
Sex-Specific Isolation and Propagation of Human Premeiotic Fetal Germ Cells and Germ Cell-Like Cells

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Permissions
2.2. Culture of hESCs
2.3. Differentiation of hESCs into hPGCLCs
2.4. Human Fetal Gonads
2.5. Fluorescence Activated Cell Sorting (FACS)
2.6. Extended Culture of hPGCLCs
2.7. Immunofluorescence, Imaging and Quantification
2.8. Analysis of RNA Sequencing (RNASeq) Data
3. Results
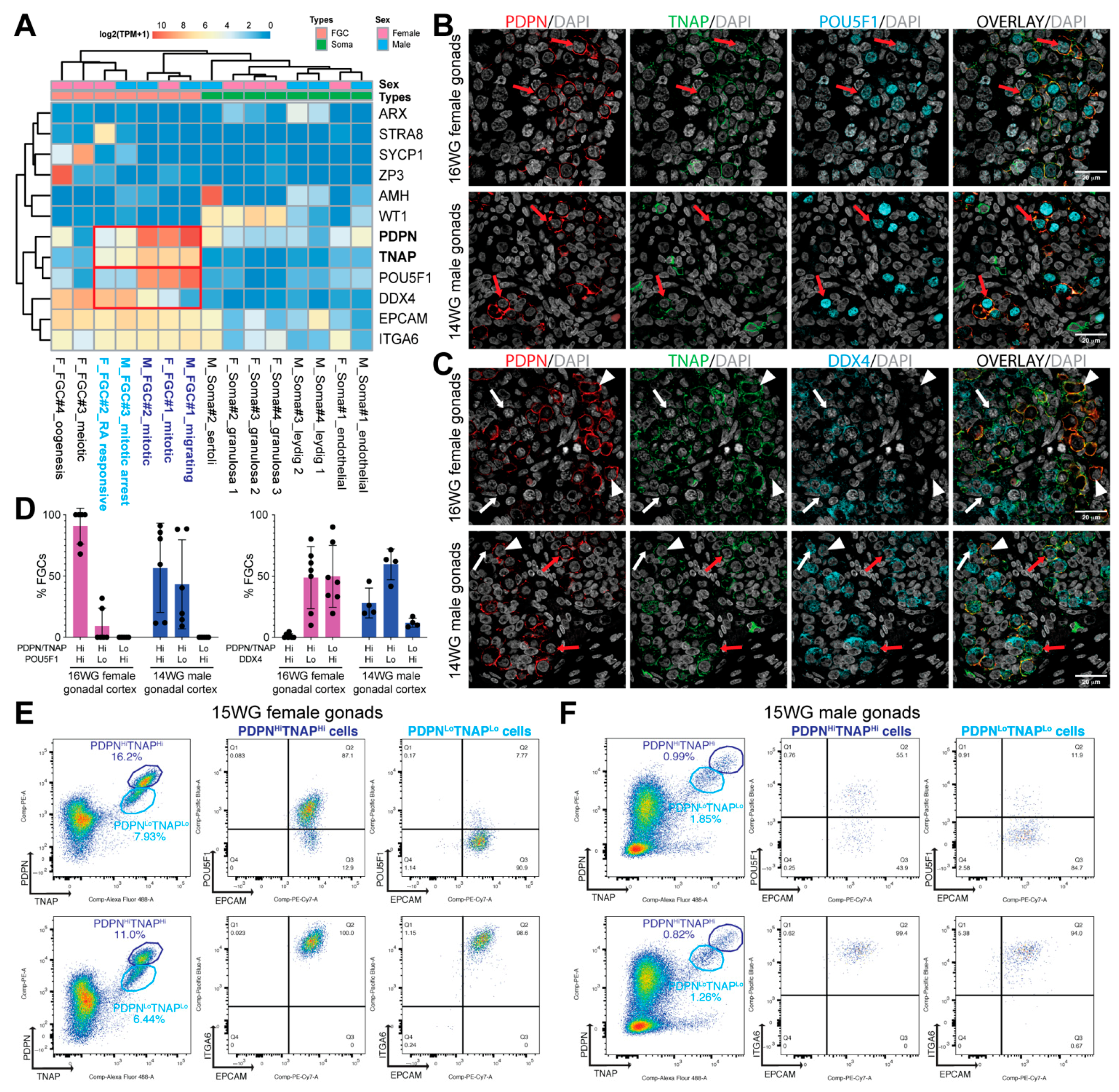
3.1. Surface Markers to Separate POU5F1HiDDX4Lo from POU5F1LoDDX4Hi Premeiotic hFGCs
3.2. Isolation of POU5F1Hi and POU5F1Lo Premeiotic hFGCs Using FACS
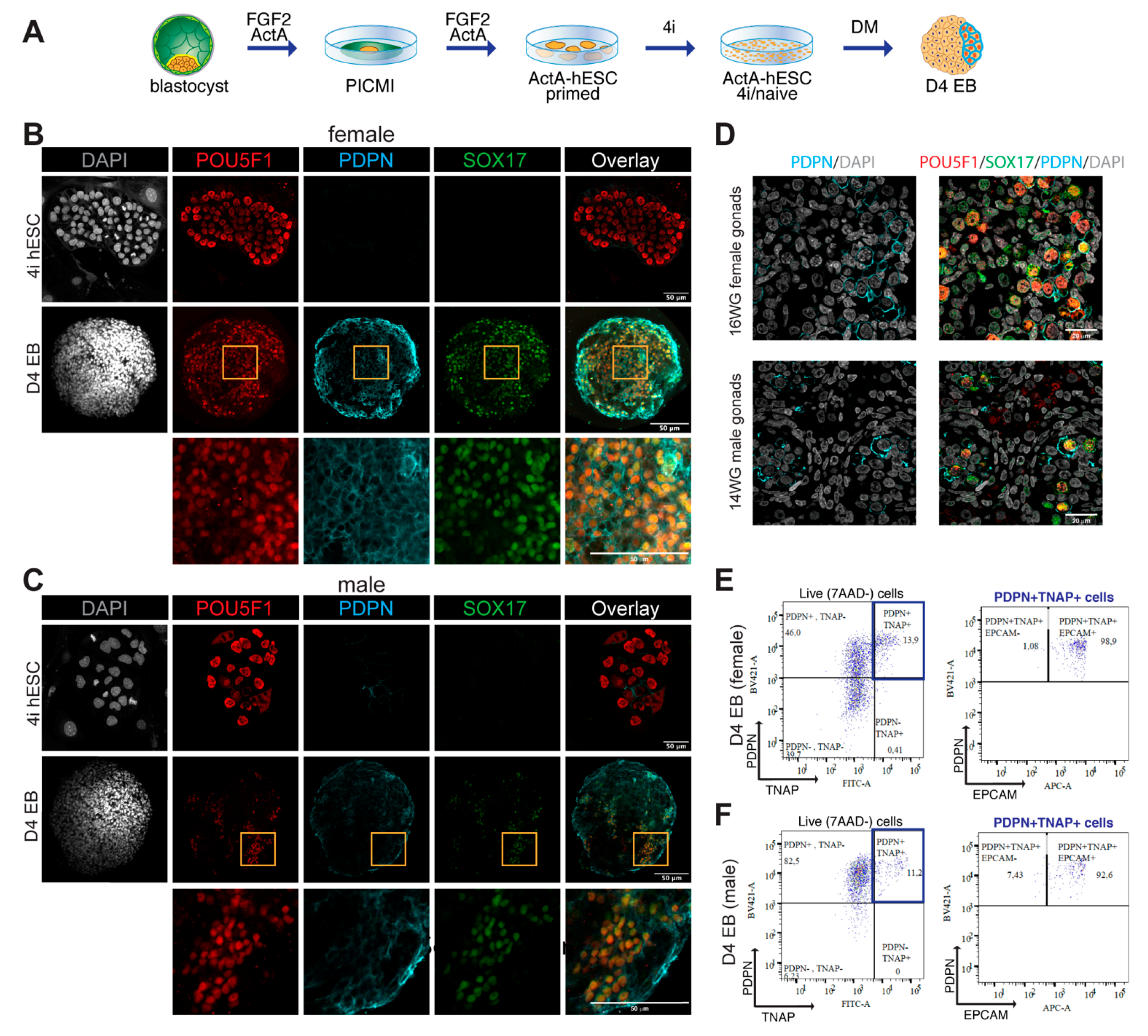
3.3. TNAP and PDPN Are Suitable to Isolate Male and Female hPGCLCs by FACS
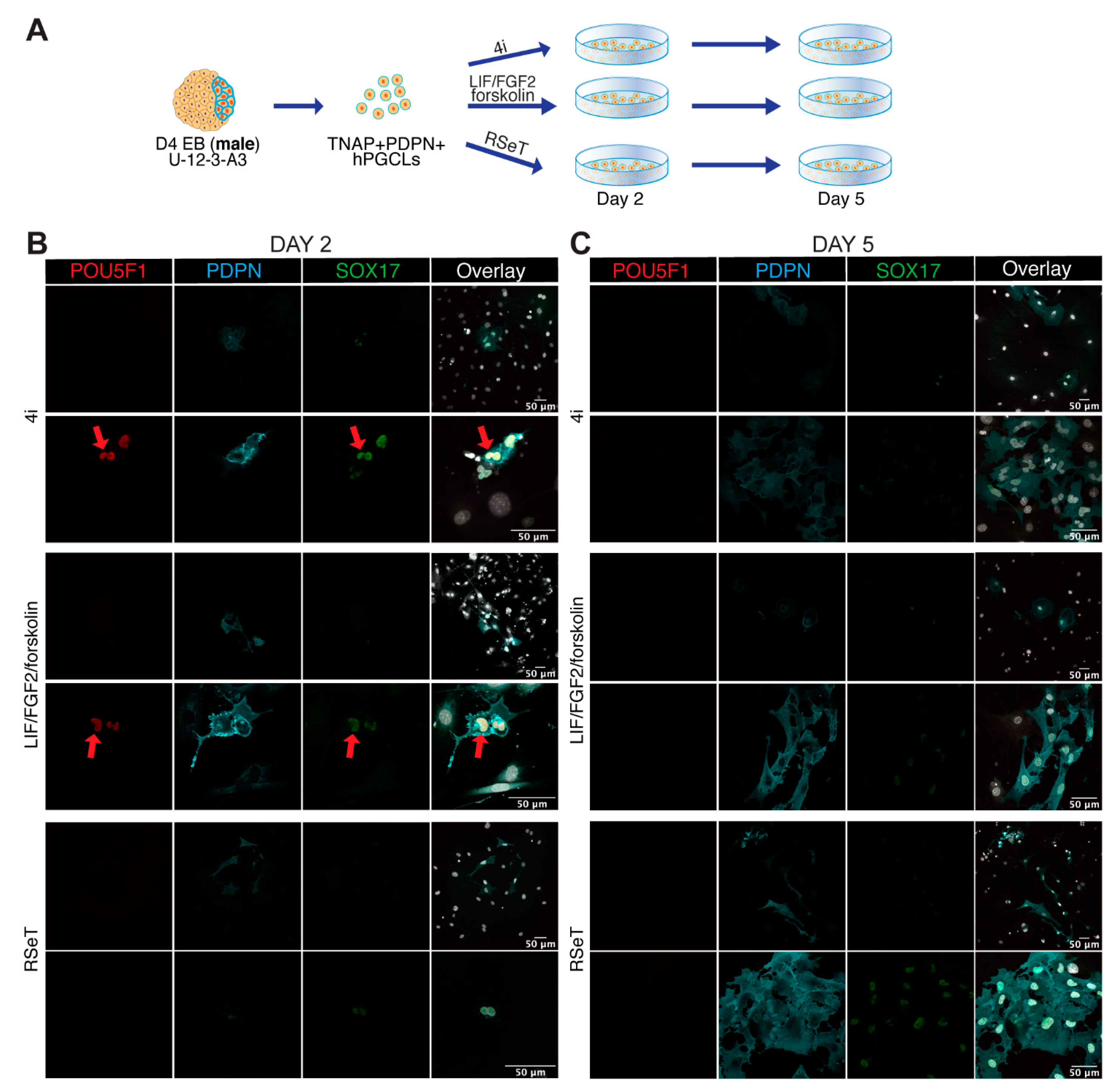
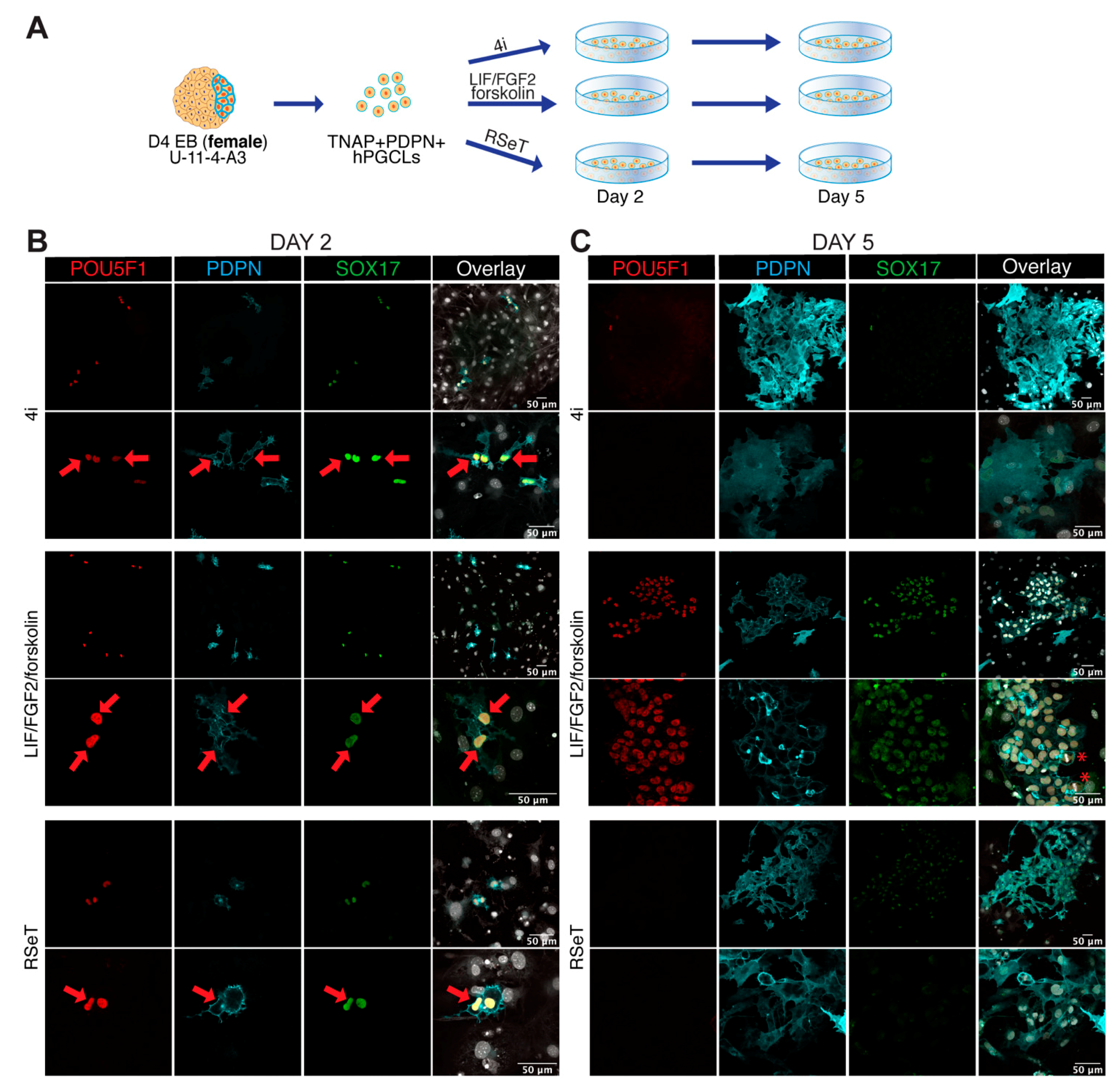
3.4. Male and Female hPGCLCs Propagate Differently in LFF Medium
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kobayashi, T.; Surani, M.A. On the origin of the human germline. Development 2018, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popovic, M.; Bialecka, M.; Gomes Fernandes, M.; Taelman, J.; Van Der Jeught, M.; De Sutter, P.; Heindryckx, B.; Chuva De Sousa Lopes, S.M. Human blastocyst outgrowths recapitulate primordial germ cell specification events. Mol. Hum. Reprod. 2019, 25, 519–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes Fernandes, M.; Bialecka, M.; Salvatori, D.C.F.; Chuva de Sousa Lopes, S.M. Characterization of migratory primordial germ cells in the aorta-gonad-mesonephros of a 4.5-week-old human embryo: A toolbox to evaluate in vitro early gametogenesis. Mol. Hum. Reprod. 2018, 24, 233–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, R.A.; Fulton, N.; Cowan, G.; Coutts, S.; Saunders, P.T. Conserved and divergent patterns of expression of DAZL, VASA and OCT4 in the germ cells of the human fetal ovary and testis. BMC Dev. Biol. 2007, 7, 136. [Google Scholar] [CrossRef] [Green Version]
- Heeren, A.M.; He, N.; de Souza, A.F.; Goercharn-Ramlal, A.; van Iperen, L.; Roost, M.S.; Gomes Fernandes, M.M.; van der Westerlaken, L.A.; Chuva de Sousa Lopes, S.M. On the development of extragonadal and gonadal human germ cells. Biol. Open 2016, 5, 185–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurilo, L.F. Oogenesis in antenatal development in man. Hum. Genet. 1981, 57, 86–92. [Google Scholar] [CrossRef]
- Li, L.; Dong, J.; Yan, L.; Yong, J.; Liu, X.; Hu, Y.; Fan, X.; Wu, X.; Guo, H.; Wang, X.; et al. Single-Cell RNA-Seq Analysis Maps Development of Human Germline Cells and Gonadal Niche Interactions. Cell Stem Cell 2017, 20, 858–873.e4. [Google Scholar] [CrossRef] [Green Version]
- Jorgensen, A.; Rajpert-De Meyts, E. Regulation of meiotic entry and gonadal sex differentiation in the human: Normal and disrupted signaling. Biomol. Concepts 2014, 5, 331–341. [Google Scholar] [CrossRef]
- Stoop, H.; Honecker, F.; Cools, M.; de Krijger, R.; Bokemeyer, C.; Looijenga, L.H. Differentiation and development of human female germ cells during prenatal gonadogenesis: An immunohistochemical study. Hum. Reprod. 2005, 20, 1466–1476. [Google Scholar] [CrossRef] [Green Version]
- Vertesy, A.; Arindrarto, W.; Roost, M.S.; Reinius, B.; Torrens-Juaneda, V.; Bialecka, M.; Moustakas, I.; Ariyurek, Y.; Kuijk, E.; Mei, H.; et al. Parental haplotype-specific single-cell transcriptomics reveal incomplete epigenetic reprogramming in human female germ cells. Nat. Commun. 2018, 9, 1873. [Google Scholar] [CrossRef]
- Chen, D.; Liu, W.; Lukianchikov, A.; Hancock, G.V.; Zimmerman, J.; Lowe, M.G.; Kim, R.; Galic, Z.; Irie, N.; Surani, M.A.; et al. Germline competency of human embryonic stem cells depends on eomesodermin. Biol. Reprod. 2017, 97, 850–861. [Google Scholar] [CrossRef] [Green Version]
- Gkountela, S.; Li, Z.; Vincent, J.J.; Zhang, K.X.; Chen, A.; Pellegrini, M.; Clark, A.T. The ontogeny of cKIT+ human primordial germ cells proves to be a resource for human germ line reprogramming, imprint erasure and in vitro differentiation. Nat. Cell Biol. 2013, 15, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.W.; Dietmann, S.; Irie, N.; Leitch, H.G.; Floros, V.I.; Bradshaw, C.R.; Hackett, J.A.; Chinnery, P.F.; Surani, M.A. A Unique Gene Regulatory Network Resets the Human Germline Epigenome for Development. Cell 2015, 161, 1453–1467. [Google Scholar] [CrossRef] [Green Version]
- Irie, N.; Weinberger, L.; Tang, W.W.; Kobayashi, T.; Viukov, S.; Manor, Y.S.; Dietmann, S.; Hanna, J.H.; Surani, M.A. SOX17 is a critical specifier of human primordial germ cell fate. Cell 2015, 160, 253–268. [Google Scholar] [CrossRef] [Green Version]
- Mitsunaga, S.; Odajima, J.; Yawata, S.; Shioda, K.; Owa, C.; Isselbacher, K.J.; Hanna, J.H.; Shioda, T. Relevance of iPSC-derived human PGC-like cells at the surface of embryoid bodies to prechemotaxis migrating PGCs. Proc. Natl. Acad Sci. USA 2017, 114, E9913–E9922. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, K.; Yokobayashi, S.; Nakamura, T.; Okamoto, I.; Yabuta, Y.; Kurimoto, K.; Ohta, H.; Moritoki, Y.; Iwatani, C.; Tsuchiya, H.; et al. Robust In Vitro Induction of Human Germ Cell Fate from Pluripotent Stem Cells. Cell Stem Cell 2015, 17, 178–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugawa, F.; Arauzo-Bravo, M.J.; Yoon, J.; Kim, K.P.; Aramaki, S.; Wu, G.; Stehling, M.; Psathaki, O.E.; Hubner, K.; Scholer, H.R. Human primordial germ cell commitment in vitro associates with a unique PRDM14 expression profile. EMBO J. 2015, 34, 1009–1024. [Google Scholar] [CrossRef] [PubMed]
- Sybirna, A.; Tang, W.W.C.; Pierson Smela, M.; Dietmann, S.; Gruhn, W.H.; Brosh, R.; Surani, M.A. A critical role of PRDM14 in human primordial germ cell fate revealed by inducible degrons. Nat. Commun. 2020, 11, 1282. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Taelman, J.; Popovic, M.; Tilleman, L.; Duthoo, E.; Van Der Jeught, M.; Deforce, D.; Van Nieuwerburgh, F.; Menten, B.; De Sutter, P.; et al. ActivinA-derived human embryonic stem cells show increased competence to differentiate into primordial germ cell-like cells. Stem Cells 2020, in press. [Google Scholar] [CrossRef]
- Murase, Y.; Yabuta, Y.; Ohta, H.; Yamashiro, C.; Nakamura, T.; Yamamoto, T.; Saitou, M. Long-term expansion with germline potential of human primordial germ cell-like cells in vitro. EMBO J. 2020, 39, e104929. [Google Scholar] [CrossRef]
- Gomes Fernandes, M.; He, N.; Wang, F.; Van Iperen, L.; Eguizabal, C.; Matorras, R.; Roelen, B.A.J.; Chuva De Sousa Lopes, S.M. Human-specific subcellular compartmentalization of P-element induced wimpy testis-like (PIWIL) granules during germ cell development and spermatogenesis. Hum. Reprod. 2018, 33, 258–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gell, J.J.; Liu, W.; Sosa, E.; Chialastri, A.; Hancock, G.; Tao, Y.; Wamaitha, S.E.; Bower, G.; Dey, S.S.; Clark, A.T. An Extended Culture System that Supports Human Primordial Germ Cell-like Cell Survival and Initiation of DNA Methylation Erasure. Stem Cell Rep. 2020, 14, 433–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shamblott, M.J.; Axelman, J.; Wang, S.; Bugg, E.M.; Littlefield, J.W.; Donovan, P.J.; Blumenthal, P.D.; Huggins, G.R.; Gearhart, J.D. Derivation of pluripotent stem cells from cultured human primordial germ cells. Proc. Natl. Acad Sci. USA 1998, 95, 13726–13731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnpenny, L.; Brickwood, S.; Spalluto, C.M.; Piper, K.; Cameron, I.T.; Wilson, D.I.; Hanley, N.A. Derivation of human embryonic germ cells: An alternative source of pluripotent stem cells. Stem Cells 2003, 21, 598–609. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bialecka, M.; Young, T.; Chuva de Sousa Lopes, S.; ten Berge, D.; Sanders, A.; Beck, F.; Deschamps, J. Cdx2 contributes to the expansion of the early primordial germ cell population in the mouse. Dev. Biol. 2012, 371, 227–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuva de Sousa Lopes, S.M.; Hayashi, K.; Shovlin, T.C.; Mifsud, W.; Surani, M.A.; McLaren, A. X chromosome activity in mouse XX primordial germ cells. PLoS Genet. 2008, 4, e30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padgett, D.M.; Cathro, H.P.; Wick, M.R.; Mills, S.E. Podoplanin is a better immunohistochemical marker for sarcomatoid mesothelioma than calretinin. Am. J. Surg. Pathol. 2008, 32, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Yang, R.; Yin, C.; Kee, K. Studying human reproductive biology through single-cell analysis and in vitro differentiation of stem cells into germ cell-like cells. Hum. Reprod. Update 2020, 26, 670–688. [Google Scholar] [CrossRef]
- Chen, D.; Gell, J.J.; Tao, Y.; Sosa, E.; Clark, A.T. Modeling human infertility with pluripotent stem cells. Stem Cell Res. 2017, 21, 187–192. [Google Scholar] [CrossRef]
- Mitsunaga, S.; Shioda, T. Evolutionarily diverse mechanisms of germline specification among mammals: What about us? Stem Cell Investig. 2018, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Zhang, H.; Tang, W.W.C.; Irie, N.; Withey, S.; Klisch, D.; Sybirna, A.; Dietmann, S.; Contreras, D.A.; Webb, R.; et al. Principles of early human development and germ cell program from conserved model systems. Nature 2017, 546, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Kojima, Y.; Sasaki, K.; Yokobayashi, S.; Sakai, Y.; Nakamura, T.; Yabuta, Y.; Nakaki, F.; Nagaoka, S.; Woltjen, K.; Hotta, A.; et al. Evolutionarily Distinctive Transcriptional and Signaling Programs Drive Human Germ Cell Lineage Specification from Pluripotent Stem Cells. Cell Stem Cell 2017, 21, 517–532.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, K.; Nakamura, T.; Okamoto, I.; Yabuta, Y.; Iwatani, C.; Tsuchiya, H.; Seita, Y.; Nakamura, S.; Shiraki, N.; Takakuwa, T.; et al. The Germ Cell Fate of Cynomolgus Monkeys Is Specified in the Nascent Amnion. Dev. Cell 2016, 39, 169–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Meyenn, F.; Berrens, R.V.; Andrews, S.; Santos, F.; Collier, A.J.; Krueger, F.; Osorno, R.; Dean, W.; Rugg-Gunn, P.J.; Reik, W. Comparative Principles of DNA Methylation Reprogramming during Human and Mouse In Vitro Primordial Germ Cell Specification. Dev. Cell 2016, 39, 104–115. [Google Scholar] [CrossRef] [Green Version]
- Greber, B.; Lehrach, H.; Adjaye, J. Fibroblast growth factor 2 modulates transforming growth factor beta signaling in mouse embryonic fibroblasts and human ESCs (hESCs) to support hESC self-renewal. Stem Cells 2007, 25, 455–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mishra, S.; Taelman, J.; Chang, Y.W.; Boel, A.; De Sutter, P.; Heindryckx, B.; Chuva De Sousa Lopes, S.M. Sex-Specific Isolation and Propagation of Human Premeiotic Fetal Germ Cells and Germ Cell-Like Cells. Cells 2021, 10, 1214. https://doi.org/10.3390/cells10051214
Mishra S, Taelman J, Chang YW, Boel A, De Sutter P, Heindryckx B, Chuva De Sousa Lopes SM. Sex-Specific Isolation and Propagation of Human Premeiotic Fetal Germ Cells and Germ Cell-Like Cells. Cells. 2021; 10(5):1214. https://doi.org/10.3390/cells10051214
Chicago/Turabian StyleMishra, Swati, Jasin Taelman, Yolanda W. Chang, Annekatrien Boel, Petra De Sutter, Björn Heindryckx, and Susana M. Chuva De Sousa Lopes. 2021. "Sex-Specific Isolation and Propagation of Human Premeiotic Fetal Germ Cells and Germ Cell-Like Cells" Cells 10, no. 5: 1214. https://doi.org/10.3390/cells10051214