
Chronic Alcohol Exposure of Cells Using Controlled Alcohol-Releasing Capillaries


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
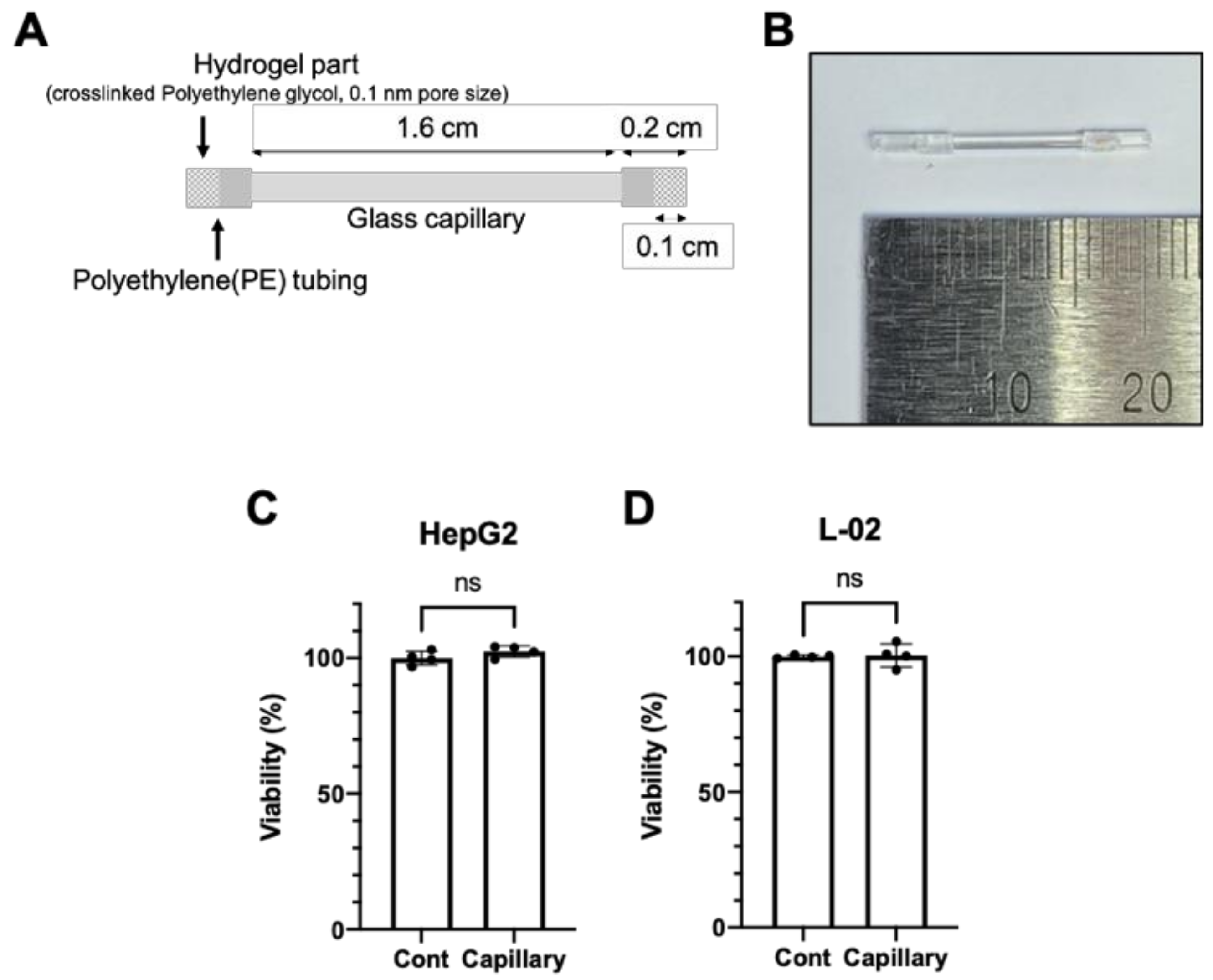
2.1. Fabrication of Glass Capillary Device
2.2. Cell Culture and Ethanol Treatment
2.3. RNA Quantification
2.4. Cell Viability and Ethanol Measurement
2.5. Oil Red O Staining
2.6. Statistical Analysis
3. Results
3.1. The Glass Capillary Device Led to a Controlled Release of Ethanol
3.2. Glass Capillary System for the Controlled Release of Ethanol
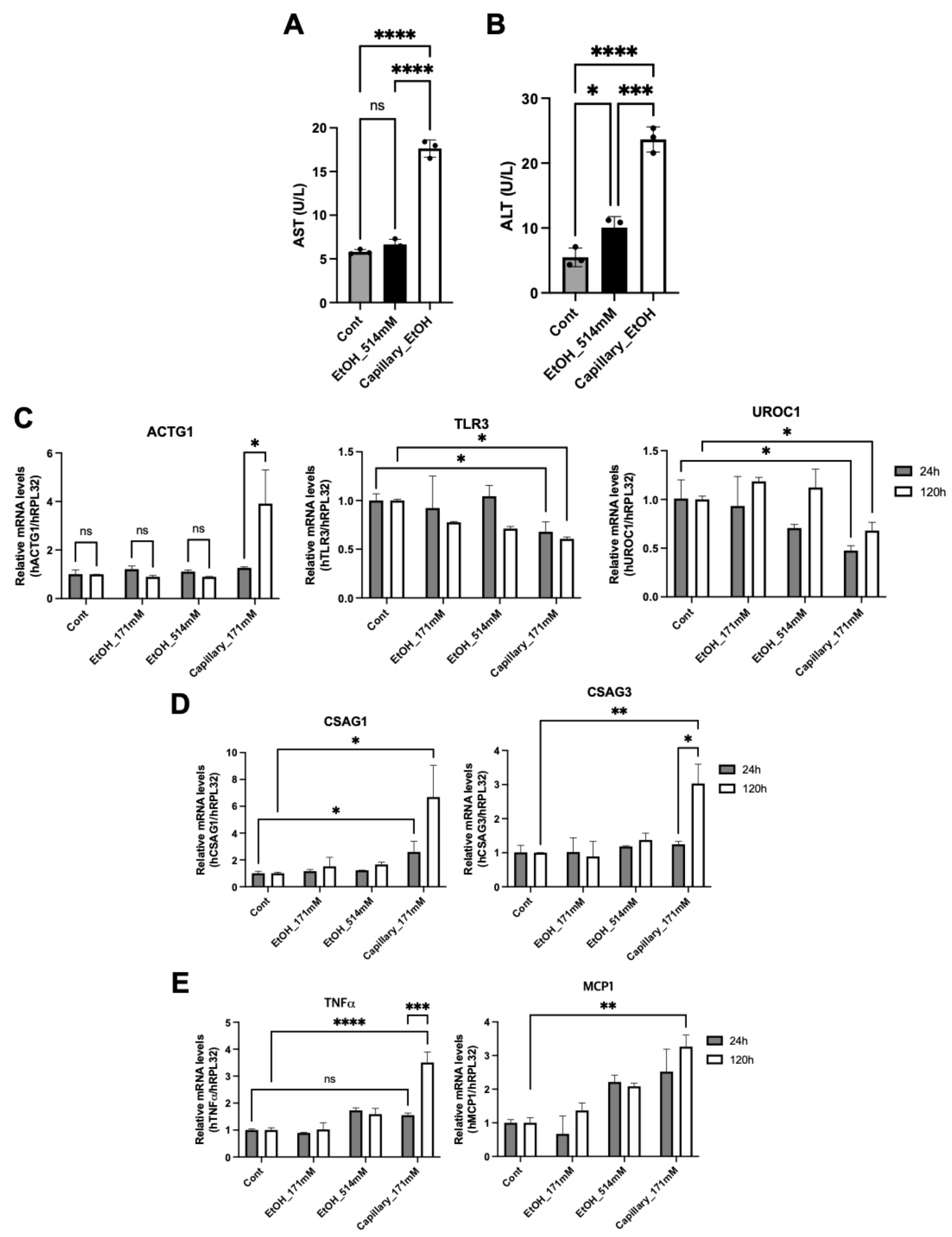
3.3. Long-Term Exposure of Ethanol Induces Hepatic Cytotoxicity
3.4. Ethanol Capillary System Shows a Different Hepatic Physiology
3.5. A Long-Term Exposure to Ethanol Induces Cellular Lipid Accumulation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- SAMHSA. Table 2.1B—Tobacco Product and Alcohol Use in Lifetime, Past Year, and Past Month among Persons Aged 12 or Older, by Age Group: Percentages, 2018 and 2019. Available online: https://www.samhsa.gov/data/sites/default/files/reports/rpt29394/NSDUHDetailedTabs2019/NSDUHDetTabsSect2pe2019.htm (accessed on 6 May 2021).
- CDC. Annual Average for United States 2011–2015 Alcohol-Attributable Deaths Due to Excessive Alcohol Use, All Ages, 2020. Alcohol and Public Health: Alcohol-Relat. Related Disease Impact (ARDI). Available online: https://nccd.cdc.gov/DPH_ARDI/Default/Report.aspx?T=AAM&P=1A04A664-0244-42C1-91DE-316F3AF6B447&R=B885BD06-13DF-45CD-8DD8-AA6B178C4ECE&M=32B5FFE7-81D2-43C5-A892-9B9B3C4246C7&L=&F=AAMCauseAgeGroupAllNew&D=H (accessed on 6 May 2021).
- National Highway Traffic Safety Administration. 2014 Crash Data Key Findings, 2015. Traffic Safety Facts Crash Stats. Available online: https://crashstats.nhtsa.dot.gov/Api/Public/ViewPublication/812219 (accessed on 6 May 2021).
- Shield, K.D.; Parry, C.; Rehm, J. Chronic diseases and conditions related to alcohol use. Alcohol Res. 2013, 35, 155–173. [Google Scholar]
- Dusak, A.; Onur, M.R.; Cicek, M.; Firat, U.; Ren, T.; Dogra, V.S. Radiological Imaging Features of Fasciola hepatica Infection–A Pictorial Review. J. Clin. Imaging Sci. 2012, 2, 2. [Google Scholar] [CrossRef]
- Stanaway, J.D.; Flaxman, A.D.; Naghavi, M.; Fitzmaurice, C.; Vos, T.; Abubakar, I.; Abu-Raddad, L.J.; Assadi, R.; Bhala, N.; Cowie, B.; et al. The global burden of viral hepatitis from 1990 to 2013: Findings from the Global Burden of Disease Study 2013. Lancet 2016, 388, 1081–1088. [Google Scholar] [CrossRef] [Green Version]
- Im, G.Y. Acute Alcoholic Hepatitis. Clin. Liver Dis. 2019, 23, 81–98. [Google Scholar] [CrossRef]
- Vuppalanchi, R.; Noureddin, M.; Alkhouri, N.; Sanyal, A.J. Therapeutic pipeline in nonalcoholic steatohepatitis. Nat. Rev. Gastroenterol. Hepatol. 2021. [Google Scholar] [CrossRef]
- Kisseleva, T.; Brenner, D. Molecular and cellular mechanisms of liver fibrosis and its regression. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, J.S. Alcohol, liver disease and the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 235–246. [Google Scholar] [CrossRef]
- Mathurin, P.; Bataller, R. Trends in the management and burden of alcoholic liver disease. J. Hepatol. 2015, 62, S38–S46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massey, V.L.; Arteel, G.E. Acute alcohol-induced liver injury. Front. Physiol. 2012, 3, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieber, C.S. Alcoholic fatty liver: Its pathogenesis and mechanism of progression to inflammation and fibrosis. Alcohol 2004, 34, 9–19. [Google Scholar] [CrossRef]
- Lucey, M.R.; Mathurin, P.; Morgan, T.R. Alcoholic hepatitis. N. Engl. J. Med. 2009, 360, 2758–2769. [Google Scholar] [CrossRef]
- Siegmund, S.V.; Dooley, S.; Brenner, D.A. Molecular mechanisms of alcohol-induced hepatic fibrosis. Dig. Dis. 2005, 23, 264–274. [Google Scholar] [CrossRef]
- Osna, N.A.; Donohue, T.M., Jr.; Kharbanda, K.K. Alcoholic Liver Disease: Pathogenesis and Current Management. Alcohol Res. 2017, 38, 147–161. [Google Scholar] [PubMed]
- Iida-Ueno, A.; Enomoto, M.; Tamori, A.; Kawada, N. Hepatitis B virus infection and alcohol consumption. World J. Gastroenterol. 2017, 23, 2651–2659. [Google Scholar] [CrossRef] [PubMed]
- Cromie, S.L.; Jenkins, P.J.; Bowden, D.S.; Dudley, F.J. Chronic hepatitis C: Effect of alcohol on hepatitic activity and viral titre. J. Hepatol. 1996, 25, 821–826. [Google Scholar] [CrossRef]
- Beier, J.I.; McClain, C.J. Mechanisms and cell signaling in alcoholic liver disease. Biol. Chem. 2010, 391, 1249–1264. [Google Scholar] [CrossRef] [Green Version]
- Louvet, A.; Mathurin, P. Alcoholic liver disease: Mechanisms of injury and targeted treatment. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Stickel, F.; Hampe, J. Genetic determinants of alcoholic liver disease. Gut 2012, 61, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Chang, B.; Li, X.; Zou, Z. Role of ALDH2 in Hepatic Disorders: Gene Polymorphism and Disease Pathogenesis. J. Clin. Transl. Hepatol. 2021, 9, 90–98. [Google Scholar] [CrossRef]
- Lu, Y.; Cederbaum, A.I. Cytochrome P450s and Alcoholic Liver Disease. Curr. Pharm. Des. 2018, 24, 1502–1517. [Google Scholar] [CrossRef]
- Sun, J.; Fu, J.; Li, L.; Chen, C.; Wang, H.; Hou, Y.; Xu, Y.; Pi, J. Nrf2 in alcoholic liver disease. Toxicol. Appl. Pharmacol. 2018, 357, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Dolganiuc, A.; Szabo, G. In vitro and in vivo models of acute alcohol exposure. World J. Gastroenterol. 2009, 15, 1168–1177. [Google Scholar] [CrossRef] [Green Version]
- Eysseric, H.; Gonthier, B.; Soubeyran, A.; Bessard, G.; Saxod, R.; Barret, L. There is not simple method to maintain a constant ethanol concentration in long-term cell culture: Keys to a solution applied to the survey of astrocytic ethanol absorption. Alcohol 1997, 14, 111–115. [Google Scholar] [CrossRef]
- Nath, V.; Reneau, J.C.; Dertien, J.S.; Agrawal, R.G.; Guerra, I.; Bhakta, Y.; Busari, K.; Neumann, M.K.; Bergeson, S.E.; Popp, R.L. An in vitro model for studying the effects of continuous ethanol exposure on N-methyl-D-aspartate receptor function. Alcohol 2012, 46, 3–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrasekaran, K.; Swaminathan, K.; Mathan Kumar, S.; Clemens, D.L.; Dey, A. In vitro evidence for chronic alcohol and high glucose mediated increased oxidative stress and hepatotoxicity. Alcohol Clin. Exp. Res. 2012, 36, 1004–1012. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Wang, S.; Ren, Z.; Frank, J.A.; Yang, X.H.; Zhang, Z.; Ke, Z.J.; Shi, X.; Luo, J. Chronic ethanol exposure enhances the aggressiveness of breast cancer: The role of p38gamma. Oncotarget 2016, 7, 3489–3505. [Google Scholar] [CrossRef] [Green Version]
- Jensen, K.P.; Lieberman, R.; Kranzler, H.R.; Gelernter, J.; Clinton, K.; Covault, J. Alcohol-responsive genes identified in human iPSC-derived neural cultures. Transl. Psychiatry 2019, 9, 96. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Meng, X.; Xu, J.; Huang, X.; Li, H.; Li, G.; Wang, S.; Man, Y.; Tang, W.; Li, J. GPR40 agonist ameliorates liver X receptor-induced lipid accumulation in liver by activating AMPK pathway. Sci. Rep. 2016, 6, 25237. [Google Scholar] [CrossRef] [Green Version]
- Mellott, M.B.; Searcy, K.; Pishko, M.V. Release of protein from highly cross-linked hydrogels of poly(ethylene glycol) diacrylate fabricated by UV polymerization. Biomaterials 2001, 22, 929–941. [Google Scholar] [CrossRef]
- Pochareddy, S.; Edenberg, H.J. Chronic alcohol exposure alters gene expression in HepG2 cells. Alcohol Clin. Exp. Res. 2012, 36, 1021–1033. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.H.; Li, W.L.; Jin, M.H.; Jin, Y.H.; Zhang, Y.Q.; Kong, L.Z.; Yu, L.Y.; Hyun, J.W.; Kwon, J.; Sun, H.N.; et al. Peroxiredoxin II Inhibits Alcohol-induced Apoptosis in L02 Hepatocytes Through AKT/beta-Catenin Signaling Pathway. Anticancer Res. 2020, 40, 4491–4504. [Google Scholar] [CrossRef]
- Wang, Y.; Tong, J.; Chang, B.; Wang, B.F.; Zhang, D.; Wang, B.Y. Effects of ethanol on the expression of caveolin-1 in HepG2 cells. Mol. Med. Rep. 2015, 11, 4409–4413. [Google Scholar] [CrossRef]
- Neuman, M.G.; Koren, G.; Tiribelli, C. In vitro assessment of the ethanol-induced hepatotoxicity on HepG2 cell line. Biochem. Biophys. Res. Commun. 1993, 197, 932–941. [Google Scholar] [CrossRef]
- Giannini, E.G.; Testa, R.; Savarino, V. Liver enzyme alteration: A guide for clinicians. CMAJ 2005, 172, 367–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, B.; Li, S.; Tan, Z.; Ma, L.; Liu, J. ACTG1 and TLR3 are biomarkers for alcohol-associated hepatocellular carcinoma. Oncol. Lett. 2019, 17, 1714–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Kang, C.; Li, N.; Liu, X.; Zhang, J.; Gao, F.; Dai, L. Identification of special key genes for alcohol-related hepatocellular carcinoma through bioinformatic analysis. PeerJ 2019, 7, e6375. [Google Scholar] [CrossRef] [Green Version]
- Yin, M.; Wheeler, M.D.; Kono, H.; Bradford, B.U.; Gallucci, R.M.; Luster, M.I.; Thurman, R.G. Essential role of tumor necrosis factor alpha in alcohol-induced liver injury in mice. Gastroenterology 1999, 117, 942–952. [Google Scholar] [CrossRef]
- Mandrekar, P.; Ambade, A.; Lim, A.; Szabo, G.; Catalano, D. An essential role for monocyte chemoattractant protein-1 in alcoholic liver injury: Regulation of proinflammatory cytokines and hepatic steatosis in mice. Hepatology 2011, 54, 2185–2197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keegan, A.; Martini, R.; Batey, R. Ethanol-related liver injury in the rat: A model of steatosis, inflammation and pericentral fibrosis. J. Hepatol. 1995, 23, 591–600. [Google Scholar] [CrossRef]
- Altamirano, J.; Bataller, R. Alcoholic liver disease: Pathogenesis and new targets for therapy. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 491–501. [Google Scholar] [CrossRef]
- Benoist, F.; Grand-Perret, T. ApoB-100 secretion by HepG2 cells is regulated by the rate of triglyceride biosynthesis but not by intracellular lipid pools. Arter. Thromb. Vasc. Biol. 1996, 16, 1229–1235. [Google Scholar] [CrossRef]
- Meex, S.J.; Andreo, U.; Sparks, J.D.; Fisher, E.A. Huh-7 or HepG2 cells: Which is the better model for studying human apolipoprotein-B100 assembly and secretion? J. Lipid Res. 2011, 52, 152–158. [Google Scholar] [CrossRef] [Green Version]
- Allister, E.M.; Mulvihill, E.E.; Barrett, P.H.; Edwards, J.Y.; Carter, L.P.; Huff, M.W. Inhibition of apoB secretion from HepG2 cells by insulin is amplified by naringenin, independent of the insulin receptor. J. Lipid Res. 2008, 49, 2218–2229. [Google Scholar] [CrossRef] [Green Version]
- Neumann, O.; Kesselmeier, M.; Geffers, R.; Pellegrino, R.; Radlwimmer, B.; Hoffmann, K.; Ehemann, V.; Schemmer, P.; Schirmacher, P.; Lorenzo Bermejo, J.; et al. Methylome analysis and integrative profiling of human HCCs identify novel protumorigenic factors. Hepatology 2012, 56, 1817–1827. [Google Scholar] [CrossRef]
- Mazzoccoli, J.P.; Feke, D.L.; Baskaran, H.; Pintauro, P.N. Mechanical and cell viability properties of crosslinked low- and high-molecular weight poly(ethylene glycol) diacrylate blends. J. Biomed. Mater. Res. A 2010, 93, 558–566. [Google Scholar] [CrossRef] [Green Version]
- Rolla, R.; Vay, D.; Mottaran, E.; Parodi, M.; Traverso, N.; Arico, S.; Sartori, M.; Bellomo, G.; Klassen, L.W.; Thiele, G.M.; et al. Detection of circulating antibodies against malondialdehyde-acetaldehyde adducts in patients with alcohol-induced liver disease. Hepatology 2000, 31, 878–884. [Google Scholar] [CrossRef]
- McKillop, I.H.; Schrum, L.W.; Thompson, K.J. Role of alcohol in the development and progression of hepatocellular carcinoma. Hepat. Oncol. 2016, 3, 29–43. [Google Scholar] [CrossRef]
- Petrasek, J.; Csak, T.; Ganz, M.; Szabo, G. Differences in innate immune signaling between alcoholic and non-alcoholic steatohepatitis. J. Gastroenterol. Hepatol. 2013, 28, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Szabo, G.; Petrasek, J.; Bala, S. Innate immunity and alcoholic liver disease. Dig. Dis. 2012, 30, 55–60. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, W.; Jeong, H.-S.; Kim, S.-C.; Choi, C.-H.; Lee, K.-H. Chronic Alcohol Exposure of Cells Using Controlled Alcohol-Releasing Capillaries. Cells 2021, 10, 1120. https://doi.org/10.3390/cells10051120
Kim W, Jeong H-S, Kim S-C, Choi C-H, Lee K-H. Chronic Alcohol Exposure of Cells Using Controlled Alcohol-Releasing Capillaries. Cells. 2021; 10(5):1120. https://doi.org/10.3390/cells10051120
Chicago/Turabian StyleKim, Wanil, Hye-Seon Jeong, Sang-Chan Kim, Chang-Hyung Choi, and Kyung-Ha Lee. 2021. "Chronic Alcohol Exposure of Cells Using Controlled Alcohol-Releasing Capillaries" Cells 10, no. 5: 1120. https://doi.org/10.3390/cells10051120