of the phosphorylation of the astrocyte marker glial fibrillary acidic protein (GFAP) in the immature rat hippocampus by glutamate and calcium ions: Possible key factor in astrocytic plasticity. Braz. J. Med. Biol. Res. 1997, 30, 325–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walz, W.; Lang, M.K. Immunocytochemical evidence for a distinct GFAP-negative subpopulation of astrocytes in the adult rat hippocampus. Neurosci. Lett. 1998, 257, 127–130. [Google Scholar] [CrossRef]
- Lee, Y.; Messing, A.; Su, M.; Brenner, M. GFAP promoter elements required for region-specific and astrocyte-specific expression. Glia 2008, 56, 481–493. [Google Scholar] [CrossRef]
- Brenner, M.; Messing, A. Regulation of GFAP Expression. ASN Neuro 2021, 13. [Google Scholar] [CrossRef]
- Akdemir, E.S.; Huang, A.Y.-S.; Deneen, B. Astrocytogenesis: Where, when, and how. F1000Res 2020, 9, F1000. [Google Scholar] [CrossRef] [PubMed]
- Sousa Vde, O.; Romão, L.; Neto, V.M.; Gomes, F.C. Glial fibrillary acidic protein gene promoter is differently modulated by transforming growth factor-beta 1 in astrocytes from distinct brain regions. Eur. J. Neurosci. 2004, 19, 1721–1730. [Google Scholar] [CrossRef] [PubMed]
- Romão, L.F.; Sousa Vde, O.; Neto, V.M.; Gomes, F.C. Glutamate activates GFAP gene promoter from cultured astrocytes through TGF-beta1 pathways. J. Neurochem. 2008, 106, 746–756. [Google Scholar] [CrossRef] [PubMed]
- Kanski, R.; Sneeboer, M.A.; van Bodegraven, E.J.; Sluijs, J.A.; Kropff, W.; Vermunt, M.W.; Creyghton, M.P.; De Filippis, L.; Vescovi, A.; Aronica, E.; et al. Histone acetylation in astrocytes suppresses GFAP and stimulates a reorganization of the intermediate filament network. J. Cell Sci. 2014, 127, 4368–4380. [Google Scholar] [CrossRef] [Green Version]
- Roelofs, R.F.; Fischer, D.F.; Houtman, S.H.; Sluijs, J.A.; Van Haren, W.; Van Leeuwen, F.W.; Hol, E.M. Adult human subventricular, subgranular, and subpial zones contain astrocytes with a specialized intermediate filament cytoskeleton. Glia 2005, 52, 289–300. [Google Scholar] [CrossRef]
- Preman, P.; Alfonso-Triguero, M.; Alberdi, E.; Verkhratsky, A.; Arranz, A.M. Astrocytes in Alzheimer’s Disease: Pathological Significance and Molecular Pathways. Cells 2021, 10, 540. [Google Scholar] [CrossRef]
- Magnusson, J.P.; Zamboni, M.; Santopolo, G.; Mold, J.E.; Barrientos-Somarribas, M.; Talavera-Lopez, C.; Andersson, B.; Frisén, J. Activation of a neural stem cell transcriptional program in parenchymal astrocytes. eLife 2020, 9, e59733. [Google Scholar] [CrossRef] [PubMed]
- Leavitt, B.R.; Hernit-Grant, C.S.; Macklis, J.D. Mature astrocytes transform into transitional radial glia within adult mouse neocortex that supports directed migration of transplanted immature neurons. Exp. Neurol. 1999, 157, 43–57. [Google Scholar] [CrossRef] [Green Version]
- Docampo-Seara, A.; Santos-Durán, G.N.; Candal, E.; Rodríguez Díaz, M.Á. Expression of radial glial markers (GFAP, BLBP and GS) during telencephalic development in the catshark (Scyliorhinus canicula). Brain Struct. Funct. 2019, 224, 33–56. [Google Scholar] [CrossRef] [Green Version]
- Fox, I.J.; Paucar, A.A.; Nakano, I.; Mottahedeh, J.; Dougherty, J.D.; Kornblum, H.I. Developmental expression of glial fibrillary acidic protein mRNA in mouse forebrain germinal zones--implications for stem cell biology. Brain Res. Dev. Brain Res. 2004, 153, 121–125. [Google Scholar] [CrossRef]
- Middeldorp, J.; Boer, K.; Sluijs, J.A.; De Filippis, L.; Encha-Razavi, F.; Vescovi, A.L.; Swaab, D.F.; Aronica, E.; Hol, E.M. GFAPδ in radial glia and subventricular zone progenitors in the developing human cortex. Development 2010, 137, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Cipriani, S.; Ferrer, I.; Aronica, E.; Kovacs, G.G.; Verney, C.; Nardelli, J.; Khung, S.; Delezoide, A.L.; Milenkovic, I.; Rasika, S.; et al. Hippocampal Radial Glial Subtypes and Their Neurogenic Potential in Human Fetuses and Healthy and Alzheimer’s Disease Adults. Cereb. Cortex 2018, 28, 2458–2478. [Google Scholar] [CrossRef] [PubMed]
- Mamber, C.; Kamphuis, W.; Haring, N.L.; Peprah, N.; Middeldorp, J.; Hol, E.M. GFAPδ expression in glia of the developmental and adolescent mouse brain. PLoS ONE 2012, 7, e52659. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, L.N.; Chong, L.; Liu, Y.; Xi, F.Y.; Zhang, H.; Duan, X.L. Prenatal ethanol exposure impairs the formation of radial glial fibers and promotes the transformation of GFAPδ-positive radial glial cells into astrocytes. Mol. Med. Rep. 2021, 23, 1. [Google Scholar] [CrossRef] [PubMed]
- Martins-Macedo, J.; Lepore, A.C.; Domingues, H.S.; Salgado, A.J.; Gomes, E.D.; Pinto, L. Glial restricted precursor cells in central nervous system disorders: Current applications and future perspectives. Glia 2021, 69, 513–531. [Google Scholar] [CrossRef]
- Biron, K.E.; Dickstein, D.L.; Gopaul, R.; Jefferies, W.A. Amyloid triggers extensive cerebral angiogenesis causing blood brain barrier permeability and hypervascularity in Alzheimer’s disease. PLoS ONE 2011, 6, e23789. [Google Scholar] [CrossRef] [Green Version]
- Pozhilenkova, E.A.; Lopatina, O.L.; Komleva, Y.K.; Salmin, V.V.; Salmina, A.B. Blood-brain barrier-supported neurogenesis in healthy and diseased brain. Rev. Neurosci. 2017, 28, 397–415. [Google Scholar] [CrossRef]
- Simpson, J.E.; Ince, P.G.; Shaw, P.J.; Heath, P.R.; Raman, R.; Garwood, C.J.; Gelsthorpe, C.; Baxter, L.; Forster, G.; Matthews, F.E.; et al. Microarray analysis of the astrocyte transcriptome in the aging brain: Relationship to Alzheimer’s pathology and APOE genotype. Neurobiol. Aging 2011, 32, 1795–1807. [Google Scholar] [CrossRef]
- Kamphuis, W.; Kooijman, L.; Orre, M.; Stassen, O.; Pekny, M.; Hol, E.M. GFAP and vimentin deficiency alters gene expression in astrocytes and microglia in wild-type mice and changes the transcriptional response of reactive glia in mouse model for Alzheimer’s disease. Glia 2015, 63, 1036–1056. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Rodrigues, J.J.; Pivoriunas, A.; Zorec, R.; Semyanov, A. Astroglial atrophy in Alzheimer’s disease. Pflüg. Arch. Eur. J. Physiol. 2019, 471, 1247–1261. [Google Scholar] [CrossRef]
- Nielsen, A.L.; Holm, I.E.; Johansen, M.; Bonven, B.; Jørgensen, P.; Jørgensen, A.L. A New Splice Variant of Glial Fibrillary Acidic Protein, GFAPε, Interacts with the Presenilin Proteins. J. Biol. Chem. 2002, 277, 29983–29991. [Google Scholar] [CrossRef] [Green Version]
- Kamphuis, W.; Middeldorp, J.; Kooijman, L.; Sluijs, J.A.; Kooi, E.-J.; Moeton, M.; Freriks, M.; Mizee, M.R.; Hol, E.M. Glial fibrillary acidic protein isoform expression in plaque related astrogliosis in Alzheimer’s disease. Neurobiol. Aging 2014, 35, 492–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frost, G.R.; Li, Y.-M. The role of astrocytes in amyloid production and Alzheimer’s disease. Open Biol. 2017, 7, 170228. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Esposito, G.; Scuderi, C.; Steardo, L.; Delli-Bovi, L.C.; Hecht, J.L.; Dickinson, B.C.; Chang, C.J.; Mori, T.; Sheen, V. S100B and APP Promote a Gliocentric Shift and Impaired Neurogenesis in Down Syndrome Neural Progenitors. PLoS ONE 2011, 6, e22126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeidán-Chuliá, F.; de Oliveira, B.H.N.; Salmina, A.B.; Casanova, M.F.; Gelain, D.P.; Noda, M.; Verkhratsky, A.; Moreira, J.C.F. Altered expression of Alzheimer’s disease-related genes in the cerebellum of autistic patients: A model for disrupted brain connectome and therapy. Cell Death Dis. 2014, 5, e1250. [Google Scholar] [CrossRef]
- Yan, S.D.; Bierhaus, A.; Nawroth, P.P.; Stern, D.M. RAGE and Alzheimer’s disease: A progression factor for amyloid-beta-induced cellular perturbation? J. Alzheimers Dis. 2009, 16, 833–843. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.-Y.; Tan, M.-S.; Yu, J.-T.; Tan, L. Role of pro-inflammatory cytokines released from microglia in Alzheimer’s disease. Ann. Transl. Med. 2015, 3, 136. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, T.; Sun, G.Y.; Ding, S. Specific disruption of astrocytic Ca2+ signaling pathway in vivo by adeno-associated viral transduction. Neuroscience 2010, 170, 992–1003. [Google Scholar] [CrossRef] [Green Version]
- Shigetomi, E.; Bushong, E.A.; Haustein, M.D.; Tong, X.; Jackson-Weaver, O.; Kracun, S.; Xu, J.; Sofroniew, M.V.; Ellisman, M.H.; Khakh, B.S. Imaging calcium microdomains within entire astrocyte territories and endfeet with GCaMPs expressed using adeno-associated viruses. J. Gen. Physiol. 2013, 141, 633–647. [Google Scholar] [CrossRef] [PubMed]
- Von Jonquieres, G.; Mersmann, N.; Klugmann, C.B.; Harasta, A.E.; Lutz, B.; Teahan, O.; Housley, G.D.; Fröhlich, D.; Krämer-Albers, E.M.; Klugmann, M. Glial promoter selectivity following AAV-delivery to the immature brain. PLoS ONE 2013, 8, e65646. [Google Scholar] [CrossRef]
- Meng, X.; Yang, F.; Ouyang, T.; Liu, B.; Wu, C.; Jiang, W. Specific gene expression in mouse cortical astrocytes is mediated by a 1740bp-GFAP promoter-driven combined adeno-associated virus 2/5/7/8/9. Neurosci. Lett. 2015, 593, 45–50. [Google Scholar] [CrossRef]
- Shinohara, Y.; Konno, A.; Takahashi, N.; Matsuzaki, Y.; Kishi, S.; Hirai, H. Viral vector-based dissection of marmoset GFAP promoter in mouse and marmoset brains. PLoS ONE 2016, 11, e0162023. [Google Scholar] [CrossRef] [Green Version]
- Testen, A.; Kim, R.; Reissner, K.J. High-resolution three-dimensional imaging of individual astrocytes using confocal microscopy. Curr. Protoc. Neurosci. 2020, 91, e92. [Google Scholar] [CrossRef]
- De Leeuw, B.; Su, M.; ter Horst, M.; Iwata, S.; Rodijk, M.; Hoeben, R.C.; Messing, A.; Smitt, P.S.; Brenner, M.J. Increased glia-specific transgene expression with glial fibrillary acidic protein promoters containing multiple enhancer elements. Neurosci. Res. 2006, 83, 744–753. [Google Scholar] [CrossRef]
- Lee, Y.; Su, M.; Messing, A.; Brenner, M. Astrocyte heterogeneity revealed by expression of a GFAP-LacZ transgene. Glia 2006, 53, 677–687. [Google Scholar] [CrossRef]
- Theofilas, P.; Brar, S.; Stewart, K.A.; Shen, H.Y.; Sandau, U.S.; Poulsen, D.; Boison, D. Adenosine kinase as a target for therapeutic antisense strategies in epilepsy. Epilepsia 2011, 52, 589–601. [Google Scholar] [CrossRef] [Green Version]
- Dirren, E.; Towne, C.L.; Setola, V.; Redmond, D.E., Jr.; Schneider, B.L.; Aebischer, P. Intracerebroventricular injection of adeno-associated virus 6 and 9 vectors for cell type-specific transgene expression in the spinal cord. Hum. Gene Ther. 2014, 25, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Vagner, T.; Dvorzhak, A.; Wójtowicz, A.M.; Harms, C.; Grantyn, R. Systemic application of AAV vectors targeting GFAP-expressing astrocytes in Z-Q175-KI Huntington’s disease mice. Mol. Cell. Neurosci. 2016, 77, 76–86. [Google Scholar] [CrossRef]
- Griffin, J.M.; Fackelmeier, B.; Fong, D.M.; Mouravlev, A.; Young, D.; O’Carroll, S.J. Astrocyte-selective AAV gene therapy through the endogenous GFAP promoter results in robust transduction in the rat spinal cord following injury. Gene Ther. 2019, 26, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Hammond, S.L.; Leek, A.N.; Richman, E.H.; Tjalkens, R.B. Cellular selectivity of AAV serotypes for gene delivery in neurons and astrocytes by neonatal intracerebroventricular injection. PLoS ONE 2017, 12, e0188830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jollé, C.; Déglon, N.; Pythoud, C.; Bouzier-Sore, A.K.; Pellerin, L. Development of Efficient AAV2/DJ-Based Viral Vectors to Selectively Downregulate the Expression of Neuronal or Astrocytic Target Proteins in the Rat Central Nervous System. Front. Mol. Neurosci. 2019, 12, 201. [Google Scholar] [CrossRef] [Green Version]
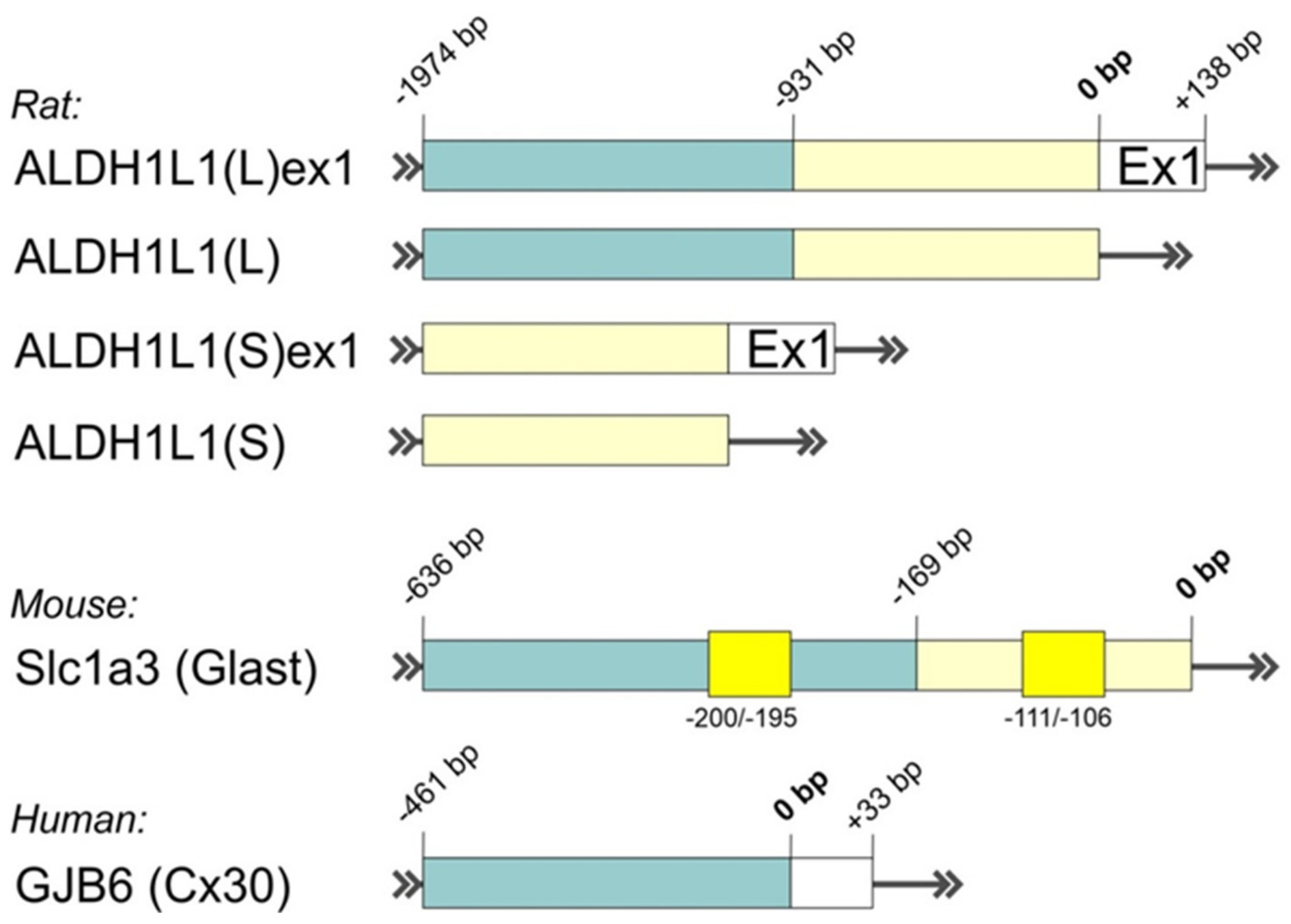
- Mudannayake, J.M.; Mouravlev, A.; Fong, D.M.; Young, D. Transcriptional activity of novel ALDH1L1 promoters in the rat brain following AAV vector-mediated gene transfer. Mol. Ther. Methods Clin. Dev. 2016, 3, 16075. [Google Scholar] [CrossRef]
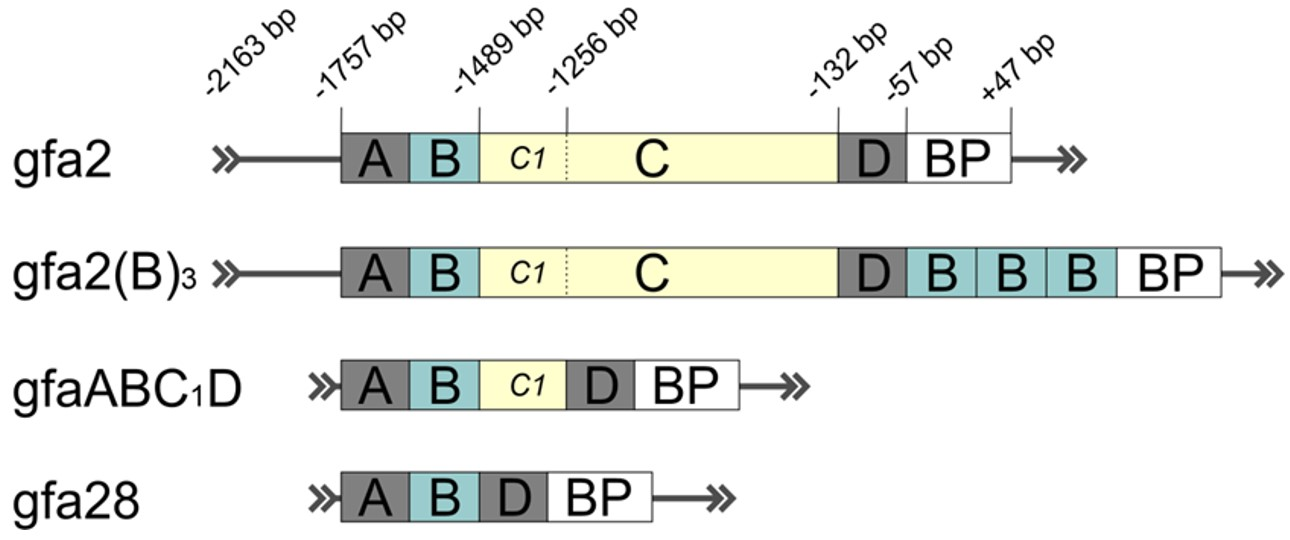
- Taschenberger, G.; Tereshchenko, J.; Kügler, S. A MicroRNA124 Target Sequence Restores Astrocyte Specificity of gfaABC1D-Driven Transgene Expression in AAV-Mediated Gene Transfer. Mol. Ther. Nucleic Acids 2017, 8, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Preston, A.N.; Cervasio, D.A.; Laughlin, S.T. Visualizing the brain’s astrocytes. Methods Enzymol. 2019, 622, 129–151. [Google Scholar] [CrossRef] [PubMed]
- Hu, N.Y.; Chen, Y.T.; Wang, Q.; Jie, W.; Liu, Y.S.; You, Q.L.; Li, Z.L.; Li, X.W.; Reibel, S.; Pfrieger, F.W.; et al. Expression Patterns of Inducible Cre Recombinase Driven by Differential Astrocyte-Specific Promoters in Transgenic Mouse Lines. Neurosci. Bull. 2020, 36, 530–544. [Google Scholar] [CrossRef]
- Srinivasan, R.; Lu, T.Y.; Chai, H.; Xu, J.; Huang, B.S.; Golshani, P.; Coppola, G.; Khakh, B.S. New Transgenic Mouse Lines for Selectively Targeting Astrocytes and Studying Calcium Signals in Astrocyte Processes in Situ and In Vivo. Neuron 2016, 92, 1181–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, W.; Park, Y.M.; Lee, S.E.; Lee, C.J. AAV-Mediated Astrocyte-Specific Gene Expression under Human ALDH1L1 Promoter in Mouse Thalamus. Exp. Neurobiol. 2017, 26, 350–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagiwara, T.; Tanaka, K.; Takai, S.; Maeno-Hikichi, Y.; Mukainaka, Y.; Wada, K. Genomic organization, promoter analysis, and chromosomal localization of the gene for the mouse glial high-affinity glutamate transporter Slc1a3. Genomics 1996, 33, 508–515. [Google Scholar] [CrossRef]
- Essenfelder, G.M.; Larderet, G.; Waksman, G.; Lamartine, J. Gene structure and promoter analysis of the human GJB6 gene encoding connexin 30. Gene 2005, 350, 33–40. [Google Scholar] [CrossRef]
- Deverman, B.E.; Pravdo, P.L.; Simpson, B.P.; Kumar, S.R.; Chan, K.Y.; Banerjee, A.; Wu, W.L.; Yang, B.; Huber, N.; Pasca, S.P.; et al. Cre-dependent selection yields AAV variants for widespread gene transfer to the adult brain. Nat. Biotechnol. 2016, 34, 204–209. [Google Scholar] [CrossRef]
- Tervo, D.G.; Hwang, B.Y.; Viswanathan, S.; Gaj, T.; Lavzin, M.; Ritola, K.D.; Lindo, S.; Michael, S.; Kuleshova, E.; Ojala, D.; et al. A Designer AAV Variant Permits Efficient Retrograde Access to Projection Neurons. Neuron 2016, 92, 372–382. [Google Scholar] [CrossRef] [Green Version]
- O’Carroll, S.J.; Cook, W.H.; Young, D. AAV Targeting of Glial Cell Types in the Central and Peripheral Nervous System and Relevance to Human Gene Therapy. Front. Mol. Neurosci. 2021, 13, 618020. [Google Scholar] [CrossRef]
- Aldewachi, H.; Al-Zidan, R.N.; Conner, M.T.; Salman, M.M. High-Throughput Screening Platforms in the Discovery of Novel Drugs for Neurodegenerative Diseases. Bioengineering 2021, 8, 30. [Google Scholar] [CrossRef]
- Salman, M.M.; Al-Obaidi, Z.; Kitchen, P.; Loreto, A.; Bill, R.M.; Wade-Martins, R. Advances in Applying Computer-Aided Drug Design for Neurodegenerative Diseases. Int. J. Mol. Sci. 2021, 22, 4688. [Google Scholar] [CrossRef]
- Blankvoort, S.; Witter, M.P.; Noonan, J.; Cotney, J.; Kentros, C. Marked Diversity of Unique Cortical Enhancers Enables Neuron-Specific Tools by Enhancer-Driven Gene Expression. Curr. Biol. 2018, 28, 2103–2114. [Google Scholar] [CrossRef] [Green Version]
- Nair, R.R.; Blankvoort, S.; Lagartos, M.J.; Kentros, C. Enhancer-Driven Gene Expression (EDGE) Enables the Generation of Viral Vectors Specific to Neuronal Subtypes. iScience 2020, 23, 100888. [Google Scholar] [CrossRef] [Green Version]
- Mich, J.K.; Graybuck, L.T.; Hess, E.E.; Mahoney, J.T.; Kojima, Y.; Ding, Y.; Somasundaram, S.; Miller, J.A.; Kalmbach, B.E.; Radaelli, C.; et al. Functional enhancer elements drive subclass-selective expression from mouse to primate neocortex. Cell Rep. 2021, 34, 108754. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Name of Element | Length, kb | Source | Addgene # | |
|---|---|---|---|---|
| Promoter | Ubiquitous promoters | |||
| CBA (CAG) | 1.6 | Synthetic | ||
| CBA mini | 0.8 | Synthetic | ||
| CBh | 0.8 | Synthetic | ||
| CMV | 0.8 | Viral | ||
| Validated astrocytic promoters | ||||
| gfa2(B)3 | 2.6 | Human | #53132 | |
| gfa2 (hGFAP) | 2.2 | Human | #53126 | |
| gfaABC1D (gfap104) | 0.7 | Human | #53131, #122630 | |
| gfa28 | 0.45 | Human | #53130 | |
| Putative astrocytic promoters | ||||
| AldH1L1 | 0.9–2.1 | Human, Rat | ||
| Slc1a3 (Glast) | 0.64 | Mouse | ||
| GJB6 (Cx30) | 0.5 | Human | ||
| GOI (Opsin) | Melanopsin | 1.4 | Human | #122630 |
| Opto-a1AR | 1.3 | Chimeric | #20947 | |
| ChR2(H134R) | 0.94 | Algae * | #112496 | |
| Fluorescent protein | tdTomato | 1.4 | Anthozoa * | |
| EGFP | 0.72 | Hydrozoa * | ||
| EYFP | 0.72 | Hydrozoa * | ||
| mCherry | 0.71 | Anthozoa * | ||
| PRE | WPRE | 0.6 | Viral | |
| WPRE3 | 0.25 | Viral * | #61463 | |
| polyA | bGH | 0.22 | Bovine | |
| SV40 late | 0.13 | Viral | ||
| hGH | 0.48 | Human | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borodinova, A.A.; Balaban, P.M.; Bezprozvanny, I.B.; Salmina, A.B.; Vlasova, O.L. Genetic Constructs for the Control of Astrocytes’ Activity. Cells 2021, 10, 1600. https://doi.org/10.3390/cells10071600
Borodinova AA, Balaban PM, Bezprozvanny IB, Salmina AB, Vlasova OL. Genetic Constructs for the Control of Astrocytes’ Activity. Cells. 2021; 10(7):1600. https://doi.org/10.3390/cells10071600
Chicago/Turabian StyleBorodinova, Anastasia A., Pavel M. Balaban, Ilya B. Bezprozvanny, Alla B. Salmina, and Olga L. Vlasova. 2021. "Genetic Constructs for the Control of Astrocytes’ Activity" Cells 10, no. 7: 1600. https://doi.org/10.3390/cells10071600