
Phosphorylation of Kindlins and the Control of Integrin Function

Abstract
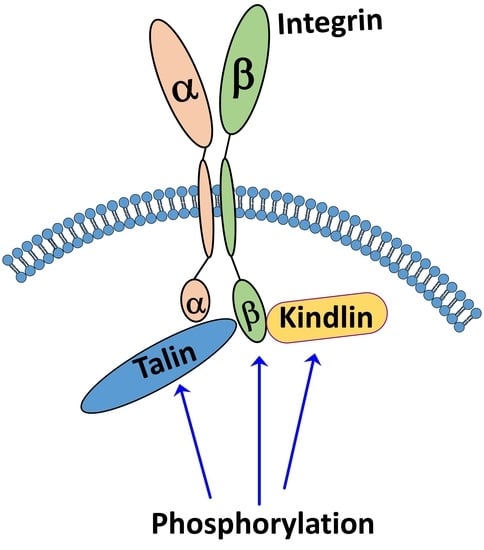
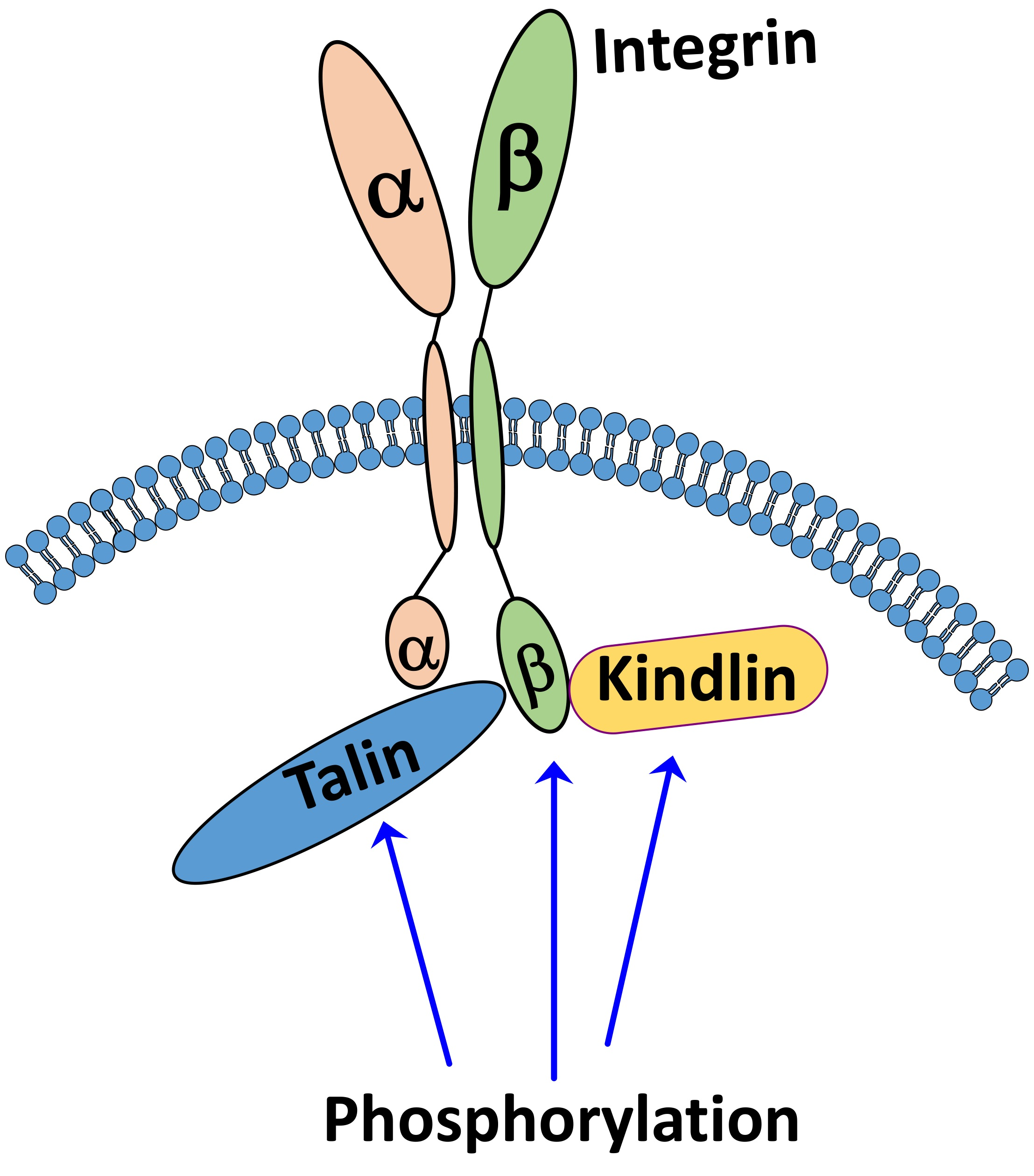
:
1. Introduction
1.1. Kindlin Structure and Its Role in Integrin Activation
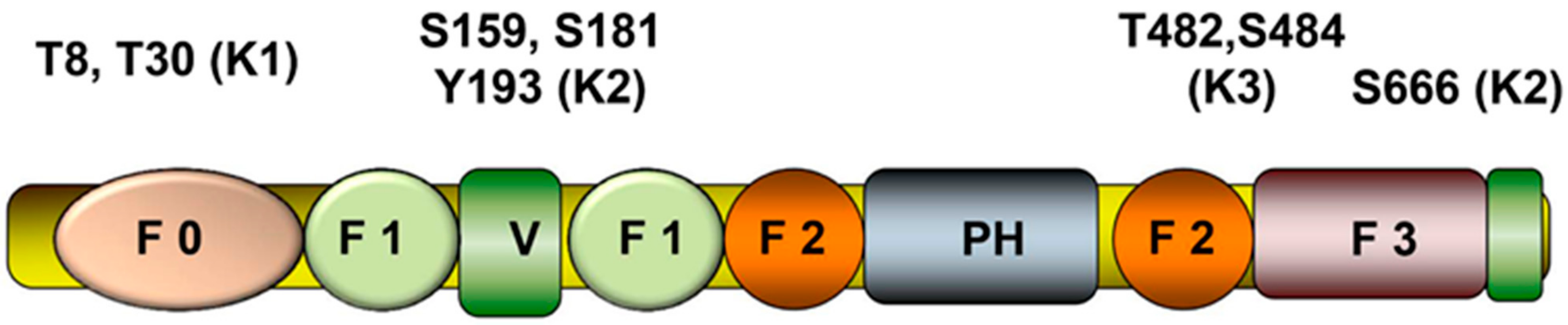
1.2. Phosphorylation of Kindlins
2. Other Post-Translational Modifications of Kindlins and Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ginsberg, M.H.; Du, X.; Plow, E.F. Inside-out signalling. Curr. Opin. Cell Biol. 1992, 4, 766–771. [Google Scholar] [CrossRef]
- Li, Z.; Delaney, M.K.; O’Brien, K.A.; Du, X. Signaling During Platelet Adhesion and Activation. Arter. Thromb. Vasc. Biol. 2010, 30, 2341–2349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durrant, T.N.; van den Bosch, M.T.; Hers, I. Integrin alphaIIbbeta3 outside-in signaling. Blood 2017, 130, 1607–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginsberg, M.H. Integrin activation. BMB Rep. 2014, 47, 655–659. [Google Scholar] [CrossRef]
- Humphries, J.D.; Chastney, M.R.; A Askari, J.; Humphries, M.J. Signal transduction via integrin adhesion complexes. Curr. Opin. Cell Biol. 2019, 56, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Horton, E.R.; Byron, A.; Askari, J.A.; Ng, D.H.J.; Millon-Frémillon, A.; Robertson, J.; Koper, E.J.; Paul, N.R.; Warwood, S.; Knight, D.P.; et al. Definition of a consensus integrin adhesome and its dynamics during adhesion complex assembly and disassembly. Nat. Cell Biol. 2015, 17, 1577–1587. [Google Scholar] [CrossRef] [Green Version]
- Vinogradova, O.; Velyvis, A.; Velyviene, A.; Hu, B.; Haas, T.A.; Plow, E.F.; Qin, J. A structural mechanism of integrin aIIbb3 “inside-out” activation as regulated by its cytoplasmic face. Cell 2002, 110, 587–597. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Plow, E.F.; Qin, J. Initiation of focal adhesion assembly by talin and kindlin: A dynamic view. Protein Sci. 2021, 30, 531–542. [Google Scholar] [CrossRef]
- Zhu, L.; Liu, H.; Lu, F.; Yang, J.; Byzova, T.V.; Qin, J. Structural Basis of Paxillin Recruitment by Kindlin-2 in Regulating Cell Adhesion. Structure 2019, 27, 1686–1697.e5. [Google Scholar] [CrossRef]
- Gao, J.; Huang, M.; Lai, J.; Mao, K.; Sun, P.; Cao, Z.; Hu, Y.; Zhang, Y.; Schulte, M.L.; Jin, C.; et al. Kindlin supports platelet integrin alphaIIbbeta3 activation by interacting with paxillin. J. Cell Sci. 2017, 130, 3764–3775. [Google Scholar] [CrossRef] [Green Version]
- Kasirer-Friede, A.; Kang, J.; Kahner, B.; Ye, F.; Ginsberg, M.H.; Shattil, S.J. ADAP interactions with talin and kindlin promote platelet integrin alphaIIbbeta3 activation and stable fibrinogen binding. Blood 2014, 20, 3156–3165. [Google Scholar] [CrossRef] [Green Version]
- Das, M.; Ithychanda, S.S.; Qin, J.; Plow, E.F. Migfilin and Filamin as Regulators of Integrin Activation in Endothelial Cells and Neutrophils. PLoS ONE 2011, 6, e26355. [Google Scholar] [CrossRef]
- Bouvard, D.; Aszodi, A.; Kostka, G.; Block, M.R.; Albigès-Rizo, C.; Fässler, R. Defective osteoblast function in ICAP-1-deficient mice. Development 2007, 134, 2615–2625. [Google Scholar] [CrossRef] [Green Version]
- Ithychanda, S.S.; Qin, J. Evidence for Multisite Ligand Binding and Stretching of Filamin by Integrin and Migfilin. Biochemistry 2011, 50, 4229–4231. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Das, M.; Yang, J.; Ithychanda, S.S.; Yakubenko, V.P.; Plow, E.F.; Qin, J. Structural mechanism of integrin inactivation by filamin. Nat. Struct. Mol. Biol. 2015, 22, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Rantala, J.K.; Pouwels, J.; Pellinen, T.; Veltel, S.; Laasola, P.; Mattila, E.; Potter, C.S.; Duffy, T.; Sundberg, J.P.; Kallioniemi, O.; et al. SHARPIN is an endogenous inhibitor of beta1-integrin activation. Nat. Cell Biol. 2011, 13, 1315–1324. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Bao, Y.; Ge, S.; Sun, P.; Sun, J.; Liu, J.; Chen, F.; Han, L.; Cao, Z.; Qin, J.; et al. Sharpin suppresses beta1-integrin activation by complexing with the beta1 tail and kindlin-1. Cell Commun. Signal. 2019, 17, 101. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Rose, D.M.; Woodside, D.G.; Goldfinger, L.E.; Ginsberg, M.H. Integrin alpha 4 beta 1-dependent T cell migration requires both phosphorylation and dephosphorylation of the alpha 4 cytoplasmic domain to regulate the reversible binding of paxillin. J. Biol. Chem. 2003, 278, 34845–34853. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, A.L.; Nannizzi-Alaimo, L.; Silver, D.; Sellers, J.R.; Ginsberg, M.H.; Law, D.A.; Phillips, D.R. Tyrosine phosphorylation of the b3 cytoplasmic domain mediates integrin-cytoskeletal interactions. J. Biol. Chem. 1998, 273, 13878–13885. [Google Scholar] [CrossRef] [Green Version]
- Gahmberg, C.G.; Gronholm, M.; Uotila, L.M. Regulation of integrin activity by phosphorylation. Adv. Exp. Med. Biol. 2014, 819, 85–96. [Google Scholar]
- Phillips, D.R.; Nannizzi-Alaimo, L.; Prasad, K.S. Beta3 tyrosine phosphorylation in alphaIIbbeta3 (platelet membrane GP IIb-IIIa) outside-in integrin signaling. Thromb. Haemost. 2001, 86, 246–258. [Google Scholar]
- Mitra, S.K.; Schlaepfer, D.D. Integrin-regulated FAK-Src signaling in normal and cancer cells. Curr. Opin. Cell Biol. 2006, 18, 516–523. [Google Scholar] [CrossRef]
- Turner, E.C.; Pavalko, F.M.; Burridge, K. The role of phosphorylation and limited proteolytic cleavage of talin and vinculin in the disruption of focal adhesion integrity. J. Biol. Chem. 1989, 264, 11938–11944. [Google Scholar] [CrossRef]
- Ratnikov, B.; Ptak, C.; Han, J.; Shabanowitz, J.; Hunt, D.F.; Ginsberg, M.H. Talin phosphorylation sites mapped by mass spectrometry. J. Cell Sci. 2005, 118, 4921–4923. [Google Scholar] [CrossRef] [Green Version]
- Larjava, H.; Plow, E.F.; Wu, C. Kindlins: Essential regulators of integrin signalling and cell–matrix adhesion. EMBO Rep. 2008, 9, 1203–1208. [Google Scholar] [CrossRef]
- Rognoni, E.; Ruppert, R.; Fässler, R. The kindlin family: Functions, signaling properties and implications for human disease. J. Cell Sci. 2016, 129, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Malinin, N.L.; Plow, E.F.; Byzova, T.V. Kindlins in FERM adhesion. Blood 2010, 115, 4011–4017. [Google Scholar] [CrossRef]
- Hirbawi, J.; Bialkowska, K.; Bledzka, K.M.; Liu, J.; Fukuda, K.; Qin, J.; Plow, E.F. The extreme C-terminal region of kindlin-2 is critical to its regulation of integrin activation. J. Biol. Chem. 2017, 292, 14258–14269. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Ma, Y.-Q.; Tu, Y.; Chen, K.; Wu, S.; Fukuda, K.; Qin, J.; Plow, E.F.; Wu, C. The MIG-2/Integrin Interaction Strengthens Cell-Matrix Adhesion and Modulates Cell Motility. J. Biol. Chem. 2007, 282, 20455–20466. [Google Scholar] [CrossRef] [Green Version]
- Rensen, S.S.M.; Doevendans, P.A.F.M.; Van Eys, G.J.J.M. Regulation and characteristics of vascular smooth muscle cell phenotypic diversity. Neth. Hearth J. 2007, 15, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Qu, H.; Tu, Y.; Shi, X.; Larjava, H.; Saleem, M.A.; Shattil, S.J.; Fukuda, K.; Qin, J.; Kretzler, M.; Wu, C. Kindlin-2 regulates podocyte adhesion and fibronectin matrix deposition through interactions with phosphoinositides and integrins. J. Cell Sci. 2011, 124, 879–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perera, D.; Ma, Y.-Q.; Yang, J.; Hirbawi, J.; Plow, E.F.; Qin, J. Membrane Binding of the N-Terminal Ubiquitin-Like Domain of Kindlin-2 Is Crucial for Its Regulation of Integrin Activation. Structure 2011, 19, 1664–1671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Fukuda, K.; Xu, Z.; Ma, Y.-Q.; Hirbawi, J.; Mao, X.; Wu, C.; Plow, E.F.; Qin, J. Structural Basis of Phosphoinositide Binding to Kindlin-2 Protein Pleckstrin Homology Domain in Regulating Integrin Activation. J. Biol. Chem. 2011, 286, 43334–43342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhu, Y.; Ye, S.; Zhang, R. Crystal structure of kindlin-2 PH domain reveals a conformational transition for its membrane anchoring and regulation of integrin activation. Protein Cell 2012, 3, 434–440. [Google Scholar] [CrossRef] [Green Version]
- Metcalf, D.G.; Moore, D.T.; Wu, Y.; Kielec, J.M.; Molnar, K.; Valentine, K.G.; Wand, A.J.; Bennett, J.S.; DeGrado, W.F. NMR analysis of the alphaIIb beta3 cytoplasmic interaction suggests a mechanism for integrin regulation. Proc. Natl. Acad. Sci. USA 2010, 107, 22481–22486. [Google Scholar] [CrossRef] [Green Version]
- Hart, R.; Stanley, P.; Chakravarty, P.; Hogg, N. The Kindlin 3 Pleckstrin Homology Domain Has an Essential Role in Lymphocyte Function-associated Antigen 1 (LFA-1) Integrin-mediated B Cell Adhesion and Migration. J. Biol. Chem. 2013, 288, 14852–14862. [Google Scholar] [CrossRef] [Green Version]
- Wen, L.; Marki, A.; Roy, P.; McArdle, S.; Sun, H.; Fan, Z.; Gingras, A.R.; Ginsberg, M.H.; Ley, K. Kindlin-3 recruitment to the plasma membrane precedes high affinity beta2 integrin and neutrophil arrest from rolling. Blood 2021, 137, 29–38. [Google Scholar] [CrossRef]
- Bledzka, K.; Liu, J.; Xu, Z.; Perera, H.D.; Yadav, S.P.; Bialkowska, K.; Qin, J.; Ma, Y.; Plow, E.F. Spatial coordination of kindlin-2 with talin head domain in interaction with integrin beta cytoplasmic tails. J. Biol. Chem. 2012, 287, 24585–24594. [Google Scholar] [CrossRef] [Green Version]
- Plow, E.F.; Das, M.; Bialkowska, K.; Sossey-Alaoui, K. Of Kindlins and Cancer. Discoveries 2016, 4, e59. [Google Scholar] [CrossRef] [Green Version]
- Plow, E.F.; Qin, J. The Kindlin Family of Adapter Proteins. Circ. Res. 2019, 124, 202–204. [Google Scholar] [CrossRef]
- Ussar, S.; Wang, H.-V.; Linder, S.; Fässler, R.; Moser, M. The Kindlins: Subcellular localization and expression during murine development. Exp. Cell Res. 2006, 312, 3142–3151. [Google Scholar] [CrossRef]
- Meves, A.; Stremmel, C.; Gottschalk, K.; Fässler, R. The Kindlin protein family: New members to the club of focal adhesion proteins. Trends Cell Biol. 2009, 19, 504–513. [Google Scholar] [CrossRef]
- Karaköse, E.; Schiller, H.B.; Fässler, R. The kindlins at a glance. J. Cell Sci. 2010, 123, 2353–2356. [Google Scholar] [CrossRef] [Green Version]
- Ussar, S.; Moser, M.; Widmaier, M.; Rognoni, E.; Harrer, C.; Genzel-Boroviczény, O.; Fässler, R. Loss of Kindlin-1 Causes Skin Atrophy and Lethal Neonatal Intestinal Epithelial Dysfunction. PLoS Genet. 2008, 4, e1000289. [Google Scholar] [CrossRef] [Green Version]
- Moser, M.; Nieswandt, B.; Ussar, S.; Pozgajova, M.; Fässler, R. Kindlin-3 is essential for integrin activation and platelet aggregation. Nat. Med. 2008, 14, 325–330. [Google Scholar] [CrossRef]
- Malinin, N.L.; Pluskota, E.; Byzova, T.V. Integrin signaling in vascular function. Curr. Opin. Hematol. 2012, 19, 206–211. [Google Scholar] [CrossRef] [Green Version]
- Montanez, E.; Ussar, S.; Schifferer, M.; Bösl, M.; Zent, R.; Moser, M.; Fässler, R. Kindlin-2 controls bidirectional signaling of integrins. Genes Dev. 2008, 22, 1325–1330. [Google Scholar] [CrossRef] [Green Version]
- Dowling, J.J.; Gibbs, E.; Russell, M.; Goldman, D.; Minarcik, J.; Golden, J.A.; Feldman, E.L. Kindlin-2 Is an Essential Component of Intercalated Discs and Is Required for Vertebrate Cardiac Structure and Function. Circ. Res. 2008, 102, 423–431. [Google Scholar] [CrossRef] [Green Version]
- Bialkowska, K.; Ma, Y.-Q.; Bledzka, K.; Sossey-Alaoui, K.; Izem, L.; Zhang, X.; Malinin, N.; Qin, J.; Byzova, T.; Plow, E.F. The Integrin Co-activator Kindlin-3 Is Expressed and Functional in a Non-hematopoietic Cell, the Endothelial Cell. J. Biol. Chem. 2010, 285, 18640–18649. [Google Scholar] [CrossRef] [Green Version]
- Harburger, D.S.; Bouaouina, M.; Calderwood, D.A. Kindlin-1 and -2 directly bind the C-terminal region of beta integrin cytoplasmic tails and exert integrin-specific activation effects. J. Biol. Chem 2009, 284, 11485–11497. [Google Scholar] [CrossRef] [Green Version]
- Sossey-Alaoui, K.; Pluskota, E.; Davuluri, G.; Bialkowska, K.; Das, M.; Szpak, D.; Lindner, D.J.; Downs-Kelly, E.; Thompson, C.L.; Plow, E.F. Kindlin-3 enhances breast cancer progression and metastasis by activating Twist-mediated angiogenesis. FASEB J. 2014, 28, 2260–2271. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Deng, Y.; Sun, K.; Yang, H.; Liu, J.; Wang, M.; Zhang, Z.; Lin, J.; Wu, C.; Wei, Z.; et al. Structural basis of kindlin-mediated integrin recognition and activation. Proc. Natl. Acad. Sci. USA 2017, 114, 9349–9354. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Xiao, D.; Ni, Y.; Zhang, T.; Cao, Z.; Xu, Z.; Nguyen, H.; Zhang, J.; White, G.C.; Ding, J.; et al. Structure basis of the FERM domain of kindlin-3 in supporting integrin alphaIIbbeta3 activation in platelets. Blood Adv. 2020, 4, 3128–3135. [Google Scholar] [CrossRef]
- Bu, W.; Levitskaya, Z.; Loh, Z.Y.; Jin, S.; Basu, S.; Ero, R.; Yan, X.; Wang, M.; Ngan, S.F.C.; Sze, S.K.; et al. Structural basis of human full-length kindlin-3 homotrimer in an auto-inhibited state. PLoS Biol. 2020, 18, e3000755. [Google Scholar] [CrossRef]
- Yates, L.A.; Lumb, C.N.; Brahme, N.N.; Zalyte, R.; Bird, L.E.; De Colibus, L.; Owens, R.J.; Calderwood, D.A.; Sansom, M.S.P.; Gilbert, R.J.C. Structural and Functional Characterization of the Kindlin-1 Pleckstrin Homology Domain. J. Biol. Chem. 2012, 287, 43246–43261. [Google Scholar] [CrossRef] [Green Version]
- Yates, L.A.; Füzéry, A.K.; Bonet, R.; Campbell, I.D.; Gilbert, R.J.C. Biophysical analysis of Kindlin-3 reveals an elongated conformation and maps integrin binding to the membrane-distal beta-subunit NPXY motif. J. Biol. Chem. 2012, 287, 37715–37731. [Google Scholar] [CrossRef] [Green Version]
- Kadry, Y.A.; Maisuria, E.M.; Huet-Calderwood, C.; Calderwood, D.A. Differences in self-association between kindlin-2 and kindlin-3 are associated with differential integrin binding. J. Biol. Chem. 2020, 295, 11161–11173. [Google Scholar] [CrossRef]
- Ye, F.; Hu, G.; Taylor, D.; Ratnikov, B.; Bobkov, A.A.; McLean, M.A.; Sligar, S.G.; Taylor, K.A.; Ginsberg, M.H. Recreation of the terminal events in physiological integrin activation. J. Cell Biol. 2010, 188, 157–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefort, C.T.; Rossaint, J.; Moser, M.; Petrich, B.G.; Zarbock, A.; Monkley, S.J.; Critchley, D.R.; Ginsberg, M.H.; Fässler, R.; Ley, K. Distinct roles for talin-1 and kindlin-3 in LFA-1 extension and affinity regulation. Blood 2012, 119, 4275–4282. [Google Scholar] [CrossRef] [PubMed]
- Ye, F.; Petrich, B.G.; Anekal, P.; Lefort, C.T.; Kasirer-Friede, A.; Shattil, S.J.; Moser, M.; Fässler, R.; Ginsberg, M.H. The mechanism of kindlin-mediated activation of integrin alphaIIbbeta3. Curr. Biol. 2013, 23, 2288–2295. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Gao, J.; Hong, J.; Ma, Y.-Q. Integrity of kindlin-2 FERM subdomains is required for supporting integrin activation. Biochem. Biophys. Res. Commun. 2013, 434, 382–387. [Google Scholar] [CrossRef] [PubMed]
- Theodosiou, M.; Widmaier, M.; Böttcher, R.T.; Rognoni, E.; Veelders, M.; Bharadwaj, M.; Lambacher, A.; Austen, K.; Müller, D.J.; Zent, R.; et al. Kindlin-2 cooperates with talin to activate integrins and induces cell spreading by directly binding paxillin. eLife 2016, 5, e10130. [Google Scholar] [CrossRef] [PubMed]
- Bachir, A.I.; Zareno, J.; Moissoglu, K.; Plow, E.F.; Gratton, E.; Horwitz, A.R. Integrin-Associated Complexes Form Hierarchically with Variable Stoichiometry in Nascent Adhesions. Curr. Biol. 2014, 24, 1845–1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasirer-Friede, A.; Moran, B.; Nagrampa-Orje, J.; Swanson, K.; Ruggeri, Z.M.; Schraven, B.; Neel, B.G.; Koretzky, G.; Shattil, S.J. ADAP is required for normal alphaIIbbeta3 activation by VWF/GP Ib-IX-V and other agonists. Blood 2007, 109, 1018–1025. [Google Scholar] [CrossRef]
- Nordenfelt, P.; Elliott, H.L.; Springer, T.A. Coordinated integrin activation by actin-dependent force during T-cell migration. Nat. Commun. 2016, 7, 13119. [Google Scholar] [CrossRef] [Green Version]
- Goult, B.T.; Yan, J.; Schwartz, M.A. Talin as a mechanosensitive signaling hub. J. Cell Biol. 2018, 217, 3776–3784. [Google Scholar] [CrossRef] [Green Version]
- Bialkowska, K.; Byzova, T.V.; Plow, E.F. Site-specific phosphorylation of kindlin-3 protein regulates its capacity to control cellular responses mediated by integrin alphaIIbbeta3. J. Biol. Chem. 2015, 290, 6226–6242. [Google Scholar] [CrossRef] [Green Version]
- Patel, H.; Zich, J.; Serrels, B.; Rickman, C.; Hardwick, K.G.; Frame, M.C.; Brunton, V.G. Kindlin-1 regulates mitotic spindle formation by interacting with integrins and Plk-1. Nat. Commun. 2013, 4, 2056. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.-F.; Tan, S.-M. The focal adhesion protein kindlin-2 controls mitotic spindle assembly by inhibiting histone deacetylase 6 and maintaining α-tubulin acetylation. J. Biol. Chem. 2020, 295, 5928–5943. [Google Scholar] [CrossRef] [Green Version]
- Qu, H.; Tu, Y.; Guan, J.-L.; Xiao, G.; Wu, C. Kindlin-2 Tyrosine Phosphorylation and Interaction with Src Serve as a Regulatable Switch in the Integrin Outside-in Signaling Circuit. J. Biol. Chem. 2014, 289, 31001–31013. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Lu, D.; Wang, X.; Wan, J.; Liu, C.; Zhang, H. Kindlin-2 phosphorylation by Src at Y193 enhances Src activity and is involved in Migfilin recruitment to the focal adhesions. FEBS Lett. 2015, 589, 2001–2010. [Google Scholar] [CrossRef] [Green Version]
- Artym, V.V.; Swatkoski, S.; Matsumoto, K.; Campbell, C.B.; Petrie, R.J.; Dimitriadis, E.K.; Li, X.; Mueller, S.C.; Bugge, T.H.; Gucek, M.; et al. Dense fibrillar collagen is a potent inducer of invadopodia via a specific signaling network. J. Cell Biol. 2015, 208, 331–350. [Google Scholar] [CrossRef]
- Liu, G.; Bao, Y.; Liu, C.; Zhu, Q.; Zhao, L.; Lu, X.; Zhu, Q.; Lv, Y.; Bai, F.; Wen, H. IKKepsilon phosphorylates kindlin-2 to induce invadopodia formation and promote colorectal cancer metastasis. Theranostics 2020, 10, 2358–2373. [Google Scholar] [CrossRef]
- Bialkowska, K.; Sossey-Alaoui, K.; Pluskota, E.; Izem, L.; Qin, J.; Plow, E.F. Site-specific phosphorylation regulates the functions of kindlin-3 in a variety of cells. Life Sci. Alliance 2019, 3, e201900594. [Google Scholar] [CrossRef]
- Margraf, A.; Germena, G.; Drexler, H.C.A.; Rossaint, J.; Ludwing, N.; Prystaj, B.; Mersamann, S.; Thomas, K.; Block, H.; Gottschilch, W.; et al. The integrin linked kinase is required for chemokine-triggered high affinity conformation of neutrophil beta2-integrin LFA1. Blood 2020, 136, 2200–2205. [Google Scholar] [CrossRef]
- Ruperez, P.; Gago-Martinez, A.; Burlingame, A.L.; Oses-Prieto, J.A. Quantitative Phosphoproteomic Analysis Reveals a Role for Serine and Threonine Kinases in the Cytoskeletal Reorganization in Early T Cell Receptor Activation in Human Primary T Cells. Mol. Cell. Proteom. 2012, 11, 171–186. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Wang, X.; Zhan, J.; Chen, Y.; Fang, W.; Zhang, L.; Zhang, H. Smurf1 inhibits integrin activation by controlling Kindlin-2 ubiquitination and degradation. J. Cell Biol. 2017, 216, 1455–1471. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Rajfur, Z.; Yousefi, N.; Chen, Z.; Jacobson, K.; Ginsberg, M.H. Talin phosphorylation by Cdk5 regulates Smurf1-mediated talin head ubiquitylation and cell migration. Nat. Cell Biol. 2009, 11, 624–630. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Malinin, N.L.; Meller, J.; Ma, Y.; West, X.Z.; Bledzka, K.; Qin, J.; Podrez, E.A.; Byzova, T.V. Regulation of Cell Adhesion and Migration by Kindlin-3 Cleavage by Calpain. J. Biol. Chem. 2012, 287, 40012–40020. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| Domain | Kindlin-1 | Kindlin-2 | Kindlin-3 |
|---|---|---|---|
| F0 | T8, S11,T30 | T32, T36, S59 | T6, S8, Y11, S14, S31 |
| F1 | S169, S170, S174, S179, Y191 | T129, S159, T172, S175, S177, Y179, S180, S181, Y185, S186, T192, Y193, S199, T205, S258 | Y162, S218 |
| F2 | S361, Y392, Y452 | S339, S351, Y378, Y395, S405, S414, T416, S435 | S337, S345, T361, S428, T482, S484, Y504 |
| F3 | S506, S520, T633 | S523, T536, S550, Y590, S610, S666 | T591, S595, S622, S625, S642 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bialkowska, K.; Qin, J.; Plow, E.F. Phosphorylation of Kindlins and the Control of Integrin Function. Cells 2021, 10, 825. https://doi.org/10.3390/cells10040825
Bialkowska K, Qin J, Plow EF. Phosphorylation of Kindlins and the Control of Integrin Function. Cells. 2021; 10(4):825. https://doi.org/10.3390/cells10040825
Chicago/Turabian StyleBialkowska, Katarzyna, Jun Qin, and Edward F. Plow. 2021. "Phosphorylation of Kindlins and the Control of Integrin Function" Cells 10, no. 4: 825. https://doi.org/10.3390/cells10040825