
Penile Cancer-Derived Cells Molecularly Characterized as Models to Guide Targeted Therapies

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Establishment of Penile Cancer-Derived Cells
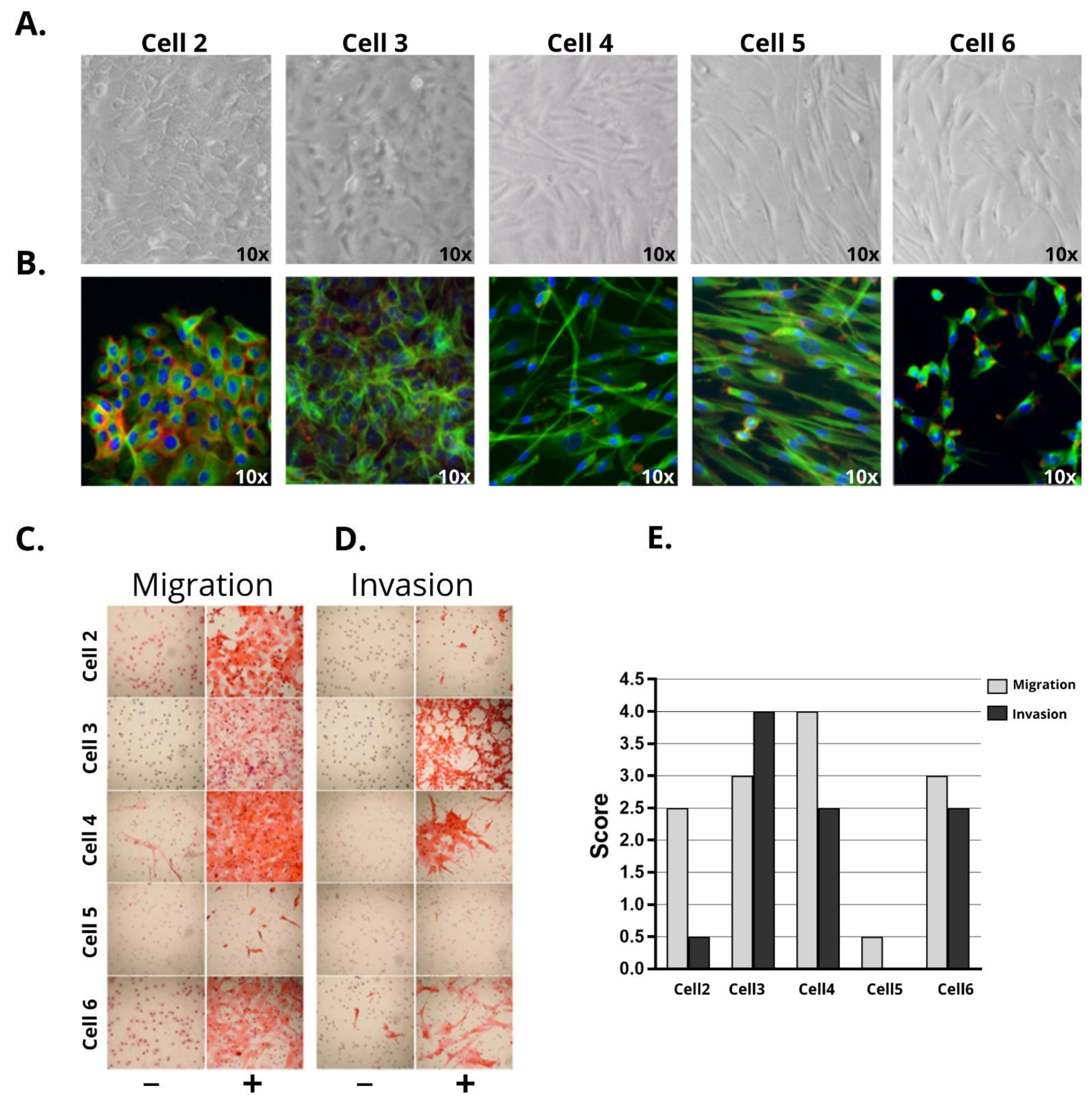
2.2. Morphological Characterization and Functional Assays
2.3. Molecular Profiling of Penile Cancer-Derived Cells
3. Results
3.1. Penile Cancer Derived-Cells Present Diverse Morphology, Proliferation, Migratory, and Invasive Characteristics
3.2. Penile Cancer-Derived Cells Recapitulate the Molecular Profile of PeCa Primary Tissues
3.3. Identification of Potential Therapeutic Targets in PeCa Cells Using Translatomic, Pathways, and Protein Analysis
3.4. Identification of Potential Therapeutic Targets for PeCa and Chemo-Sensitivity Assays
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Christodoulidou, M.; Sahdev, V.; Houssein, S.; Muneer, A. Epidemiology of penile cancer. Curr. Probl. Cancer 2015, 39, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Douglawi, A.; Masterson, T.A. Updates on the epidemiology and risk factors for penile cancer. Transl. Androl. Urol. 2017, 6, 785–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhoeven, R.; Janssen-Heijnen, M.; Saum, K.; Zanetti, R.; Caldarella, A.; Holleczek, B.; Brewster, D.; Hakulinen, T.; Horenblas, S.; Brenner, H.; et al. Population-based survival of penile cancer patients in Europe and the United States of America: No improvement since 1990. Eur. J. Cancer 2013, 49, 1414–1421. [Google Scholar] [CrossRef] [PubMed]
- Trama, A.; Foschi, R.; Larrañaga, N.; Sant, M.; Fuentes-Raspall, R.; Serraino, D.; Tavilla, A.; Van Eycken, L.; Nicolai, N.; Hackl, M.; et al. Survival of male genital cancers (prostate, testis and penis) in Europe 1999–2007: Results from the EUROCARE-5 study. Eur. J. Cancer 2015, 51, 2206–2216. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.; Necchi, A.; Muneer, A.; Tobias-Machado, M.; Tran, A.T.H.; Van Rompuy, A.-S.; Spiess, P.E.; Albersen, M. Penile cancer. Nat. Rev. Dis. Prim. 2021, 7, 1–24. [Google Scholar] [CrossRef]
- Kuasne, H.; Marchi, F.A.; Rogatto, S.R.; Cólus, I.M.D.S. Epigenetic Mechanisms in Penile Carcinoma. Int. J. Mol. Sci. 2013, 14, 10791–10808. [Google Scholar] [CrossRef] [Green Version]
- Thomas, A.; Vanthoor, J.; Vos, G.; Tsaur, I.; Albersen, M. Risk factors and molecular characterization of penile cancer. Curr. Opin. Urol. 2020, 30, 202–207. [Google Scholar] [CrossRef]
- Busso-Lopes, A.F.; Marchi, F.A.; Kuasne, H.; Scapulatempo-Neto, C.; Trindade-Filho, J.C.S.; De Jesus, C.M.N.; Lopes, A.; Guimarães, G.C.; Rogatto, S.R. Genomic Profiling of Human Penile Carcinoma Predicts Worse Prognosis and Survival. Cancer Prev. Res. 2015, 8, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Kuasne, H.; Barros-Filho, M.C.; Busso-Lopes, A.; Marchi, F.A.; Pinheiro, M.; Muñoz, J.J.M.; Scapulatempo-Neto, C.; Faria, E.F.; Guimarães, G.C.; Lopes, A.; et al. Integrative miRNA and mRNA analysis in penile carcinomas reveals markers and pathways with potential clinical impact. Oncotarget 2017, 8, 15294–15306. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, J.J.A.M.; Drigo, S.A.; Kuasne, H.; Villacis, R.A.R.; Marchi, F.A.; Domingues, M.A.C.; Lopes, A.; Santos, T.G.; Rogatto, S.R. A comprehensive characterization of cell cultures and xenografts derived from a human verrucous penile carcinoma. Tumor Biol. 2016, 37, 11375–11384. [Google Scholar] [CrossRef]
- Muñoz, J.J.; Drigo, S.A.; Barros-Filho, M.C.; Marchi, F.A.; Scapulatempo-Neto, C.; Pessoa, G.S.; Guimarães, G.C.; Filho, J.C.S.T.; Lopes, A.; Arruda, M.A.; et al. Down-Regulation of SLC8A1 as a Putative Apoptosis Evasion Mechanism by Modulation of Calcium Levels in Penile Carcinoma. J. Urol. 2015, 194, 245–251. [Google Scholar] [CrossRef]
- Medeiros-Fonseca, B.; Cubilla, A.; Brito, H.; Martins, T.; Medeiros, R.; Oliveira, P.; Gil da Costa, R. Experimental Models for Studying HPV-Positive and HPV-Negative Penile Cancer: New Tools for An Old Disease. Cancers 2021, 13, 460. [Google Scholar] [CrossRef] [PubMed]
- Yamane, I.; Tsuda, T. Establishment of a Cell Line in vitro from the Lesion of a Clinical Case of Penis Cancroid. Tohoku J. Exp. Med. 1966, 88, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikawa, S.; Kanoh, S.; Nemoto, S. Establishment of a cell line (TSUS-1) derived from a human squamous cell carcinoma of the penis. Hinyokika kiyo. Acta Urol. Jpn. 1983, 29, 373–376. [Google Scholar]
- Gentile, G.; Giraldo, G.; Stabile, M.; Beth-Giraldo, E.; Lonardo, F.; Kyalwazi, S.K.; Perone, L.; Ventruto, V. Cytogenetic study of a cell line of human penile cancer. Ann. de Génétique 1987, 30, 164–169. [Google Scholar]
- Tsukamoto, T. Establishment and characterization of a cell line (KU-8) from squamous cell carcinoma of the penis. Keio J. Med. 1989, 38, 277–293. [Google Scholar] [CrossRef] [PubMed]
- Naumann, C.M.; Sperveslage, J.; Hamann, M.F.; Leuschner, I.; Weder, L.; Al-Najar, A.A.; Lemke, J.; Sipos, B.; Jünemann, K.-P.; Kalthoff, H. Establishment and Characterization of Primary Cell Lines of Squamous Cell Carcinoma of the Penis and its Metastasis. J. Urol. 2012, 187, 2236–2242. [Google Scholar] [CrossRef]
- Chen, J.; Yao, K.; Li, Z.; Deng, C.; Wang, L.; Yu, X.; Liang, P.; Xie, Q.; Chen, P.; Qin, Z.; et al. Establishment and characterization of a penile cancer cell line, penl1, with a deleterious TP53 mutation as a paradigm of HPV-negative penile carcinogenesis. Oncotarget 2016, 7, 51687–51698. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.-H.; Deng, C.-Z.; Li, Z.-S.; Chen, J.-P.; Yao, K.; Huang, K.-B.; Liu, T.-Y.; Liu, Z.-W.; Qin, Z.-K.; Zhou, F.-J.; et al. Molecular characterization and integrative genomic analysis of a panel of newly established penile cancer cell lines. Cell Death Dis. 2018, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Fenner, F.; Goody, D.; Protzel, C.; Erbersdobler, A.; Richter, C.; Hartz, J.M.; Naumann, C.M.; Kalthoff, H.; Herchenröder, O.; Hakenberg, O.W.; et al. E2F1 Signalling is Predictive of Chemoresistance and Lymphogenic Metastasis in Penile Cancer: A Pilot Functional Study Reveals New Prognostic Biomarkers. Eur. Urol. Focus 2018, 4, 599–607. [Google Scholar] [CrossRef]
- Wang, L.; Saci, A.; Szabo, P.M.; Chasalow, S.D.; Castillo-Martin, M.; Domingo-Domenech, J.; Siefker-Radtke, A.; Sharma, P.; Sfakianos, J.P.; Gong, Y.; et al. EMT- and stroma-related gene expression and resistance to PD-1 blockade in urothelial cancer. Nat. Commun. 2018, 9, 3503. [Google Scholar] [CrossRef]
- Goulet, C.R.; Champagne, A.; Bernard, G.; Vandal, D.; Chabaud, S.; Pouliot, F.; Bolduc, S. Cancer-associated fibroblasts induce epithelial–mesenchymal transition of bladder cancer cells through paracrine IL-6 signalling. BMC Cancer 2019, 19, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Di Donato, M.; Zamagni, A.; Galasso, G.; Di Zazzo, E.; Giovannelli, P.; Barone, M.V.; Zanoni, M.; Gunelli, R.; Costantini, M.; Auricchio, F.; et al. The androgen receptor/filamin a complex as a target in prostate cancer microenvironment. Cell Death Dis. 2021, 12, 1–17. [Google Scholar] [CrossRef]
- Sahai, E.; Astsaturov, I.; Cukierman, E.; DeNardo, D.G.; Egeblad, M.; Evans, R.M.; Fearon, D.; Greten, F.R.; Hingorani, S.R.; Hunter, T.; et al. A framework for advancing our understanding of cancer-associated fibroblasts. Nat. Rev. Cancer 2020, 20, 174–186. [Google Scholar] [CrossRef] [Green Version]
- Villacis, R.A.R.; Basso, T.R.; Canto, L.M.; Nóbrega, A.F.; Achatz, M.I.; Rogatto, S.R. Germline large genomic alterations on 7q in patients with multiple primary cancers. Sci. Rep. 2017, 7, srep41677. [Google Scholar] [CrossRef]
- Canto, L.M.D.; Larsen, S.J.; Kupper, B.E.C.; Begnami, M.D.F.D.S.; Scapulatempo-Neto, C.; Petersen, A.H.; Aagaard, M.M.; Baumbach, J.; Aguiar, S.J.; Rogatto, S.R. Increased Levels of Genomic Instability and Mutations in Homologous Recombination Genes in Locally Advanced Rectal Carcinomas. Front. Oncol. 2019, 9, 395. [Google Scholar] [CrossRef] [PubMed]
- Mayrhofer, M.; Viklund, B.; Isaksson, A. Rawcopy: Improved copy number analysis with Affymetrix arrays. Sci. Rep. 2016, 6, 36158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karczewski, K.J.; Genome Aggregation Database Consortium; Francioli, L.C.; Tiao, G.; Cummings, B.B.; Alfoeldi, J.; Wang, Q.S.; Collins, R.L.; Laricchia, K.M.; Ganna, A.; et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature 2020, 581, 434–443. [Google Scholar] [CrossRef]
- Naslavsky, M.S.; Yamamoto, G.L.; De Almeida, T.F.; Ezquina, S.A.M.; Sunaga, D.Y.; Pho, N.; Bozoklian, D.; Sandberg, T.O.M.; Brito, L.A.; Lazar, M.; et al. Exomic variants of an elderly cohort of Brazilians in the ABraOM database. Hum. Mutat. 2017, 38, 751–763. [Google Scholar] [CrossRef] [PubMed]
- Roffé, M.; Hajj, G.N.; Azevedo, H.F.; Alves, V.S.; Castilho, B.A. IMPACT Is a Developmentally Regulated Protein in Neurons That Opposes the Eukaryotic Initiation Factor 2α Kinase GCN2 in the modulation of Neurite Outgrowth*. J. Biol. Chem. 2013, 288, 10860–10869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Han, C.; Wang, S.; Fang, P.; Ma, Z.; Xu, L.; Yin, R. Cancer-associated fibroblasts: An emerging target of anti-cancer immunotherapy. J. Hematol. Oncol. 2019, 12, 1–15. [Google Scholar] [CrossRef]
- Kalluri, R.; Zeisberg, M. Fibroblasts in cancer. Nat. Rev. Cancer 2006, 6, 392–401. [Google Scholar] [CrossRef]
- Nurmik, M.; Ullmann, P.; Rodriguez, F.; Haan, S.; Letellier, E. In search of definitions: Cancer-associated fibroblasts and their markers. Int. J. Cancer 2020, 146, 895–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichner, J.; Heubach, Y.; Ruff, M.; Kohlhof, H.; Strobl, S.; Mayer, B.; Pawlak, M.; Templin, M.F.; Zell, A. RPPApipe: A pipeline for the analysis of reverse-phase protein array data. Biosystems 2014, 122, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Kuasne, H.; Cólus, I.M.D.S.; Busso, A.F.; Hernandez-Vargas, H.; Barros-Filho, M.C.; Marchi, F.A.; Scapulatempo-Neto, C.; Faria, E.F.; Lopes, A.; Guimarães, G.C.; et al. Genome-wide methylation and transcriptome analysis in penile carcinoma: Uncovering new molecular markers. Clin. Epigenetics 2015, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stankiewicz, E.; Prowse, D.M.; Ng, M.; Cuzick, J.; Mesher, D.; Hiscock, F.; Lu, Y.-J.; Watkin, N.; Corbishley, C.; Lam, W.; et al. Alternative HER/PTEN/Akt Pathway Activation in HPV Positive and Negative Penile Carcinomas. PLoS ONE 2011, 6, e17517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaux, A.; Munari, E.; Cubilla, A.L.; Hicks, J.; Lecksell, K.; Burnett, A.L.; Netto, G.J. Immunohistochemical expression of the mammalian target of rapamycin pathway in penile squamous cell carcinomas: A tissue microarray study of 112 cases. Histopathology 2014, 64, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Azizi, M.; Tang, D.H.; Verduzco, D.; Peyton, C.C.; Chipollini, J.; Yuan, Z.; Schaible, B.J.; Zhou, J.-M.; Johnstone, P.A.; Giuliano, A.; et al. Impact of PI3K-AKT-mTOR Signaling Pathway Up-regulation on Prognosis of Penile Squamous-Cell Carcinoma: Results From a Tissue Microarray Study and Review of the Literature. Clin. Genitourin. Cancer 2019, 17, e80–e91. [Google Scholar] [CrossRef] [PubMed]
- Faraj, S.F.; Chaux, A.; Gonzalez-Roibon, N.; Munari, E.; Cubilla, A.L.; Shih, I.-M.; Netto, G.J. Immunohistochemical expression of ARID1A in penile squamous cell carcinomas: A tissue microarray study of 112 cases. Hum. Pathol. 2015, 46, 761–766. [Google Scholar] [CrossRef] [Green Version]
- Gunia, S.; Erbersdobler, A.; Hakenberg, O.W.; Koch, S.; May, M. C-MET is expressed in the majority of penile squamous cell carcinomas and correlates with polysomy-7 but is not associated with MET oncogene amplification, pertinent histopathologic parameters, or with cancer-specific survival. Pathol.-Res. Pract. 2013, 209, 215–220. [Google Scholar] [CrossRef]
- Arya, M.; Thrasivoulou, C.; Henrique, R.; Millar, M.; Hamblin, R.; Davda, R.; Aare, K.; Masters, J.R.; Thomson, C.; Muneer, A.; et al. Targets of Wnt/ß-Catenin Transcription in Penile Carcinoma. PLoS ONE 2015, 10, e0124395. [Google Scholar] [CrossRef]
- Amancio, A.M.T.D.S.; Da Cunha, I.W.; Neves, J.I.; Quetz, J.D.S.; Carraro, D.M.; Rocha, R.M.; Zequi, S.C.; Cubilla, A.L.; Da Fonseca, F.P.; Lopes, A.; et al. Epidermal growth factor receptor as an adverse survival predictor in squamous cell carcinoma of the penis. Hum. Pathol. 2017, 61, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Dorff, T.B.; Schuckman, A.K.; Schwartz, R.; Rashad, S.; Bulbul, A.; Cai, J.; Pinski, J.; Ma, Y.; Danenberg, K.; Skinner, E.; et al. Epidermal Growth Factor Receptor, Excision-Repair Cross-Complementation Group 1 Protein, and Thymidylate Synthase Expression in Penile Cancer. Clin. Genitourin. Cancer 2016, 14, 450–456.e1. [Google Scholar] [CrossRef] [PubMed]
- Ferrandiz-Pulido, C.; Masferrer, E.; Toll, A.; Hernandez-Losa, J.; Mojal, S.; Pujol, R.M.; Cajal, S.R.Y.; De Torres, I.; Garcia-Patos, V. mTOR Signaling Pathway in Penile Squamous Cell Carcinoma: pmTOR and peIF4E Over Expression Correlate with Aggressive Tumor Behavior. J. Urol. 2013, 190, 2288–2295. [Google Scholar] [CrossRef]
- Steinestel, J.; Al Ghazal, A.; Arndt, A.; Schnoeller, T.J.; Schrader, A.J.; Moeller, P.; Steinestel, K. The role of histologic subtype, p16(INK4a) expression, and presence of human papillomavirus DNA in penile squamous cell carcinoma. BMC Cancer 2015, 15, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cubilla, A.L.; Lloveras, B.; Alejo, M.; Clavero, O.; Chaux, A.; Kasamatsu, E.; Monfulleda, N.; Tous, S.; Alemany, L.; Klaustermeier, J.; et al. Value of p16INK4a in the Pathology of Invasive Penile Squamous Cell Carcinomas. Am. J. Surg. Pathol. 2011, 35, 253–261. [Google Scholar] [CrossRef]
- Tang, D.H.; Clark, P.E.; Giannico, G.; Hameed, O.; Chang, S.S.; Gellert, L.L. Lack of P16 ink4a Over Expression in Penile Squamous Cell Carcinoma is Associated with Recurrence after Lymph Node Dissection. J. Urol. 2015, 193, 519–525. [Google Scholar] [CrossRef]
- Mannweiler, S.; Sygulla, S.; Winter, E.; Regauer, S. Two major pathways of penile carcinogenesis: HPV-induced penile cancers overexpress p16ink4a, HPV-negative cancers associated with dermatoses express p53, but lack p16ink4a overexpression. J. Am. Acad. Dermatol. 2013, 69, 73–81. [Google Scholar] [CrossRef]
- Gunia, S.; Kakies, C.; Erbersdobler, A.; Hakenberg, O.W.; Koch, S.; May, M. Expression of p53, p21 and cyclin D1 in penile cancer: p53 predicts poor prognosis. J. Clin. Pathol. 2011, 65, 232–236. [Google Scholar] [CrossRef]
- Rocha, R.M.; Ignácio, J.A.; Jordán, J.; Carraro, D.M.; Lisboa, B.; Lopes, A.; Carvalho, K.C.; Da Cunha, I.W.; Cubilla, A.; Guimarães, G.C.; et al. A clinical, pathologic, and molecular study of p53 and murine double minute 2 in penile carcinogenesis and its relation to prognosis. Hum. Pathol. 2012, 43, 481–488. [Google Scholar] [CrossRef]
- Deng, C.; Li, Z.; Guo, S.; Chen, P.; Chen, X.; Zhou, Q.; Chen, J.; Yu, X.; Wu, X.; Ma, W.; et al. Tumor PD-L1 expression is correlated with increased TILs and poor prognosis in penile squamous cell carcinoma. OncoImmunology 2017, 6, e1269047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udager, A.M.; Liu, T.-Y.; Skala, S.L.; Magers, M.J.; McDaniel, A.S.; Spratt, D.E.; Feng, F.Y.; Siddiqui, J.; Cao, X.; Fields, K.L.; et al. Frequent PD-L1 expression in primary and metastatic penile squamous cell carcinoma: Potential opportunities for immunotherapeutic approaches. Ann. Oncol. 2016, 27, 1706–1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cocks, M.; Taheri, D.; Ball, M.W.; Bezerra, S.M.; Rodriguez, M.D.C.; Ricardo, B.F.; Bivalacqua, T.J.; Sharma, R.B.; Meeker, A.; Chaux, A.; et al. Immune-checkpoint status in penile squamous cell carcinoma: A North American cohort. Hum. Pathol. 2017, 59, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Ottenhof, S.R.; Djajadiningrat, R.S.; De Jong, J.; Thygesen, H.H.; Horenblas, S.; Jordanova, E.S. Expression of Programmed Death Ligand 1 in Penile Cancer is of Prognostic Value and Associated with HPV Status. J. Urol. 2017, 197, 690–697. [Google Scholar] [CrossRef] [PubMed]
- Termini, L.; Fregnani, J.H.; Boccardo, E.; Da Costa, W.H.; Longatto-Filho, A.; Andreoli, M.A.; Costa, M.C.; Lopes, A.; Da Cunha, I.W.; Soares, F.A.; et al. SOD2 immunoexpression predicts lymph node metastasis in penile cancer. BMC Clin. Pathol. 2015, 15, 3. [Google Scholar] [CrossRef] [Green Version]
- Lippert, T.H.; Ruoff, H.-J.; Volm, M. Current Status of Methods to Assess Cancer Drug Resistance. Int. J. Med Sci. 2011, 8, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Garnett, M.J.; McDermott, U. The evolving role of cancer cell line-based screens to define the impact of cancer genomes on drug response. Curr. Opin. Genet. Dev. 2014, 24, 114–119. [Google Scholar] [CrossRef] [Green Version]
- Cheung, P.F.-Y.; Yip, C.W.; Ng, L.W.-C.; Lo, K.W.; Wong, N.; Choy, K.W.; Chow, C.; Chan, K.F.; Cheung, T.T.; Poon, R.T.-P.; et al. Establishment and characterization of a novel primary hepatocellular carcinoma cell line with metastatic ability in vivo. Cancer Cell Int. 2014, 14, 103. [Google Scholar] [CrossRef] [Green Version]
- Ibarrola-Villava, M.; Cervantes, A.; Bardelli, A. Corrigendum to Preclinical models for precision oncology. BBACAN 1870/2 (2018) 239–246. Biochim. et Biophys. Acta (BBA)-Bioenerg. 2019, 1872, 188292. [Google Scholar] [CrossRef]
- Truffi, M.; Sorrentino, L.; Corsi, F. Fibroblasts in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2020, 1234, 15–29. [Google Scholar] [CrossRef]
- Miyai, Y.; Esaki, N.; Takahashi, M.; Enomoto, A. Cancer-associated fibroblasts that restrain cancer progression: Hypotheses and perspectives. Cancer Sci. 2020, 111, 1047–1057. [Google Scholar] [CrossRef] [Green Version]
- Burd, E.M. Human Papillomavirus and Cervical Cancer. Clin. Microbiol. Rev. 2003, 16, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guimarães, G.C.; Cunha, I.W.; Soares, F.A.; Lopes, A.; Torres, J.; Chaux, A.; Velazquez, E.F.; Ayala, G.; Cubilla, A.L. Penile Squamous Cell Carcinoma Clinicopathological Features, Nodal Metastasis and Outcome in 333 Cases. J. Urol. 2009, 182, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, D.F.; Soares, F.; Alvarado-Cabrero, I.; Cañete, S.; Fernández-Nestosa, M.J.; Rodríguez, I.M.; Barreto, J.; Cubilla, A.L. Pathological factors, behavior, and histological prognostic risk groups in subtypes of penile squamous cell carcinomas (SCC). Semin. Diagn. Pathol. 2015, 32, 222–231. [Google Scholar] [CrossRef]
- King, H.A.; Gerber, A.P. Translatome profiling: Methods for genome-scale analysis of mRNA translation. Briefings Funct. Genom. 2014, 15, 22–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaux, A.; Munari, E.; Katz, B.; Sharma, R.; Lecksell, K.; Cubilla, A.L.; Burnett, A.L.; Netto, G.J. The epidermal growth factor receptor is frequently overexpressed in penile squamous cell carcinomas: A tissue microarray and digital image analysis study of 112 cases. Hum. Pathol. 2013, 44, 2690–2695. [Google Scholar] [CrossRef]
- Di Lorenzo, G.; Buonerba, C.; Ferro, M.; Calderoni, G.; Bozza, G.; Federico, P.; Tedesco, B.; Ruggieri, V.; Aieta, M.; Federico, P. The epidermal growth factor receptors as biological targets in penile cancer. Expert Opin. Biol. Ther. 2014, 15, 473–476. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Sonpavde, G. Emerging Systemic Therapies for the Management of Penile Cancer. Urol. Clin. N. Am. 2016, 43, 481–491. [Google Scholar] [CrossRef]
- Huang, K.-B.; Liu, R.-Y.; Peng, Q.-H.; Li, Z.-S.; Jiang, L.-J.; Guo, S.-J.; Zhou, Q.-H.; Liu, T.-Y.; Deng, C.-Z.; Yao, K.; et al. EGFR mono-antibody salvage therapy for locally advanced and distant metastatic penile cancer: Clinical outcomes and genetic analysis. Urol. Oncol. Semin. Orig. Investig. 2019, 37, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Necchi, A.; Vullo, S.L.; Perrone, F.; Raggi, D.; Giannatempo, P.; Calareso, G.; Nicolai, N.; Piva, L.; Biasoni, D.; Catanzaro, M.; et al. First-line therapy with dacomitinib, an orally available pan-HER tyrosine kinase inhibitor, for locally advanced or metastatic penile squamous cell carcinoma: Results of an open-label, single-arm, single-centre, phase 2 study. BJU Int. 2017, 121, 348–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parseghian, C.M.; Napolitano, S.; Loree, J.M.; Kopetz, S. Mechanisms of Innate and Acquired Resistance to Anti-EGFR Therapy: A Review of Current Knowledge with a Focus on Rechallenge Therapies. Clin. Cancer Res. 2019, 25, 6899–6908. [Google Scholar] [CrossRef]
- Kuasne, H.; Barros-Filho, M.C.; Marchi, F.A.; Drigo, S.A.; Scapulatempo-Neto, C.; Faria, E.F.; Rogatto, S.R. Nuclear loss and cytoplasmic expression of androgen receptor in penile carcinomas: Role as a driver event and as a prognosis factor. Virchows Archiv für Pathol. Anat. und Physiol. und für Klin. Med. 2018, 473, 607–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, J.M.; Ferry, E.K.; Gay, L.M.; Elvin, J.A.; Vergilio, J.-A.; Ramkissoon, S.; Severson, E.; Necchi, A.; Killian, J.K.; Ali, S.M.; et al. Comparative Genomic Profiling of Refractory and Metastatic Penile and Nonpenile Cutaneous Squamous Cell Carcinoma: Implications for Selection of Systemic Therapy. J. Urol. 2019, 201, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Chahoud, J.; Pickering, C.R.; Pettaway, C.A. Genetics and penile cancer. Curr. Opin. Urol. 2019, 29, 364–370. [Google Scholar] [CrossRef] [PubMed]
- McGregor, B.; Sonpavde, G. Immunotherapy for advanced penile cancer—Rationale and potential. Nat. Rev. Urol. 2018, 15, 721–723. [Google Scholar] [CrossRef]
- Fruman, D.A.; Chiu, H.; Hopkins, B.D.; Bagrodia, S.; Cantley, L.C.; Abraham, R.T. The PI3K Pathway in Human Disease. Cell 2017, 170, 605–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T.-X.; Guan, X.-Y.; Fu, L. Therapeutic targeting of the crosstalk between cancer-associated fibroblasts and cancer stem cells. Am. J. Cancer Res. 2019, 9, 1889–1904. [Google Scholar]
- Skoda, A.M.; Simovic, D.; Karin, V.; Kardum, V.; Vranic, S.; Serman, L. The role of the Hedgehog signaling pathway in cancer: A comprehensive review. Bosn. J. Basic Med. Sci. 2018, 18, 8–20. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient/ Cell Line | Age (Years) | Histological Subtype | Surgery Type | HPV | TNM | Perineural Invasion | Clinical Stage | Follow-up (Months) |
|---|---|---|---|---|---|---|---|---|
| 2/Cell 2 | 85 | Usual + Sarcomatoid | Partial penectomy | HPV16 | T3N0M0 | Yes | III | 120 |
| 3/Cell 3 | 71 | Verrucous | Partial penectomy | Negative | T1N0M0 | No | I | 138 |
| 4/Cell 4 | 43 | Usual | Partial penectomy | Negative | T1N0M0 | Yes | I | 102 |
| 5/Cell 5 | 57 | Usual | Total penectomy | Negative | T4N0M1 | Yes | IV | 30 |
| 6/Cell 6 | 70 | Basaloid | Partial penectomy | Negative | T2N1M2 | No | IV | 18 |
| ID | Gene | Classification | Chr: Location | Type of Alteration | dbSNP | Transcript | Base Change |
|---|---|---|---|---|---|---|---|
| Cell 2/tumor 2 | PIK3CA | P | 3:17,8936,082 | Missense | rs121913273 | NM_006218.4 | c.1624G > A |
| Cell 2/tumor 2 | PIK3CA | P | 3:178,936,093 | Missense | rs121913275 | NM_006218.4 | c.1635G > C |
| Cell 2/tumor 2 | FGFR1 | VUS | 8:38,270,403 | Other | rs1364534792 | NM_023110.3 | c.*743dupA |
| Cell 2 | MMP1 | VUS | 11:102,667,445 | Missense | NM_002421.4 | c.575G > C | |
| Cell 2 | STAT3 | P | 17:40,486,045 | LoF | NM_139276.2 | c.820C > T | |
| Cell 3 | - | ||||||
| Cell 4 | RAD50 | VUS | 5:131,953,850 | Missense | rs143189763 | NM_005732.4 | c.3253A > G |
| Cell 4 | MMP1 | VUS | 11:102,668,717 | LoF | rs139018071 | NM_002421.4 | c.105+2T > C |
| Cell 4 | FLT3 | LP | 13:28,589,804 | Missense | rs903856095 | NM_004119.3 | c.2576G > A |
| Cell 5/tumor 5 | COL11A1 | LP | 1:103,380,339 | Missense | NM_001854.4 | c.3845G > T | |
| Cell 5/tumor 5 | ALK | VUS | 2:29,455,260 | Missense | NM_004304.5 | c.2542G > A | |
| Cell 5/tumor 5 | PIK3CA | P | 3:178,952,085 | Missense | rs121913279 | NM_006218.4 | c.3140A > G |
| Cell 5/tumor 5 | FBXW7 | LP | 4:153,247,367 | Missense | rs747241612 | NM_001349798.2 | c.1435C > G |
| Cell 5/tumor 5 | NOTCH1 | P | 9:139,413,064 | LoF | NM_017617.5 | c.1078G > T | |
| Cell 5/tumor 5 | BRCA2 | P | 13:32,968,951 | LoF | rs80359212 | NM_000059.3 | c.9382C > T |
| Cell 5/tumor 5 | IGF1R | VUS | 15:99507206 | Other | NM_000875.5 | c.*6535T > G | |
| Cell 5/tumor 5 | NF1 | VUS | 17:29,702,854 | Other | rs909909591 | NM_001042492.3 | c.*1683_*1685delGAA |
| Cell 5/tumor 5 | SMAD4 | LP | 18:48,575,116 | Missense | NM_005359.6 | c.310C > T | |
| Cell 6/tumor 6 | Near APC | VUS | 5:112,043,188 | upstream transcript variant | rs1554060178 | NC_000005.10 | 112707490:C:G |
| Tumor 6 | MLH3 | VUS | 14:75,483,802 | Missense | NM_001040108.1 | c.4345C > T | |
| Tumor 6 | ERBB2 | LP | 17:37,866,662 | Missense | NM_004448.3 | c.829G > T |
| Cell 2 (564) | Cell 3 (1199) | Cell 4 (262) | Cell 5 (205) | Cell 6 (163) | |||||
|---|---|---|---|---|---|---|---|---|---|
| Top 15 Overexpressed Genes | |||||||||
| Gene | FC | Gene | FC | Gene | FC | Gene | FC | Gene | FC |
| S100A8 | 54897.5 | OSR1 | 1865.57 | SFRP2 | 1936.2 | IL24 | 305.3 | TMEM176B | 2376.2 |
| COL17A1 | 35757.4 | FOXG1 | 1259.5 | TRPA1 | 964.7 | APOD | 101.4 | TMEM176A | 1686.2 |
| KRT5 | 25856.7 | PRKAR2B | 619.61 | LINC01436 | 458.9 | SFRP2 | 97.6 | APOD | 935.5 |
| S100A9 | 25339.7 | HOXD8 | 333.72 | TMEM176B | 288.4 | STMN2 | 83.6 | SLC14A1 | 873.7 |
| LCN2 | 23704.2 | LGALSL | 172.59 | TMEM176A | 252.5 | EGR1 | 79.7 | XGY2 | 446.0 |
| CDH1 | 18954.7 | PORCN | 119.5 | XGY2 | 247.8 | XG | 66.9 | XG | 374.6 |
| LAMC2 | 17269.9 | NRN1 | 86.36 | SAT1 | 113.8 | SNORD50A | 59.6 | CCND2 | 343.2 |
| MAL2 | 15941.8 | TBX15 | 74.35 | XG | 100.1 | ZBTB16 | 51.1 | FAM105A | 213.8 |
| GJB2 | 14969.1 | PRPF39 | 63.2 | IL33 | 98.9 | TNFRSF21 | 44.0 | SAA1 | 163.9 |
| FXYD3 | 13924.8 | MSI2 | 41.57 | APOD | 98.2 | DUXAP10 | 40.9 | STMN2 | 160.7 |
| KRT17 | 11118.8 | SLC44A1 | 40.69 | OSR1 | 88.5 | USP53 | 37.3 | AQP1 | 158.7 |
| DSG3 | 10259.7 | STX6 | 36.88 | PDGFRL | 72.3 | LINC01296 | 31.6 | IL13RA2 | 155.7 |
| FGFBP1 | 10054.2 | CUL3 | 34.26 | CCND2 | 65.9 | RGCC | 25.6 | DUSP6 | 147.2 |
| PI3 | 10052.7 | S100A7 | 32.87 | CLU | 62.3 | SFRP1 | 22.5 | FMO3 | 138.3 |
| S100A2 | 9269.6 | RTCB | 29.47 | FGF7 | 58.3 | EIF3A | 21.5 | ITGA8 | 138.2 |
| Top 15 Down-Expressed Genes | |||||||||
| Gene | FC | Gene | FC | Gene | FC | Gene | FC | Gene | FC |
| BGN | –15958.7 | CCL2 | –24130.3 | GFRA1 | –1175.8 | GFRA1 | –1156.6 | GFRA1 | –738.2 |
| GREM1 | –14798.9 | CLDN11 | –9744.1 | ADAM12 | –1155.9 | ADAM12 | –615.7 | CPA3 | –522.3 |
| UCHL1 | –13551.2 | UCHL1 | –9705.2 | IGFBP3 | –556.8 | POSTN | –585.0 | POSTN | –468.1 |
| MGST1 | –13191.8 | LUM | –9606.2 | PSG5 | –416.9 | CPA3 | –422.7 | CXCL12 | –169.9 |
| THY1 | –11417.6 | SULF1 | –7813.0 | CPA3 | –411.1 | UCP2 | –309.4 | PLPP4 | –147.7 |
| MXRA8 | –8181.8 | HIST1H2BM | –7623.5 | PLPP4 | –378.8 | UCHL1 | –255.8 | PSG2 | –119.4 |
| LOXL1 | –8004.0 | MXRA8 | –6464.9 | CNN1 | –365.3 | ACKR3 | –209.7 | PSG1 | –106.9 |
| ENG | –7918.5 | PSG5 | –6399.8 | CLDN11 | –275.9 | CNN1 | –189.1 | HIST1H3F | –96.9 |
| FBLN5 | –6160.9 | MGST1 | –6082.7 | PLPPR4 | –245.8 | COL1A1 | –133.3 | BEX1 | –95.9 |
| MFAP4 | –3745.0 | MRPL20 | –4740.9 | UCP2 | –243.7 | HIST1H3F | –114.1 | PSG8 | –66.6 |
| ADGRA2 | –3508.0 | FARP1 | –4690.7 | LBH | –220.6 | KRT7 | –79.1 | SHOX | –62.4 |
| COL1A2 | –2748.2 | THY1 | –4498.7 | PSG8 | –167.9 | MYBL2 | –73.4 | UCP2 | –55.3 |
| GFRA1 | –2705.4 | TGFBR1 | –4386.3 | SLC7A5 | –162.6 | LYPD6B | –63.9 | DAPK1 | –49.0 |
| F2RL2 | –2520.2 | HIST2H4B | –4137.9 | PSG11 | –160.0 | SEL1L3 | –53.0 | KCND3 | –45.9 |
| MYADM | –2340.8 | CSRP1 | –4129.1 | MEST | –138.6 | KCND3 | –44.6 | COLEC12 | –45.2 |
| Protein | Number of Cases | Expression | Reference | Cell | ||||
|---|---|---|---|---|---|---|---|---|
| 2 | 3 | 4 | 5 | 6 | ||||
| Akt1 | 148 | ↑ | Stankiewicz et al., 2011 [36] | ↓ | ↑ | - | ↑ | - |
| pAkt | 112 | ↑ | Chaux et al., 2014 [37] | - | ↑ | - | ↑ | ↓ |
| pAkt | 148 | ↑ | Stankiewicz et al., 2011 [36] | - | ↑ | - | ↑ | ↓ |
| pAkt | 57 | ↑ | Azizi et al., 2019 [38] | - | ↑ | - | ↑ | ↓ |
| ARID1A | 112 | ↑ | Faraj et al., 2015 [39] | ↑ | ↑ | ↓ | ↑ | ↑ |
| c-MET | 92 | ↑ | Gunia et al., 2013 [40] | - | - | - | - | - |
| c-MYC | 141 | ↓ | Arya et al., 2015 [41] | - | - | - | - | ↓ |
| cyclinD1 | 141 | ↓ | Arya et al., 2015 [41] | - | - | - | - | - |
| EGFR | 139 | ↑ | Silva Amancio et al., 2017 [42] | ↑ | - | - | ↑ | - |
| EGFR | 52 | ↑ | Dorff et al., 2016 [43] | ↑ | - | - | ↑ | - |
| EI2F | 13 | ↑ | Fenner et al., 2018 [20] | ↑ | ↑ | - | ↑ | - |
| eIF4E | 67 | ↑ | Ferrandiz-Pulido et al., 2013 [44] | ↑ | ↑ | - | - | ↓ |
| peIF4E | 67 | ↑ | Ferrandiz-Pulido et al., 2013 [44] | - | - | - | - | - |
| HER2 | 148 | ↑ | Stankiewicz et al., 2011 [36] | ↑ | ↑ | - | - | - |
| HER3 | 148 | ↑ | Stankiewicz et al., 2011 [36] | ↑ | ↑ | - | - | - |
| HER4 | 148 | ↑ | Stankiewicz et al., 2011 [36] | - | - | - | - | - |
| p16 | 58 | ↑ | Steinestel et al., 2015 [45] | ↓ | ↓ | - | ↓ | ↓ |
| p16 | 202 | ↑ | Cubilla et al., 2011 [46] | ↓ | ↓ | - | ↓ | ↓ |
| p16 | 119 | ↑ | Tang et al., 2015 [47] | ↓ | ↓ | - | ↓ | ↓ |
| p16 | 123 | ↑ | Mannweiler et al., 2013 [48] | ↓ | ↓ | - | ↓ | ↓ |
| p4E-BP1 | 67 | ↑ | Ferrandiz-Pulido et al., 2013 [44] | ↑ | ↑ | - | - | - |
| p53 | 123 | ↑ | Mannweiler et al., 2013 [48] | - | - | - | - | - |
| p53 | 110 | ↑ | Gunia et al., 2013 [49] | - | - | - | - | - |
| p53 | 297 | ↑ | Rocha et al., 2012 [50] | - | - | - | - | - |
| PDL1 | 116 | ↑ | Deng et al., 2017 [51] | - | ↑ | - | - | - |
| PDL1 | 37 | ↑ | Udager et al., 2016 [52] | - | ↑ | - | - | - |
| PDL1 | 52 | ↑ | Cocks et al., 2017 [53] | - | ↑ | - | - | - |
| PDL1 | 213 | ↑ | Ottenhof et al., 2017 [54] | - | ↑ | - | - | - |
| pmTOR | 67 | ↑ | Ferrandiz-Pulido et al., 2013 [44] | ↑ | - | - | - | ↑ |
| pmTOR | 112 | ↑ | Chaux et al., 2014 [37] | ↑ | - | - | - | ↑ |
| PTEN | 112 | ↓ | Chaux et al., 2014 [37] | - | - | - | - | - |
| PTEN | 148 | ↓ | Stankiewicz et al., 2011 [36] | ↓ | ↑ | - | ↓ | ↓ |
| PTEN | 57 | ↓ | Azizi et al., 2019 [38] | ↓ | ↑ | - | ↓ | ↓ |
| pS6 | 57 | ↑ | Azizi et al., 2019 [38] | ↑ | - | - | ↑ | - |
| SOD2 | 125 | ↑ | Termini et al., 2015 [55] | ↑ | ↑ | ↑ | ↑ | - |
| Cell Line | Gene Symbol a | mRNA FC | Location | Drug(s) b |
|---|---|---|---|---|
| mRNA | ||||
| Cell 2 | FRK | 144.007 | Nucleus | Regorafenib |
| Cell 2 | ERBB3 c | 121.938 | Plasma membrane | Afatinib/cetuximab/osimertinib/erlotinib |
| Cell 2 | MST1R | 54.569 | Plasma membrane | Crizotinib/erlotinib/gefitinib/pazopanib |
| Cell 2 | PRKDC | 43.713 | Nucleus | CC-115/MSC2490484A/panulisib |
| Cell 2 | EGFRc | 38.586 | Plasma membrane | Gefitinib/erlotinib/afatinib/cetuximab |
| Cell 2 | MET | 30.91 | Plasma membrane | Crizotinib/cabozantinib/ABT-700/altiratinib |
| Cell 3 | SRC | 28.443 | Cytoplasm | Dasatinib/bosutinib/blinatumomab/AZD0424 |
| Cell 2 | DDR1 | 24.59 | Plasma membrane | Blinatumomab/dacomitinib/dasatinib/imatinib |
| RPPA | ||||
| Cell 2 | KDR | 4.729 | Plasma membrane | 5-azacytidine/sorafenib/sAEE788/apatinib |
| Cell 2 | RPS6KA1 | 3.775 | Cytoplasm | PMD-026 |
| Cell 2 | MAP2K1 | 3.493 | Cytoplasm | ARRY-424704/AS703988/binimetinib |
| Cell 2 | MTOR | 2.925 | Nucleus | ABI-009/apitolisib/sirolimus/tacrolimus/everolimus |
| Cell 2 | SRC | 2.792 | Cytoplasm | AZD0424/dasatinib/bosutinib/ponatinib |
| Cell 3 | CDK1 | 2.637 | Nucleus | Alvocidib/dinaciclib/milciclib/riviciclib |
| Cell 2 | MKNK1 | 2.62 | Cytoplasm | BAY1143269/dacomitinib/ETC-1907206/tomivosertib |
| Cell 3 | AKT1 | 2.386 | Cytoplasm | BAY1125976/capivasertib/patasertib/miransertib |
| Cell 2 | BRD4 | 2.241 | Nucleus | AZD5153/BI 894999/PLX2853/PLX51107 |
| Cell 2 | CDK1 | 2.051 | Nucleus | Alvocidib/dinaciclib/milciclib/riviciclib |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuasne, H.; Canto, L.M.d.; Aagaard, M.M.; Muñoz, J.J.M.; Jamblinne, C.D.; Marchi, F.A.; Scapulatempo-Neto, C.; Faria, E.F.; Lopes, A.; Carréno, S.; et al. Penile Cancer-Derived Cells Molecularly Characterized as Models to Guide Targeted Therapies. Cells 2021, 10, 814. https://doi.org/10.3390/cells10040814
Kuasne H, Canto LMd, Aagaard MM, Muñoz JJM, Jamblinne CD, Marchi FA, Scapulatempo-Neto C, Faria EF, Lopes A, Carréno S, et al. Penile Cancer-Derived Cells Molecularly Characterized as Models to Guide Targeted Therapies. Cells. 2021; 10(4):814. https://doi.org/10.3390/cells10040814
Chicago/Turabian StyleKuasne, Hellen, Luisa Matos do Canto, Mads Malik Aagaard, Juan Jose Moyano Muñoz, Camille De Jamblinne, Fabio Albuquerque Marchi, Cristovam Scapulatempo-Neto, Eliney Ferreira Faria, Ademar Lopes, Sébastien Carréno, and et al. 2021. "Penile Cancer-Derived Cells Molecularly Characterized as Models to Guide Targeted Therapies" Cells 10, no. 4: 814. https://doi.org/10.3390/cells10040814