
Cholate Disrupts Regulatory Functions of Cytochrome c Oxidase

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Isolation of the Enzyme
2.2. UV/VIS Absorption Spectroscopy
2.3. Equilibrium Dialysis
2.4. Kinetic Measurements
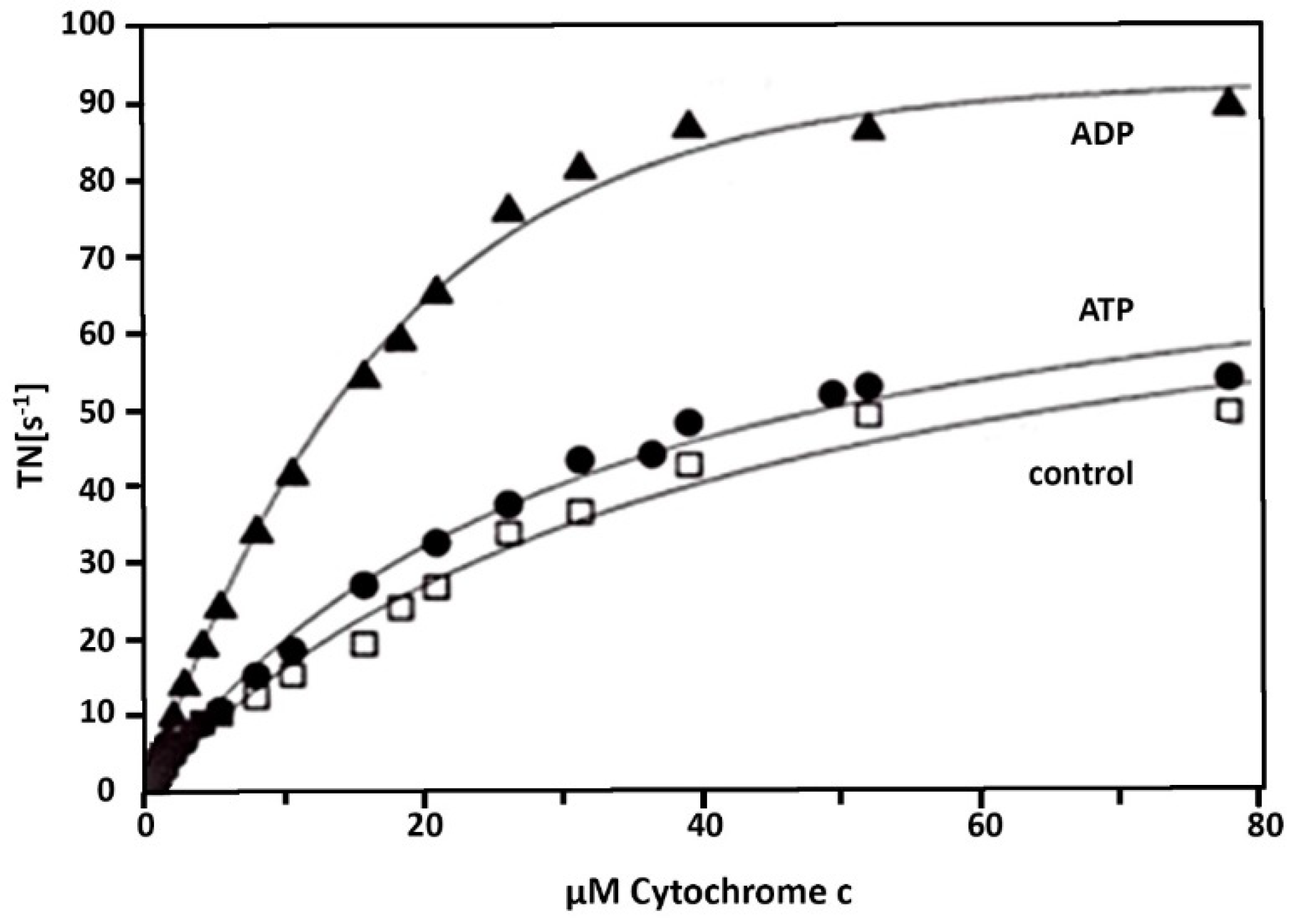
3. Results
4. Discussion
4.1. Cholate Mimicks the Nucleotide Binding on CytOx
4.2. Cholate Induces Dimerization and Reduces Complex IV Activity
4.3. Different Binding Properties of Nucleotids and Cholate Results in Different Enzymatic Activities of CytOx
4.4. Functions of CytOx Subunits and Supercomplexes Are not Fully Understood
4.5. Cholate May Hamper the Formation of Mitochondrial Supercomplexes
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Villani, G.; Attardi, G. In vivo control of respiration by cytochrome c oxidase in wildtype and mitochondrial DNA mutation carrying human cells. Proc. Natl. Acad. Sci. USA 1997, 94, 1166–1171. [Google Scholar] [CrossRef] [Green Version]
- Villani, G.; Attardi, G. In vivo control of respiration by cytochrome c oxidase in human cells. Free Radic. Biol. Med. 2000, 29, 202–210. [Google Scholar] [CrossRef]
- Ramzan, R.; Rhiel, A.; Weber, P.; Kadenbach, B.; Vogt, S. Reversible dimerization of cytochrome c oxidase regulates mitochondrial respiration. Mitochondrion 2019, 49, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Arnold, S.; Kadenbach, B. Priority Paper. Cell respiration is controlled by ATP, an allosteric inhibitor of cytochrome c oxidase. Eur. J. Biochem. 1997, 249, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Ramzan, R.; Vogt, S.; Kadenbach, B. Stress-mediated generation of deleterious ROS in healthy individuals—Role of cytochrome c oxidase. J. Mol. Med. 2020, 98, 651–657. [Google Scholar] [CrossRef] [Green Version]
- Kadenbach, B.; Arnold, S. Minireview. A second mechanism of respiratory control. FEBS Lett. 1999, 447, 131–134. [Google Scholar] [CrossRef] [Green Version]
- Vogt, S.; Rhiel, A.; Weber, P.; Ramzan, R. Revisiting Kadenbach: Electron flux rate through cytochrome c-oxidase determines the ATP-inhibitory effect and subsequent production of ROS. Bioessays 2016, 38, 556–567. [Google Scholar] [CrossRef] [Green Version]
- Ramzan, R.; Kadenbach, B.; Vogt, S. Multiple Mechanisms Regulate Eukaryotic Cytochrome C Oxidase. Cells 2021, 10, 514. [Google Scholar] [CrossRef]
- Ramzan, R.; Schaper, A.K.; Weber, P.; Rhiel, A.; Siddiq, M.S.; Vogt, S. Mitochondrial cytochrome c oxidase is inhibited by ATP only at very high ATP/ADP ratios. Biol. Chem. 2017, 398, 737–750. [Google Scholar] [CrossRef]
- Kadenbach, B.; Stroh, A.; Ungibauer, M.; Kuhn-Nentwig, L.; Büge, U.; Jarausch, J. Isoenzymes of cytochrome c oxidase: Characterization and isolation from different tissues. Methods Enzymol. 1986, 126, 32–45. [Google Scholar]
- Yoshikawa, S.; Tera, T.; Takahashi, Y.; Tsukihara, T. Crystalline cytochrome c oxidase of bovine heart mitochondrial membrane: Composition and x-ray diffraction studies. Proc. Natl. Acad. Sci. USA 1988, 85, 1354–1358. [Google Scholar] [CrossRef] [Green Version]
- Musatov, A.; Robinson, N.C. Cholate-Induced Dimerization of Detergent- or Phospholipid-Solubilized Bovine Cytochrome c Oxidase. Biochemistry 2002, 41, 4371–4376. [Google Scholar] [CrossRef]
- Napiwotzki, J.; Kadenbach, B. Extramitochondrial ATP/ADP-ratios regulate cytochrome c oxidase activity via binding to the cytosolic domain of subunit IV. Biol. Chem. 1998, 379, 335–339. [Google Scholar] [CrossRef]
- Kadenbach, B. Regulation of mammalian 13-subunit cytochrome c oxidase and binding of other proteins. Role of NDUFA4. Trends Endocrinol. Metab. 2017, 28, 761–770. [Google Scholar] [CrossRef]
- Kang, Y.; Fielden, L.F.; Stojanovski, D. Mitochondrial protein transport in health and disease. Semin. Cell Dev. Biol. 2018, 76, 142–153. [Google Scholar]
- Timón-Gómez, A.; Nývltová, E.; Abriata, L.A.; Vila, A.J.; Hosler, J.; Barrientos, A. Mitochondrial cytochrome c oxidase biogenesis: Recent developments. Semin. Cell Dev. Biol. 2018, 76, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Kadenbach, B.; Hüttemann, M. The subunit composition and function of mammalian cytochrome c oxidase. Mitochondrion 2015, 24, 64–76. [Google Scholar] [CrossRef]
- Sinkler, C.A.; Kalpage, H.; Shay, J.; Lee, I.; Malek, M.H.; Grossman, L.I.; Hüttemann, M. Tissue- and condition-specific isoforms of mammalian cytochrome c oxidase subunits: From function to human disease. Oxid. Med. Cell Longev. 2017, 2017, 1534056. [Google Scholar] [CrossRef] [Green Version]
- Acin-Perez, R.; Gatti, D.L.; Bai, Y.; Manfredi, G. Protein Phosphorylation and Prevention of Cytochrome Oxidase Inhibition by ATP: Coupled Mechanisms of Energy Metabolism Regulation. Cell Metab. 2011, 13, 712–719. [Google Scholar] [CrossRef] [Green Version]
- Horvat, S.; Beyer, C.; Arnold, S. Effect of hypoxia on the transcription pattern of subunit isoforms and the kinetics of cytochrome c oxidase in cortical astrocytes and cerebellar neurons. J. Neurochem. 2006, 99, 937–951. [Google Scholar] [CrossRef] [PubMed]
- Sommer, N.; Hüttemann, M.; Pak, O.; Scheibe, S.; Knoepp, F.; Sinkler, C.; Malczyk, M.; Gierhardt, M.; Esfandiary, A.; Kraut, S.; et al. Mitochondrial Complex IV Subunit 4 Isoform 2 Is Essential for Acute Pulmonary Oxygen Sensing. Circ. Res. 2017, 121, 424–438. [Google Scholar] [CrossRef]
- Arnold, S.; Goglia, F.; Kadenbach, B. 3,5-Diiodothyronine binds to subunit Va of cytochrome c oxidase and abolishes the allosteric inhibition of respiration by ATP. Eur. J. Biochem. 1998, 252, 325–330. [Google Scholar] [CrossRef]
- Kuznetsova, S.S.; Azarkina, N.V.; Vygodina, T.V.; Siletsky, S.A.; Konstantinov, A.A. Zinc Ions as Cytochrome c Oxidase Inhibitors: Two Sites of Action. Biochemistry 2005, 70, 128–136, Translated from Biokhimiya 2005, 70, 160–170. [Google Scholar] [CrossRef]
- Martino, P.-L.; Capitanio, G.; Capitanio, N.; Papa, S. Inhibition of proton pumping in membrane reconstituted bovine heart cytochrome c oxidase by zinc binding at the inner matrix side. Biochim. Biophys. Acta 2011, 1807, 1075–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anthony, G.; Reimann, A.; Kadenbach, B. Tissue-specific regulation of bovine heart cytochrome c oxidase by ADP via interaction with subunit VIa. Proc. Natl. Acad. Sci. USA 1993, 90, 1652–1656. [Google Scholar] [CrossRef] [Green Version]
- Frank, V.; Kadenbach, B. Regulation of the H+/e--stoichiometry of cytochrome c oxidase from bovine heart by intraliposomal ATP/ADP ratios. FEBS Lett. 1996, 382, 121–124. [Google Scholar] [CrossRef] [Green Version]
- Napiwotzki, J. Bindung von Adeninnukleotiden an die Cytochrom c Oxidase und deren Regulation der Enzymaktivität. Ph.D. Thesis, Fachbereich Chemie, Philipps-University, Marburg, Germany, 1997. Available online: http://archiv.ub.unimarburg.de/diss/z1997/0325/pdf/djn.pdf (accessed on 1 January 2021).
- Napiwotzki, J.; Shinzawa-Itoh, K.; Yoshikawa, S.; Kadenbach, B. ATP and ADP bind to cytochrome c oxidase and regulate its activity. Biol. Chem. 1997, 378, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Arnold, S.; Kadenbach, B. The intramitochondrial ATP/ADP-ratio controls cytochrome c oxidase activity allosterically. FEBS Lett. 1999, 443, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Tsukihara, T.; Aoyama, H.; Yamashita, E.; Tomizaki, T.; Yamaguchi, H.; Shinzawa-Itoh, K.; Nakashima, R.; Yaono, R.; Yoshikawa, S. The whole structure of the 13-subunit oxidized cytochrome c oxidase at 2.8 Å. Science 1996, 272, 1136–1144. [Google Scholar] [CrossRef]
- Sedlák, E.; Varha, R.; Musatov, A.; Robinson, N.C. The Kinetic Stability of Cytochrome c Oxidase: Effect of Bound Phospholipid and Dimerization. Biophys. J. 2014, 107, 2941–2949. [Google Scholar] [CrossRef] [Green Version]
- Musatov, A.; Ortega-Lopez, J.; Robinson, N.C. Detergent-Solubilized Bovine Cytochrome c Oxidase: Dimerization Depends on the Amphiphilic Environment. Biochemistry 2000, 39, 12996–13004. [Google Scholar] [CrossRef]
- Kadenbach, B.; Jarausch, J.; Hartmann, R.; Merle, P. Separation of mammalian cytochrome c oxidase into 13 poly-peptides by a sodium dodecyl sulfate-gel electrophoretic procedure. Anal. Biochem. 1983, 129, 517–521. [Google Scholar] [CrossRef]
- Riistama, S.; Verkhovsky, M.I.; Laakkonen, L.; Wikström, M.; Puustinen, A. Interaction between the formyl group of heme a and arginine 54 in cytochrome aa(3) from Paracoccus denitrificans. Biochim. Biophys. Acta 2000, 1456, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Berg, J.; Liu, J.; Svahn, E.; Ferguson-Miller, S.; Brzezinski, P. Structural changes at the surface of cytochrome c oxidase alter the proton-pumping stoichiometry. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148116. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, S.; Shimada, A. Reaction Mechanism of Cytochrome c Oxidase. Chem. Rev. 2015, 115, 1936–1989. [Google Scholar] [CrossRef]
- Shinzawa-Itoh, K.; Sugimura, T.; Misaki, T.; Tadehara, Y.; Yamamoto, S.; Hanada, M.; Yano, N.; Nakagawa, T.; Uene, S.; Yamada, T.; et al. Monomeric structure of an active form of bovine cytochrome c oxidase. Proc. Natl. Acad. Sci. USA 2019, 116, 19945–19951. [Google Scholar] [CrossRef] [Green Version]
- Kadenbach, B.; Ramzan, R.; Wen, L.; Vogt, S. New extension of the Mitchell Theory for oxidative phosphorylation in mitochondria of living organisms. Biochim. Biophys. Acta 2010, 1800, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Zong, S.; Wu, M.; Gu, J.; Liu, T.; Guo, R.; Yang, M. Structure of the intact 14-subunit human cytochrome c oxidase. Cell Res. 2018, 28, 1026–1034. [Google Scholar] [CrossRef] [Green Version]
- Liko, I.; Degiacomi, M.T.; Mohammed, S.; Yoshikawa, S.; Schmidt, C.; Robinson, C.V. Dimer interface of bovine cytochrome c oxidase is influenced by local posttranslational modifications and lipid binding. Proc. Natl. Acad. Sci. USA 2016, 113, 8230–8235. [Google Scholar] [CrossRef] [Green Version]
- Vidoni, S.; Harbour, M.E.; Guerrero-Castillo, S.; Signes, A.; Ding, S.; Fearnley, I.M.; Taylor, R.W.; Tiranti, V.; Arnold, S.; Fernandez-Vizarra, E.; et al. MR-1S Interacts with PET100 and PET117 in Module-Based Assembly of Human Cytochrome c Oxidase. Cell Rep. 2017, 18, 1727–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadenbach, B. Regulation of cytochrome c oxidase contributes to health and optimal life. World J. Biol. Chem. 2020, 11, 52–61. [Google Scholar] [CrossRef]
- Kadenbach, B. Complex IV—The regulatory center of mitochondrial oxidative phosphorylation. Mitochondrion 2021, 58, 296–302. [Google Scholar] [CrossRef]
- Hartley, A.M.; Lukoyanova, N.; Zhang, Y.; Cabrera-Orefice, A.; Arnold, S.; Meunier, B.; Pinotsis, N.; Maréchal, A. Structure of yeast cytochrome c oxidase in a supercomplex with cytochrome bc1. Nat. Struct. Mol. Biol. 2019, 26, 78–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helling, S.; Hüttemann, M.; Ramzan, R.; Kim, S.H.; Lee, I.; Müller, T.; Langenfeld, E.; Meyer, H.E.; Kadenbach, B.; Vogt, S.; et al. Multiple phosphorylations of cytochrome c oxidase and their functions. Proteomics 2012, 12, 950–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helling, S.; Hüttemann, M.; Kadenbach, B.; Ramzan, R.; Vogt, S.; Marcus, K. Discovering the phosphoproteome of the hydrophobic cytochrome c oxidase membrane protein complex. Methods Mol. Biol. 2012, 893, 345–358. [Google Scholar] [PubMed]
- Hüttemann, M.; Helling, S.; Sanderson, T.H.; Sinkler, C.; Samavati, L.; Mahapatra, G.; Varughese, A.; Lu, G.; Liu, J.; Ramzan, R.; et al. Regulation of mitochondrial respiration and apoptosis through cell signaling: Cytochrome c oxidase and cytochrome c in ischemia/reperfusion injury and inflammation. Biochim. Biophys. Acta 2012, 1817, 598–609. [Google Scholar] [CrossRef]
- Bultema, J.B.; Braun, H.P.; Boekema, E.J.; Kouril, R. Megacomplex organization of the oxidative phosphorylation system by structural analysis of respiratory supercomplexes from potato. Biochim. Biophys. Acta 2009, 1787, 60–67. [Google Scholar] [CrossRef] [Green Version]
- Shoji, K.; Giuffro, A.; D’Itri, E.; Hagiwara, K.; Yamanaka, T.; Brunori, M.; Sarti, P. The ratio between the fast and slow forms of bovine cytochrome c oxidase is changed by cholate or nucleotides bound to the cholate-binding site close to the cytochrome a3/CuB binuclear centre. Cell Mol. Life Sci. 2000, 57, 1482–1487. [Google Scholar] [CrossRef]
- Rolo, A.P.; Oliveira, P.J.; Moreno, A.J.; Palmeira, C.M. Bile acids affect liver mitochondrial bioenergetics: Possible relevance for cholestasis therapy. Toxicol. Sci. 2000, 57, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Krähenbühl, S.; Talos, C.; Fischer, S.; Reichen, J. Toxicity of bile acids on the electron transport chain of isolated rat liver mitochondria. Hepatology 1994, 19, 471–479. [Google Scholar] [CrossRef]
- Palmeira, C.M.; Rolo, A.P. Mitochondrially-mediated toxicity of bile acids. Toxicology 2004, 203, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, M.; Coxito, P.M.; Sardão, V.A.; Palmeira, C.M.; Oliveira, P.J. Bile acids are toxic for isolated cardiac mitochondria: A possible cause for hepatic-derived cardiomyopathies? Cardiovasc. Toxicol. 2005, 5, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Stuchebrukhov, A.; Schäfer, J.; Berg, J.; Brzezinski, P. Kinetic advantage of forming respiratory supercomplexes. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148193. [Google Scholar] [CrossRef]
- Volwerk, J.J.; Mrsny, R.J.; Patapoff, T.W.; Jost, P.C.; Griffith, O.H. Multiple equilibria binding treatment of lipid and detergent interactions with membrane proteins. Application to cytochrome c oxidase solubilized in cholate. Biochemistry 1987, 26, 466–475. [Google Scholar] [CrossRef]
- Nesterov, S.; Chesnokov, Y.; Kamyshinsky, R.; Panteleeva, A.; Lyamzaev, K.; Vasilov, R.; Yaguzhinsky, L. Ordered Clusters of the Complete Oxidative Phosphorylation System in Cardiac Mitochondria. Int. J. Mol. Sci. 2021, 22, 1462. [Google Scholar] [CrossRef] [PubMed]
- Acín-Pérez, R.; Fernández-Silva, P.; Peleato, M.L.; Pérez-Martos, A.; Enriquez, J.A. Respiratory active mitochondrial supercomplexes. Mol. Cell. 2008, 32, 529–539. [Google Scholar] [CrossRef]
- Ishibe, N.; Lynch, S.R.; Copeland, R.A. The pH dependence of cytochrome a conformation in cytochrome c oxidase. J. Biol. Chem. 1991, 266, 23916–23920. [Google Scholar] [CrossRef]
- Virji, M.; Knowles, P.F. Protein-lipid interactions in cytochrome oxidase from Saccharomyces cerevisiae. Effects of detergents and reconstitution of enzyme activity by phospholipids by using cholate-mediated exchange. Biochem. J. 1978, 169, 343–353. [Google Scholar] [CrossRef] [Green Version]
- Shinzawa-Itoh, K.; Hatanaka, M.; Fujita, K.; Yano, N.; Ogasawara, Y.; Iwata, J.; Yamashita, E.; Tsukihara, T.; Yoshikawa, S.; Muramoto, K. The 1.3-Å resolution structure of bovine cytochrome c oxidase suggests a dimerization mechanism. BBA Adv. 2021, 1, 100009. [Google Scholar] [CrossRef]
- Ramírez-Aguilar, S.J.; Keuthe, M.; Rocha, M.; Fedyaev, V.V.; Kramp, K.; Gupta, K.J.; Rasmusson, A.G.; Schulze, W.X.; van Dongen, J.T. The composition of plant mitochondrial supercomplexes changes with oxygen availability. J. Biol. Chem. 2011, 286, 43045–43053. [Google Scholar] [CrossRef] [Green Version]
- Vasavan, T.; Ferraro, E.; Ibrahim, E.; Dixon, P.; Gorelik, J.; Williamson, C. Heart and bile acids—Clinical consequences of altered bile acid metabolism. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864 Pt B, 1345–1355. [Google Scholar] [CrossRef]
- Kanaan, G.N.; Patten, D.A.; Redpath, C.J.; Harper, M.E. Atrial Fibrillation Is Associated with Impaired Atrial Mitochondrial Energetics and Supercomplex Formation in Adults with Type 2 Diabetes. Can. J. Diabetes 2019, 43, 67–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramzan, R.; Napiwotzki, J.; Weber, P.; Kadenbach, B.; Vogt, S. Cholate Disrupts Regulatory Functions of Cytochrome c Oxidase. Cells 2021, 10, 1579. https://doi.org/10.3390/cells10071579
Ramzan R, Napiwotzki J, Weber P, Kadenbach B, Vogt S. Cholate Disrupts Regulatory Functions of Cytochrome c Oxidase. Cells. 2021; 10(7):1579. https://doi.org/10.3390/cells10071579
Chicago/Turabian StyleRamzan, Rabia, Jörg Napiwotzki, Petra Weber, Bernhard Kadenbach, and Sebastian Vogt. 2021. "Cholate Disrupts Regulatory Functions of Cytochrome c Oxidase" Cells 10, no. 7: 1579. https://doi.org/10.3390/cells10071579