
The Role of Galectin-9 as Mediator of Atopic Dermatitis: Effect on Keratinocytes

 , , , and
, , , and 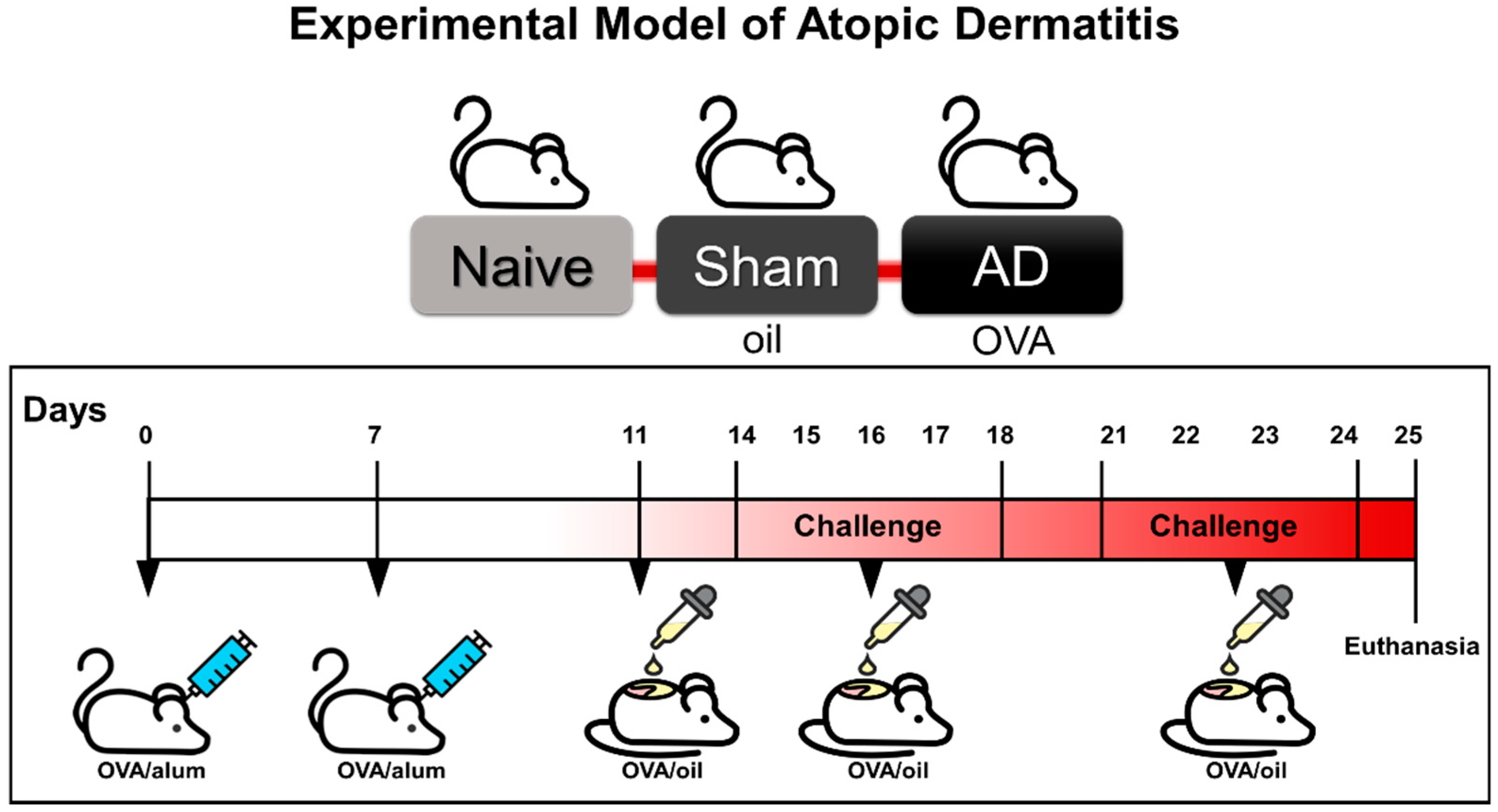{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Model of Atopic Dermatitis
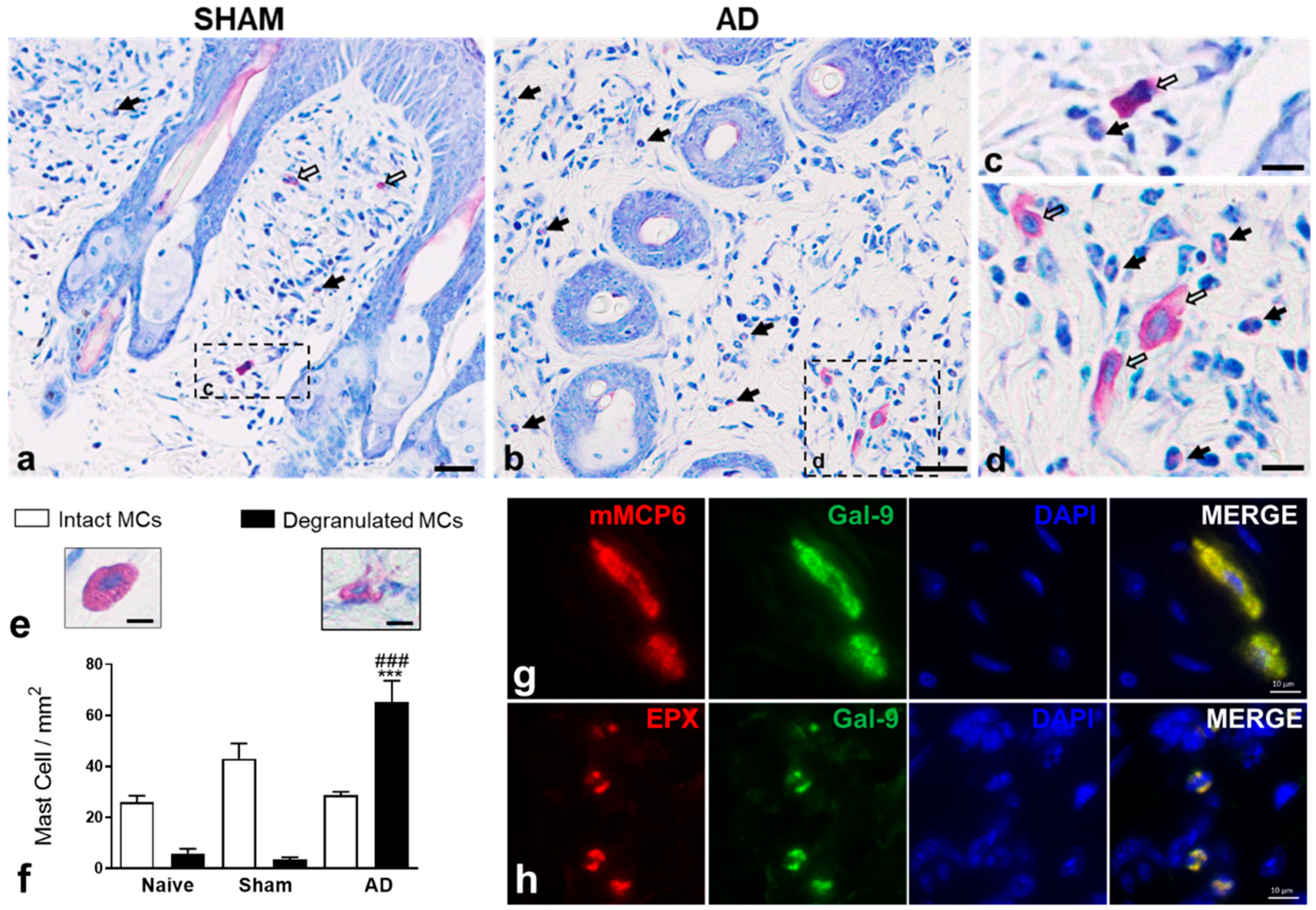
2.2. Histological Analysis and Quantification of Mast Cells in Skin
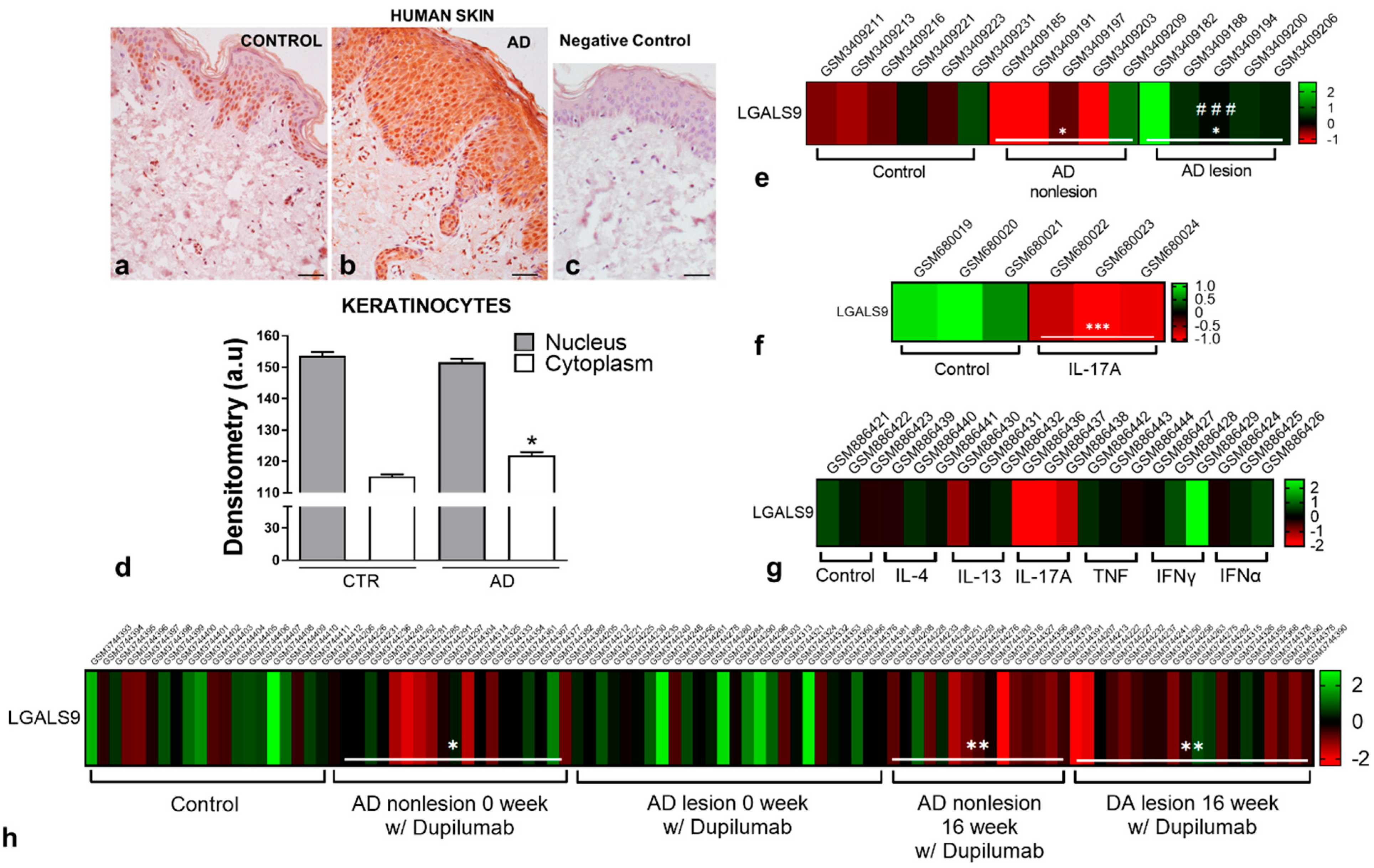
2.3. Human Skin Biopsies
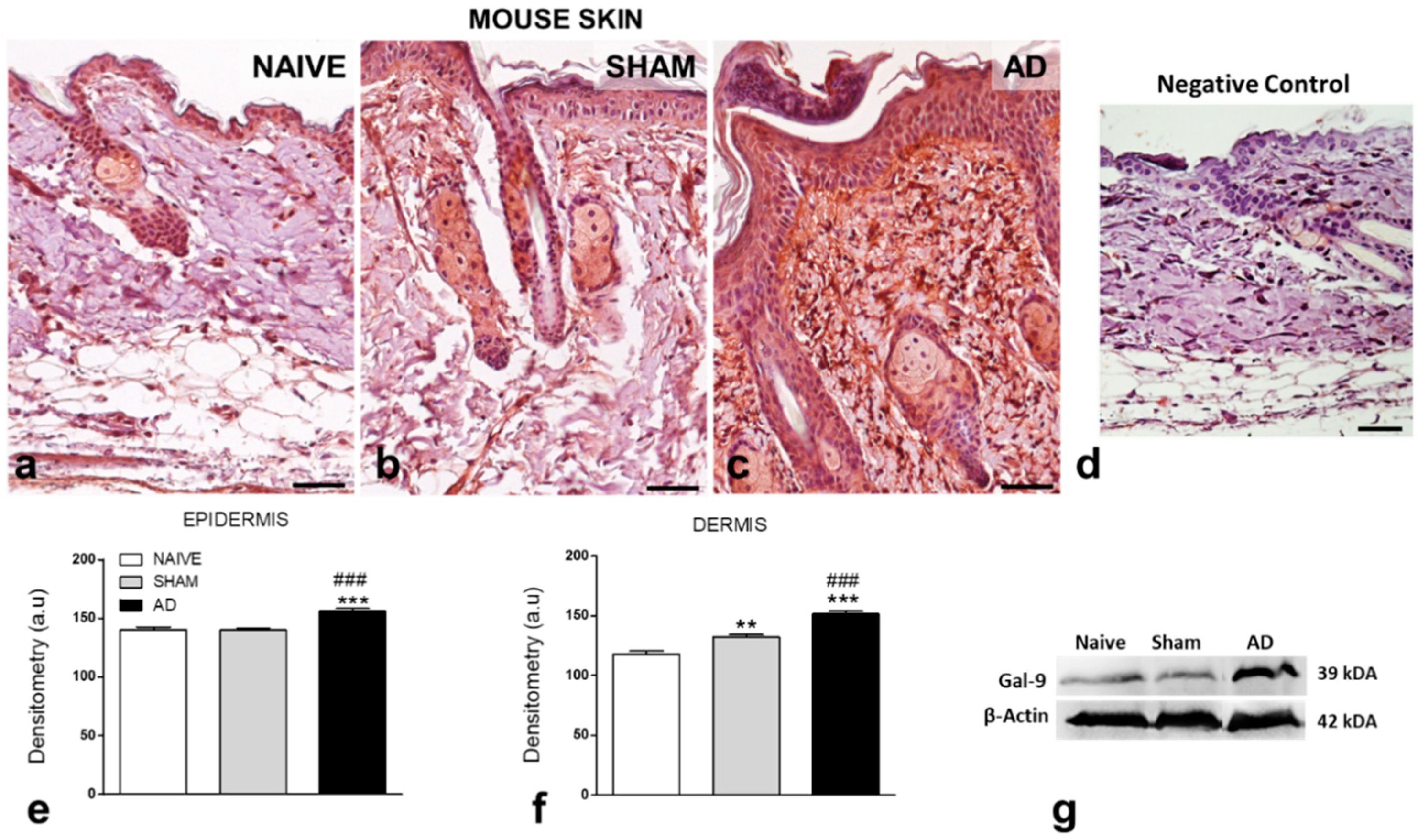
2.4. Galectin-9 Levels: Immunohistochemistry and Western Blotting
2.5. Immunofluorescence
2.6. Human Keratinocyte Culture and Treatments
2.7. Proinflammatory Cytokine and RANTES/CCL5 Levels
2.8. Cell Proliferation Assay
2.9. Scratch-Wound Assay
2.10. Bioinformatic Analysis
2.11. Statistical Analysis
3. Results
3.1. Gal-9 Levels Are Upregulated in Murine and Human AD
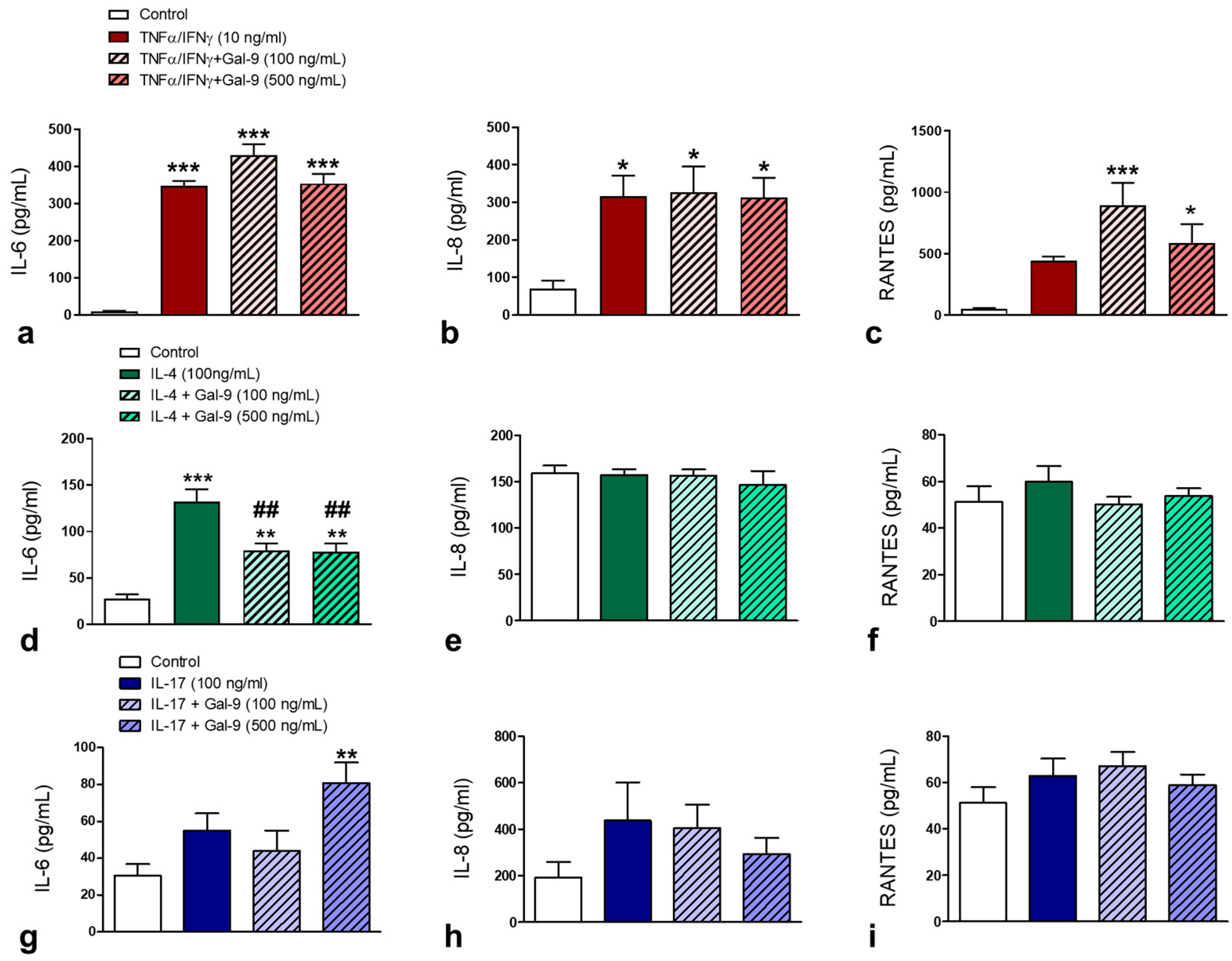
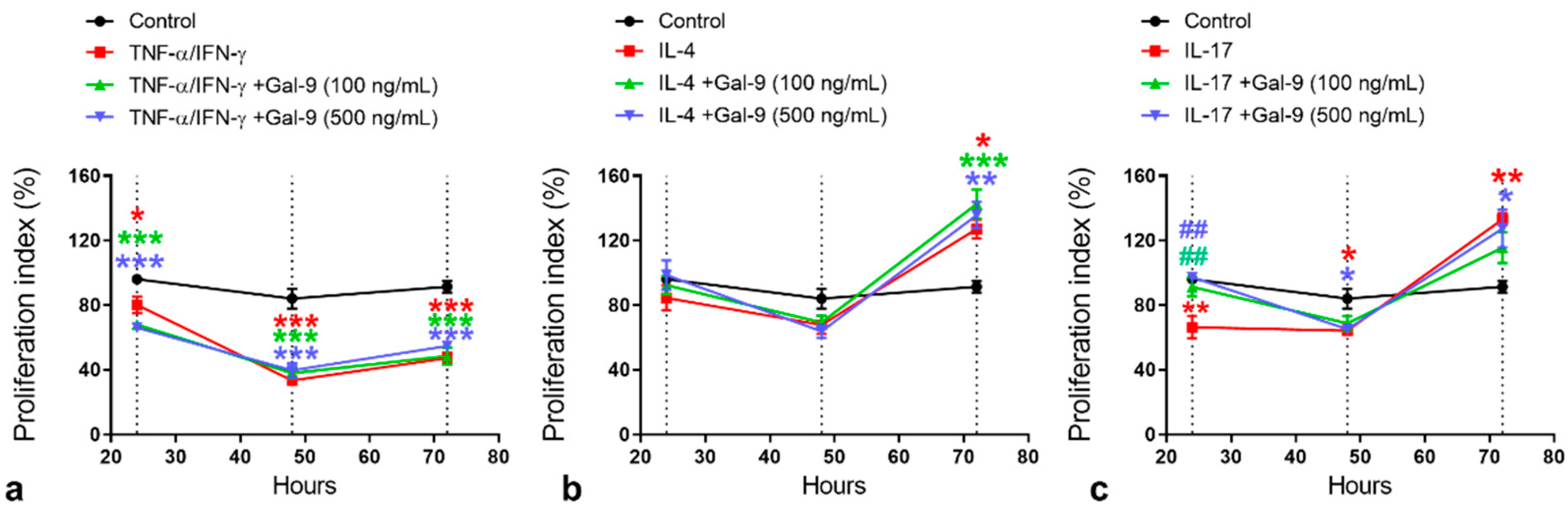
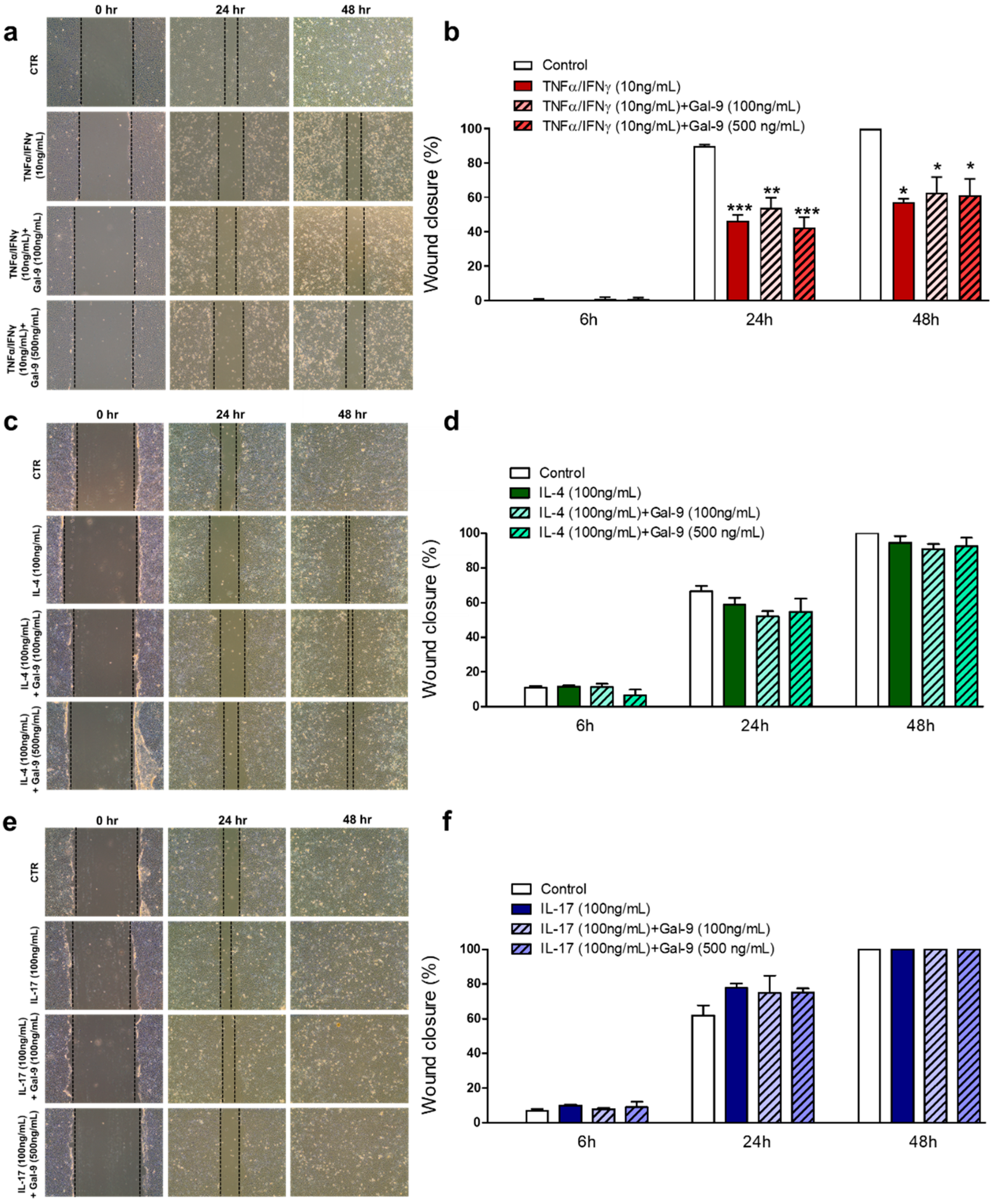
3.2. Effect of Exogenous Administration of Gal-9 on Keratinocytes: Cytokine Release, Proliferation and Migration Rates
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leung, D.Y.; Guttman-Yassky, E. Deciphering the complexities of atopic dermatitis: Shifting paradigms in treatment approaches. J. Allergy Clin. Immunol. 2014, 134, 769–779. [Google Scholar] [CrossRef] [Green Version]
- Guttman-Yassky, E.; Krueger, J.G. Atopic dermatitis and psoriasis: Two different immune diseases or one spectrum? Curr. Opin. Immunol. 2017, 48, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Dainichi, T.; Kitoh, A.; Otsuka, A.; Nakajima, S.; Nomura, T.; Kaplan, D.H.; Kabashima, K. The epithelial immune microenvironment (EIME) in atopic dermatitis and psoriasis. Nat. Immunol. 2018, 19, 1286–1298. [Google Scholar] [CrossRef] [PubMed]
- Elola, M.T.; Ferragut, F.; Méndez-Huergo, S.P.; Croci, D.O.; Bracalente, C.; Rabinovich, G.A. Galectins: Multitask signaling molecules linking fibroblast, endothelial and immune cell programs in the tumor microenvironment. Cell Immunol. 2018, 333, 34–45. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.T.; Rabinovich, G.A. Galectins: Regulators of acute and chronic inflammation. Ann. N. Y. Acad. Sci. 2010, 1183, 158–182. [Google Scholar] [CrossRef]
- Nakajima, R.; Miyagaki, T.; Oka, T.; Nakao, M.; Kawaguchi, M.; Suga, H.; Morimura, S.; Kai, H.; Asano, Y.; Tada, Y.; et al. Elevated serum galectin-9 levels in patients with atopic dermatitis. J. Dermatol. 2015, 42, 723–726. [Google Scholar] [CrossRef]
- Niwa, H.; Satoh, T.; Matsushima, Y.; Hosoya, K.; Saeki, K.; Niki, T.; Hirashima, M.; Yokozeki, H. Stable form of galectin-9, a Tim-3 ligand, inhibits contact hypersensitivity and psoriatic reactions: A potent therapeutic tool for Th1- and/or Th17-mediated skin inflammation. Clin. Immunol. 2009, 132, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Cedeno-Laurent, F.; Barthel, S.R.; Opperman, M.J.; Lee, D.M.; Clark, R.A.; Dimitroff, C.J. Development of a nascent galectin-1 chimeric molecule for studying the role of leukocyte galectin-1 ligands and immune disease modulation. J. Immunol. 2010, 185, 4659–4672. [Google Scholar] [CrossRef] [Green Version]
- Niki, T.; Tsutsui, S.; Hirose, S.; Aradono, S.; Sugimoto, Y.; Takeshita, K.; Nishi, N.; Hirashima, M. Galectin-9 is a high affinity IgE-binding lectin with anti-allergic effect by blocking IgE-antigen complex formation. J. Biol. Chem. 2009, 284, 32344–32352. [Google Scholar] [CrossRef] [Green Version]
- Kojima, R.; Ohno, T.; Iikura, M.; Niki, T.; Hirashima, M.; Iwaya, K.; Tsuda, H.; Nonoyama, S.; Matsuda, A.; Saito, H.; et al. Galectin-9 enhances cytokine secretion, but suppresses survival and degranulation, in human mast cell line. PLoS ONE 2014, 9, e86106. [Google Scholar] [CrossRef]
- Sato, M.; Nishi, N.; Shoji, H.; Seki, M.; Hashidate, T.; Hirabayashi, J.; Kasai Ki, K.; Hata, Y.; Suzuki, S.; Hirashima, M.; et al. Functional analysis of the carbohydrate recognition domains and a linker peptide of galectin-9 as to eosinophil chemoattractant activity. Glycobiology 2002, 12, 191–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igawa, K.; Satoh, T.; Hirashima, M.; Yokozeki, H. Regulatory mechanisms of galectin-9 and eotaxin-3 synthesis in epidermal keratinocytes: Possible involvement of galectin-9 in dermal eosinophilia of Th1-polarized skin inflammation. Allergy 2006, 61, 1385–1391. [Google Scholar] [CrossRef]
- Corrêa, M.P.; Andrade, F.E.C.; Gimenes, A.D.; Gil, C.D. Anti-inflammatory effect of galectin-1 in a murine model of atopic dermatitis. J. Mol. Med. Berl. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, L.A.; Thaçi, D.; Hamilton, J.D.; Graham, N.M.; Bieber, T.; Rocklin, R.; Ming, J.E.; Ren, H.; Kao, R.; Simpson, E.; et al. Dupilumab treatment in adults with moderate-to-severe atopic dermatitis. N. Engl. J. Med. 2014, 371, 130–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.; Qi, F.; Gong, Y.; Zhang, J.; Zhu, B. Biological Therapies for Atopic Dermatitis: A Systematic Review. Dermatology 2021, 1–11. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, Y.J.; Kang, M.J.; Seo, J.H.; Kim, H.Y.; Jeong, S.K.; Lee, S.H.; Kim, J.M.; Hong, S.J. A novel mouse model of atopic dermatitis with epicutaneous allergen sensitization and the effect of Lactobacillus rhamnosus. Exp. Dermatol. 2012, 21, 672–675. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, H.; Maki, N.; Yoshida, S.; Arai, M.; Wang, J.; Oikawa, Y.; Ikeda, T.; Hirota, N.; Nakagawa, H.; Ishii, A. A mouse model of the atopic eczema/dermatitis syndrome by repeated application of a crude extract of house-dust mite Dermatophagoides farinae. Allergy 2003, 58, 139–145. [Google Scholar] [CrossRef]
- Kim, H.; Kim, J.R.; Kang, H.; Choi, J.; Yang, H.; Lee, P.; Kim, J.; Lee, K.W. 7,8,4’-Trihydroxyisoflavone attenuates DNCB-induced atopic dermatitis-like symptoms in NC/Nga mice. PLoS ONE 2014, 9, e104938. [Google Scholar] [CrossRef]
- Heo, W.I.; Lee, K.E.; Hong, J.Y.; Kim, M.N.; Oh, M.S.; Kim, Y.S.; Kim, K.W.; Kim, K.E.; Sohn, M.H. The role of interleukin-17 in mouse models of atopic dermatitis and contact dermatitis. Clin. Exp. Dermatol. 2015, 40, 665–671. [Google Scholar] [CrossRef]
- Miyazaki, D.; Tominaga, T.; Yakura, K.; Kuo, C.H.; Komatsu, N.; Inoue, Y.; Ono, S.J. Conjunctival mast cell as a mediator of eosinophilic response in ocular allergy. Mol. Vis. 2008, 14, 1525–1532. [Google Scholar]
- Fukuda, K.; Ohbayashi, M.; Morohoshi, K.; Zhang, L.; Liu, F.T.; Ono, S.J. Critical role of IgE-dependent mast cell activation in a murine model of allergic conjunctivitis. J. Allergy Clin. Immunol. 2009, 124, 827–833.e2. [Google Scholar] [CrossRef]
- Saita, N.; Goto, E.; Yamamoto, T.; Cho, I.; Tsumori, K.; Kohrogi, H.; Maruo, K.; Ono, T.; Takeya, M.; Kashio, Y.; et al. Association of galectin-9 with eosinophil apoptosis. Int. Arch. Allergy Immunol. 2002, 128, 42–50. [Google Scholar] [CrossRef]
- Yamamoto, H.; Kashio, Y.; Shoji, H.; Shinonaga, R.; Yoshimura, T.; Nishi, N.; Nabe, T.; Nakamura, T.; Kohno, S.; Hirashima, M. Involvement of galectin-9 in guinea pig allergic airway inflammation. Int. Arch. Allergy Immunol. 2007, 143 (Suppl. 1), 95–105. [Google Scholar] [CrossRef]
- Sziksz, E.; Kozma, G.T.; Pállinger, E.; Komlósi, Z.I.; Adori, C.; Kovács, L.; Szebeni, B.; Rusai, K.; Losonczy, G.; Szabó, A.; et al. Galectin-9 in allergic airway inflammation and hyper-responsiveness in mice. Int. Arch. Allergy Immunol. 2010, 151, 308–317. [Google Scholar] [CrossRef]
- Katoh, S.; Shimizu, H.; Obase, Y.; Oomizu, S.; Niki, T.; Ikeda, M.; Mouri, K.; Kobashi, Y.; Hirashima, M.; Oka, M. Preventive effect of galectin-9 on double-stranded RNA-induced airway hyperresponsiveness in an exacerbation model of mite antigen-induced asthma in mice. Exp. Lung Res. 2013, 39, 453–462. [Google Scholar] [CrossRef]
- Farag, A.G.A.; Al-Sharaky, D.R.; Allam, S.S.; Khaled, H.N. Role of Galectin-9 in Atopic Dermatitis—Is It Mediated Through E Selectin? A Clinical and Immunohistochemical Study. Clin. Cosmet Investig. Dermatol. 2020, 13, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Guttman-Yassky, E.; Bissonnette, R.; Ungar, B.; Suárez-Fariñas, M.; Ardeleanu, M.; Esaki, H.; Suprun, M.; Estrada, Y.; Xu, H.; Peng, X.; et al. Dupilumab progressively improves systemic and cutaneous abnormalities in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2019, 143, 155–172. [Google Scholar] [CrossRef] [Green Version]
- De Kivit, S.; Saeland, E.; Kraneveld, A.D.; van de Kant, H.J.; Schouten, B.; van Esch, B.C.; Knol, J.; Sprikkelman, A.B.; van der Aa, L.B.; Knippels, L.M.; et al. Galectin-9 induced by dietary synbiotics is involved in suppression of allergic symptoms in mice and humans. Allergy 2012, 67, 343–352. [Google Scholar] [CrossRef]
- Kim, H.W.; Ju, D.B.; Kye, Y.C.; Ju, Y.J.; Kim, C.G.; Lee, I.K.; Park, S.M.; Choi, I.S.; Cho, K.K.; Lee, S.H.; et al. Galectin-9 Induced by Dietary Probiotic Mixture Regulates Immune Balance to Reduce Atopic Dermatitis Symptoms in Mice. Front Immunol. 2019, 10, 3063. [Google Scholar] [CrossRef]
- Dai, S.Y.; Nakagawa, R.; Itoh, A.; Murakami, H.; Kashio, Y.; Abe, H.; Katoh, S.; Kontani, K.; Kihara, M.; Zhang, S.L.; et al. Galectin-9 induces maturation of human monocyte-derived dendritic cells. J. Immunol. 2005, 175, 2974–2981. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, M.; Katoh, S.; Shimizu, H.; Hasegawa, A.; Ohashi-Doi, K.; Oka, M. Beneficial effects of Galectin-9 on allergen-specific sublingual immunotherapy in a Dermatophagoides farinae-induced mouse model of chronic asthma. Allergol. Int. 2017, 66, 432–439. [Google Scholar] [CrossRef] [Green Version]
- Katoh, S.; Ishii, N.; Nobumoto, A.; Takeshita, K.; Dai, S.Y.; Shinonaga, R.; Niki, T.; Nishi, N.; Tominaga, A.; Yamauchi, A.; et al. Galectin-9 inhibits CD44-hyaluronan interaction and suppresses a murine model of allergic asthma. Am. J. Respir. Crit. Care Med. 2007, 176, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Diehl, S.; Rincón, M. The two faces of IL-6 on Th1/Th2 differentiation. Mol. Immunol. 2002, 39, 531–536. [Google Scholar] [CrossRef]
- Chieosilapatham, P.; Kiatsurayanon, C.; Umehara, Y.; Trujillo-Paez, J.V.; Peng, G.; Yue, H.; Nguyen, L.T.H.; Niyonsaba, F. Keratinocytes: Innate immune cells in atopic dermatitis. Clin. Exp. Immunol. 2021. [Google Scholar] [CrossRef]
- Nakajima, S.; Kitoh, A.; Egawa, G.; Natsuaki, Y.; Nakamizo, S.; Moniaga, C.S.; Otsuka, A.; Honda, T.; Hanakawa, S.; Amano, W.; et al. IL-17A as an inducer for Th2 immune responses in murine atopic dermatitis models. J. Investig. Dermatol. 2014, 134, 2122–2130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asahina, R.; Maeda, S. A review of the roles of keratinocyte-derived cytokines and chemokines in the pathogenesis of atopic dermatitis in humans and dogs. Vet. Dermatol. 2017, 28, 16.e15. [Google Scholar] [CrossRef]
- Kameyoshi, Y.; Dörschner, A.; Mallet, A.I.; Christophers, E.; Schröder, J.M. Cytokine RANTES released by thrombin-stimulated platelets is a potent attractant for human eosinophils. J. Exp. Med. 1992, 176, 587–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.T.; Goodarzi, H.; Chen, H.Y. IgE, mast cells, and eosinophils in atopic dermatitis. Clin. Rev. Allergy Immunol. 2011, 41, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Grewe, M.; Czech, W.; Morita, A.; Werfel, T.; Klammer, M.; Kapp, A.; Ruzicka, T.; Schöpf, E.; Krutmann, J. Human eosinophils produce biologically active IL-12: Implications for control of T cell responses. J. Immunol. 1998, 161, 415–420. [Google Scholar] [PubMed]
- Kasamatsu, A.; Uzawa, K.; Nakashima, D.; Koike, H.; Shiiba, M.; Bukawa, H.; Yokoe, H.; Tanzawa, H. Galectin-9 as a regulator of cellular adhesion in human oral squamous cell carcinoma cell lines. Int. J. Mol. Med. 2005, 16, 269–273. [Google Scholar] [CrossRef]
- Brunner, P.M.; Guttman-Yassky, E.; Leung, D.Y. The immunology of atopic dermatitis and its reversibility with broad-spectrum and targeted therapies. J. Allergy Clin. Immunol. 2017, 139, S65–S76. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corrêa, M.P.; Areias, L.L.; Correia-Silva, R.D.; D’Ávila, S.C.G.P.; Leopoldino, A.M.; Greco, K.V.; Gil, C.D. The Role of Galectin-9 as Mediator of Atopic Dermatitis: Effect on Keratinocytes. Cells 2021, 10, 947. https://doi.org/10.3390/cells10040947
Corrêa MP, Areias LL, Correia-Silva RD, D’Ávila SCGP, Leopoldino AM, Greco KV, Gil CD. The Role of Galectin-9 as Mediator of Atopic Dermatitis: Effect on Keratinocytes. Cells. 2021; 10(4):947. https://doi.org/10.3390/cells10040947
Chicago/Turabian StyleCorrêa, Mab P., Libnah L. Areias, Rebeca D. Correia-Silva, Solange C. G. P. D’Ávila, Andréia M. Leopoldino, Karin V. Greco, and Cristiane D. Gil. 2021. "The Role of Galectin-9 as Mediator of Atopic Dermatitis: Effect on Keratinocytes" Cells 10, no. 4: 947. https://doi.org/10.3390/cells10040947