
Targeting the Hypoxic and Acidic Tumor Microenvironment with pH-Sensitive Peptides


Abstract
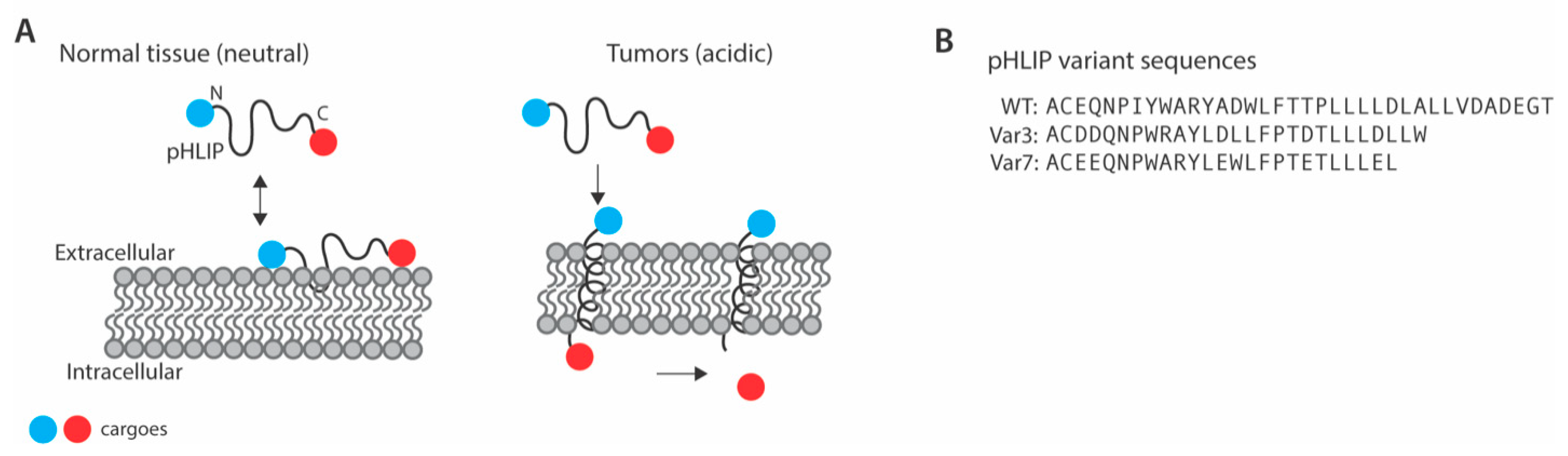
:1. Introduction
2. Targeting of the Tumor Microenvironment with pHLIP
3. Biodistribution of pHLIP
4. pHLIP Variants
5. pHLIP as a Small-Molecule Transporter of Toxins and Drugs to Tumor Cells
6. pHLIP as a Transporter of Nanoparticle-Based Encapsulated Therapeutics to Cancer Cells
7. pHLIP Mediated Delivery of Genetic Material (Peptide Nucleic Acids)
8. pHLIP-Based Approaches to Targeting DNA Repair and DNA Damage
9. pHLIP as a Tool for Diagnosing and Imaging Tumor Microenvironments
10. Non-Tumor Applications of pHLIP Technology
11. Limitations of pHLIP Technology
12. Conclusions and Perspectives
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vaupel, P.; Kallinowski, F.; Okunieff, P. Blood flow, oxygen and nutrient supply, and metabolic microenvironment of human tumors: A review. Cancer Res. 1989, 49, 6449–6465. [Google Scholar] [PubMed]
- Algayer, B.; O’Brien, A.; Momose, A.; Murphy, D.J.; Procopio, W.; Tellers, D.M.; Tucker, T.J. Novel pH Selective, Highly Lytic Peptides Based on a Chimeric Influenza Hemagglutinin Peptide/Cell Penetrating Peptide Motif. Molecules 2019, 24, 2079. [Google Scholar] [CrossRef] [Green Version]
- Svoronos, A.A.; Bahal, R.; Pereira, M.C.; Barrera, F.N.; Deacon, J.C.; Bosenberg, M.; DiMaio, D.; Glazer, P.M.; Engelman, D.M. Tumor-Targeted, Cytoplasmic Delivery of Large, Polar Molecules Using a pH-Low Insertion Peptide. Mol. Pharm. 2020, 17, 461–471. [Google Scholar] [CrossRef]
- Reshetnyak, Y.K.; Andreev, O.A.; Lehnert, U.; Engelman, D.M. Translocation of molecules into cells by pH-dependent insertion of a transmembrane helix. Proc. Natl. Acad. Sci. USA 2006, 103, 6460–6465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, J.F.; Rath, P.; Rothschild, K.J.; Engelman, D.M. Spontaneous, pH-dependent membrane insertion of a transbilayer alpha-helix. Biochemistry 1997, 36, 15177–15192. [Google Scholar] [CrossRef] [PubMed]
- Reshetnyak, Y.K.; Yao, L.; Zheng, S.; Kuznetsov, S.; Engelman, D.M.; Andreev, O.A. Measuring tumor aggressiveness and targeting metastatic lesions with fluorescent pHLIP. Mol. Imaging Biol. 2011, 13, 1146–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreev, O.A.; Dupuy, A.D.; Segala, M.; Sandugu, S.; Serra, D.A.; Chichester, C.O.; Engelman, D.M.; Reshetnyak, Y.K. Mechanism and uses of a membrane peptide that targets tumors and other acidic tissues in vivo. Proc. Natl. Acad. Sci. USA 2007, 104, 7893–7898. [Google Scholar] [CrossRef] [Green Version]
- Webb, B.A.; Chimenti, M.; Jacobson, M.P.; Barber, D.L. Dysregulated pH: A perfect storm for cancer progression. Nat. Rev. Cancer 2011, 11, 671–677. [Google Scholar] [CrossRef]
- Demoin, D.W.; Wyatt, L.C.; Edwards, K.J.; Abdel-Atti, D.; Sarparanta, M.; Pourat, J.; Longo, V.A.; Carlin, S.D.; Engelman, D.M.; Andreev, O.A.; et al. PET Imaging of Extracellular pH in Tumors with (64)Cu- and (18)F-Labeled pHLIP Peptides: A Structure-Activity Optimization Study. Bioconjug. Chem. 2016, 27, 2014–2023. [Google Scholar] [CrossRef]
- Dubois, L.G.; Campanati, L.; Righy, C.; D’Andrea-Meira, I.; Spohr, T.C.; Porto-Carreiro, I.; Pereira, C.M.; Balca-Silva, J.; Kahn, S.A.; DosSantos, M.F.; et al. Gliomas and the vascular fragility of the blood brain barrier. Front. Cell. Neurosci. 2014, 8, 418. [Google Scholar] [CrossRef] [Green Version]
- Burns, K.E.; Hensley, H.; Robinson, M.K.; Thevenin, D. Therapeutic Efficacy of a Family of pHLIP-MMAF Conjugates in Cancer Cells and Mouse Models. Mol. Pharm. 2017, 14, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Weerakkody, D.; Moshnikova, A.; Thakur, M.S.; Moshnikova, V.; Daniels, J.; Engelman, D.M.; Andreev, O.A.; Reshetnyak, Y.K. Family of pH (low) insertion peptides for tumor targeting. Proc. Natl. Acad. Sci. USA 2013, 110, 5834–5839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onyango, J.O.; Chung, M.S.; Eng, C.H.; Klees, L.M.; Langenbacher, R.; Yao, L.; An, M. Noncanonical amino acids to improve the pH response of pHLIP insertion at tumor acidity. Angew. Chem. Int. Ed. Engl. 2015, 54, 3658–3663. [Google Scholar] [CrossRef]
- Wyatt, L.C.; Moshnikova, A.; Crawford, T.; Engelman, D.M.; Andreev, O.A.; Reshetnyak, Y.K. Peptides of pHLIP family for targeted intracellular and extracellular delivery of cargo molecules to tumors. Proc. Natl. Acad. Sci. USA 2018, 115, E2811–E2818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, V.P.; Alves, D.S.; Scott, H.L.; Davis, F.L.; Barrera, F.N. A Novel Soluble Peptide with pH-Responsive Membrane Insertion. Biochemistry 2015, 54, 6567–6575. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A.; Franco Lombardo, B.W.; Dominy, P.J.F. Experimental and computational approaches to estimate solubility and permeability in drug discovery and developmentqsettings. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]
- Andreev, O.A.; Engelman, D.M.; Reshetnyak, Y.K. Targeting diseased tissues by pHLIP insertion at low cell surface pH. Front. Physiol. 2014, 5, 97. [Google Scholar] [CrossRef] [Green Version]
- Wijesinghe, D.; Engelman, D.M.; Andreev, O.A.; Reshetnyak, Y.K. Tuning a polar molecule for selective cytoplasmic delivery by a pH (Low) insertion peptide. Biochemistry 2011, 50, 10215–10222. [Google Scholar] [CrossRef] [Green Version]
- An, M.; Wijesinghe, D.; Andreev, O.A.; Reshetnyak, Y.K.; Engelman, D.M. pH-(low)-insertion-peptide (pHLIP) translocation of membrane impermeable phalloidin toxin inhibits cancer cell proliferation. Proc. Natl. Acad. Sci. USA 2010, 107, 20246–20250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moshnikova, A.; Moshnikova, V.; Andreev, O.A.; Reshetnyak, Y.K. Antiproliferative effect of pHLIP-amanitin. Biochemistry 2013, 52, 1171–1178. [Google Scholar] [CrossRef] [Green Version]
- Song, Q.; Chuan, X.; Chen, B.; He, B.; Zhang, H.; Dai, W.; Wang, X.; Zhang, Q. A smart tumor targeting peptide-drug conjugate, pHLIP-SS-DOX: Synthesis and cellular uptake on MCF-7 and MCF-7/Adr cells. Drug. Deliv. 2016, 23, 1734–1746. [Google Scholar] [CrossRef] [Green Version]
- Burns, K.E.; Robinson, M.K.; Thevenin, D. Inhibition of cancer cell proliferation and breast tumor targeting of pHLIP-monomethyl auristatin E conjugates. Mol. Pharm. 2015, 12, 1250–1258. [Google Scholar] [CrossRef]
- Adochite, R.C.; Moshnikova, A.; Carlin, S.D.; Guerrieri, R.A.; Andreev, O.A.; Lewis, J.S.; Reshetnyak, Y.K. Targeting breast tumors with pH (low) insertion peptides. Mol. Pharm. 2014, 11, 2896–2905. [Google Scholar] [CrossRef]
- Crawford, T.; Moshnikova, A.; Roles, S.; Weerakkody, D.; DuPont, M.; Carter, L.M.; Shen, J.; Engelman, D.M.; Lewis, J.S.; Andreev, O.A.; et al. pHLIP ICG for delineation of tumors and blood flow during fluorescence-guided surgery. Sci. Rep. 2020, 10, 18356. [Google Scholar] [CrossRef] [PubMed]
- Macholl, S.; Morrison, M.S.; Iveson, P.; Arbo, B.E.; Andreev, O.A.; Reshetnyak, Y.K.; Engelman, D.M.; Johannesen, E. In vivo pH imaging with (99m)Tc-pHLIP. Mol. Imaging Biol. 2012, 14, 725–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.; Meng, H.; Wang, N.; Donovan, M.J.; Fu, T.; You, M.; Chen, Z.; Zhang, X.; Tan, W. A controlled-release nanocarrier with extracellular pH value driven tumor targeting and translocation for drug delivery. Angew. Chem. Int. Ed. Engl. 2013, 52, 7487–7491. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Daniels, J.; Wijesinghe, D.; Andreev, O.A.; Reshetnyak, Y.K. pHLIP(R)-mediated delivery of PEGylated liposomes to cancer cells. J. Control. Release 2013, 167, 228–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijesinghe, D.; Arachchige, M.C.; Lu, A.; Reshetnyak, Y.K.; Andreev, O.A. pH dependent transfer of nano-pores into membrane of cancer cells to induce apoptosis. Sci. Rep. 2013, 3, 3560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, L.; Ma, H.; Guo, Y.; Kuang, Y.; He, X.; Jiang, C. pH-controlled delivery of nanoparticles into tumor cells. Adv. Healthc. Mater. 2013, 2, 1435–1439. [Google Scholar] [CrossRef]
- Yao, L.; Daniels, J.; Moshnikova, A.; Kuznetsov, S.; Ahmed, A.; Engelman, D.M.; Reshetnyak, Y.K.; Andreev, O.A. pHLIP peptide targets nanogold particles to tumors. Proc. Natl. Acad. Sci. USA 2013, 110, 465–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antosh, M.P.; Wijesinghe, D.D.; Shrestha, S.; Lanou, R.; Huang, Y.H.; Hasselbacher, T.; Fox, D.; Neretti, N.; Sun, S.; Katenka, N.; et al. Enhancement of radiation effect on cancer cells by gold-pHLIP. Proc. Natl. Acad. Sci. USA 2015, 112, 5372–5376. [Google Scholar] [CrossRef] [Green Version]
- Daniels, J.L.; Crawford, T.M.; Andreev, O.A.; Reshetnyak, Y.K. Synthesis and characterization of pHLIP((R)) coated gold nanoparticles. Biochem. Biophys. Rep. 2017, 10, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Deacon, J.; Yan, H.; Sun, B.; Liu, Y.; Hegan, D.; Li, Q.; Coman, D.; Parent, M.; Hyder, F.; et al. Tumor-targeted pH-low insertion peptide delivery of theranostic gadolinium nanoparticles for image-guided nanoparticle-enhanced radiation therapy. Transl. Oncol. 2020, 13, 100839. [Google Scholar] [CrossRef]
- Cheng, C.J.; Bahal, R.; Babar, I.A.; Pincus, Z.; Barrera, F.; Liu, C.; Svoronos, A.; Braddock, D.T.; Glazer, P.M.; Engelman, D.M.; et al. MicroRNA silencing for cancer therapy targeted to the tumour microenvironment. Nature 2015, 518, 107–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahraei, M.; Chaube, B.; Liu, Y.; Sun, J.; Kaplan, A.; Price, N.L.; Ding, W.; Oyaghire, S.; Garcia-Milian, R.; Mehta, S.; et al. Suppressing miR-21 activity in tumor-associated macrophages promotes an antitumor immune response. J. Clin. Investig. 2019, 129, 5518–5536. [Google Scholar] [CrossRef] [Green Version]
- Ozes, A.R.; Wang, Y.; Zong, X.; Fang, F.; Pilrose, J.; Nephew, K.P. Therapeutic targeting using tumor specific peptides inhibits long non-coding RNA HOTAIR activity in ovarian and breast cancer. Sci. Rep. 2017, 7, 894. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, A.R.; Pham, H.; Liu, Y.; Oyaghire, S.; Bahal, R.; Engelman, D.M.; Glazer, P.M. Ku80-Targeted pH-Sensitive Peptide-PNA Conjugates Are Tumor Selective and Sensitize Cancer Cells to Ionizing Radiation. Mol. Cancer Res. 2020, 18, 873–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quijano, E.; Bahal, R.; Ricciardi, A.; Saltzman, W.M.; Glazer, P.M. Therapeutic Peptide Nucleic Acids: Principles, Limitations, and Opportunities. Yale J. Biol. Med. 2017, 90, 583–598. [Google Scholar]
- Wagner, E. Tumor-targeted Delivery of Anti-microRNA for Cancer Therapy: pHLIP is Key. Angew. Chem. Int. Ed. Engl. 2015, 54, 5824–5826. [Google Scholar] [CrossRef]
- Ozes, A.R.; Miller, D.F.; Ozes, O.N.; Fang, F.; Liu, Y.; Matei, D.; Huang, T.; Nephew, K.P. NF-kappaB-HOTAIR axis links DNA damage response, chemoresistance and cellular senescence in ovarian cancer. Oncogene 2016, 35, 5350–5361. [Google Scholar] [CrossRef] [Green Version]
- Pilie, P.G.; Tang, C.; Mills, G.B.; Yap, T.A. State-of-the-art strategies for targeting the DNA damage response in cancer. Nat. Rev. Clin. Oncol. 2019, 16, 81–104. [Google Scholar] [CrossRef] [PubMed]
- De Ruysscher, D.; Niedermann, G.; Burnet, N.G.; Siva, S.; Lee, A.W.M.; Hegi-Johnson, F. Radiotherapy toxicity. Nat. Rev. Dis. Primers 2019, 5, 13. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.X.; Wu, H.L.; Shi, H.Y.; Su, L.; Zhang, X. The efficacy and safety of olaparib in the treatment of cancers: A meta-analysis of randomized controlled trials. Cancer Manag. Res. 2018, 10, 2553–2562. [Google Scholar] [CrossRef] [Green Version]
- Nussenzweig, A.; Sokol, K.; Burgman, P.; Li, L.; Li, G.C. Hypersensitivity of Ku80-deficient cell lines and mice to DNA damage: The effects of ionizing radiation on growth, survival, and development. Proc. Natl. Acad. Sci. USA 1997, 94, 13588–13593. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, H.; Nussenzweig, A.; Kurimasa, A.; Soares, V.C.; Li, X.; Cordon-Cardo, C.; Li, W.; Cheong, N.; Nussenzweig, M.; Iliakis, G.; et al. Ku70 is required for DNA repair but not for T cell antigen receptor gene recombination In vivo. J. Exp. Med. 1997, 186, 921–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pospisilova, M.; Seifrtova, M.; Rezacova, M. Small molecule inhibitors of DNA-PK for tumor sensitization to anticancer therapy. J. Physiol. Pharmacol. 2017, 68, 337–344. [Google Scholar]
- Davidson, D.; Amrein, L.; Panasci, L.; Aloyz, R. Small Molecules, Inhibitors of DNA-PK, Targeting DNA Repair, and Beyond. Front. Pharmacol. 2013, 4, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, A.K.; Cerniglia, G.J.; Mick, R.; Ahmed, M.S.; Bakanauskas, V.J.; Muschel, R.J.; McKenna, W.G. Radiation sensitization of human cancer cells in vivo by inhibiting the activity of PI3K using LY294002. Int. J. Radiat. Oncol. Biol. Phys. 2003, 56, 846–853. [Google Scholar] [CrossRef]
- Nutley, B.P.; Smith, N.F.; Hayes, A.; Kelland, L.R.; Brunton, L.; Golding, B.T.; Smith, G.C.; Martin, N.M.; Workman, P.; Raynaud, F.I. Preclinical pharmacokinetics and metabolism of a novel prototype DNA-PK inhibitor NU7026. Br. J. Cancer 2005, 93, 1011–1018. [Google Scholar] [CrossRef] [Green Version]
- Belenkov, A.I.; Paiement, J.P.; Panasci, L.C.; Monia, B.P.; Chow, T.Y. An antisense oligonucleotide targeted to human Ku86 messenger RNA sensitizes M059K malignant glioma cells to ionizing radiation, bleomycin, and etoposide but not DNA cross-linking agents. Cancer Res. 2002, 62, 5888–5896. [Google Scholar]
- Marangoni, E.; Le Romancer, M.; Foray, N.; Muller, C.; Douc-Rasy, S.; Vaganay, S.; Abdulkarim, B.; Barrois, M.; Calsou, P.; Bernier, J.; et al. Transfer of Ku86 RNA antisense decreases the radioresistance of human fibroblasts. Cancer Gene Ther. 2000, 7, 339–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinstock, D.M.; Jasin, M. Alternative pathways for the repair of RAG-induced DNA breaks. Mol. Cell. Biol. 2006, 26, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Gottlieb, T.M.; Jackson, S.P. The DNA-dependent protein kinase: Requirement for DNA ends and association with Ku antigen. Cell 1993, 72, 131–142. [Google Scholar] [CrossRef]
- Sahu, B.; Sacui, I.; Rapireddy, S.; Zanotti, K.J.; Bahal, R.; Armitage, B.A.; Ly, D.H. Synthesis and characterization of conformationally preorganized, (R)-diethylene glycol-containing gamma-peptide nucleic acids with superior hybridization properties and water solubility. J. Org. Chem. 2011, 76, 5614–5627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bindra, R.; Sundaram, R.K.; Aiello, R.J.; Marshall, D.; Bourassa, P.; Csengery, J.; Zhang, Q.; Robinson, B.; lopresti-Morrow, L.; Bechtold, J.; et al. Unlocking PARP inhibitor efficacy for HRD-negative cancers using the alphalex tumor targeting platform inhibitor efficacy for HRD-negative cancers using the alphalex tumor targeting platform. J. Clin. Oncol. 2019, 37, e14664. [Google Scholar] [CrossRef]
- Paralkar, V.; Aiello, R.J.; Marshall, D.; Csengery, J.; Bourassa, P.; Zhang, Q.; Robinson, B.S.; Lopresti-Morrow, L.; Bechtold, J.; Tylaska, L.; et al. Abstract 2981: Targeting solid tumor acidic microenvironment with an alphalex PARP inhibitor. Cancer Res. 2019, 79, 2981. [Google Scholar] [CrossRef]
- Bryant, H.E.; Schultz, N.; Thomas, H.D.; Parker, K.M.; Flower, D.; Lopez, E.; Kyle, S.; Meuth, M.; Curtin, N.J.; Helleday, T. Specific killing of BRCA2-deficient tumours with inhibitors of poly(ADP-ribose) polymerase. Nature 2005, 434, 913–917. [Google Scholar] [CrossRef] [PubMed]
- Farmer, H.; McCabe, N.; Lord, C.J.; Tutt, A.N.; Johnson, D.A.; Richardson, T.B.; Santarosa, M.; Dillon, K.J.; Hickson, I.; Knights, C.; et al. Targeting the DNA repair defect in BRCA mutant cells as a therapeutic strategy. Nature 2005, 434, 917–921. [Google Scholar] [CrossRef]
- Mirza, M.R.; Monk, B.J.; Herrstedt, J.; Oza, A.M.; Mahner, S.; Redondo, A.; Fabbro, M.; Ledermann, J.A.; Lorusso, D.; Vergote, I.; et al. Niraparib Maintenance Therapy in Platinum-Sensitive, Recurrent Ovarian Cancer. N. Engl. J. Med. 2016, 375, 2154–2164. [Google Scholar] [CrossRef] [PubMed]
- Coleman, R.L.; Oza, A.M.; Lorusso, D.; Aghajanian, C.; Oaknin, A.; Dean, A.; Colombo, N.; Weberpals, J.I.; Clamp, A.; Scambia, G.; et al. Rucaparib maintenance treatment for recurrent ovarian carcinoma after response to platinum therapy (ARIEL3): A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet 2017, 390, 1949–1961. [Google Scholar] [CrossRef] [Green Version]
- Pujade-Lauraine, E.; Ledermann, J.A.; Selle, F.; Gebski, V.; Penson, R.T.; Oza, A.M.; Korach, J.; Huzarski, T.; Poveda, A.; Pignata, S.; et al. Olaparib tablets as maintenance therapy in patients with platinum-sensitive, relapsed ovarian cancer and a BRCA1/2 mutation (SOLO2/ENGOT-Ov21): A double-blind, randomised, placebo-controlled, phase 3 trial. Lancet Oncol. 2017, 18, 1274–1284. [Google Scholar] [CrossRef] [Green Version]
- Robson, M.; Im, S.A.; Senkus, E.; Xu, B.; Domchek, S.M.; Masuda, N.; Delaloge, S.; Li, W.; Tung, N.; Armstrong, A.; et al. Olaparib for Metastatic Breast Cancer in Patients with a Germline BRCA Mutation. N. Engl. J. Med. 2017, 377, 523–533. [Google Scholar] [CrossRef]
- Robson, M.E.; Tung, N.; Conte, P.; Im, S.A.; Senkus, E.; Xu, B.; Masuda, N.; Delaloge, S.; Li, W.; Armstrong, A.; et al. OlympiAD final overall survival and tolerability results: Olaparib versus chemotherapy treatment of physician’s choice in patients with a germline BRCA mutation and HER2-negative metastatic breast cancer. Ann. Oncol. 2019, 30, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Aiello, R.J.; Gayle, S.; Bechtold, J.; Bourassa, P.; Csengery, J.; Deshpande, K.; Jones, K.; Lopresti-Morrow, L.; Maguire, R.; Marshall, D.; et al. Abstract 6249: CBX-12: A low pH targeting alphalex™-exatecan conjugate for the treatment of solid tumors. Cancer Res. 2020, 80, 6249. [Google Scholar] [CrossRef]
- Pommier, Y. Topoisomerase I inhibitors: Camptothecins and beyond. Nat. Rev. Cancer 2006, 6, 789–802. [Google Scholar] [CrossRef]
- Conroy, T.; Hammel, P.; Hebbar, M.; Ben Abdelghani, M.; Wei, A.C.; Raoul, J.L.; Chone, L.; Francois, E.; Artru, P.; Biagi, J.J.; et al. FOLFIRINOX or Gemcitabine as Adjuvant Therapy for Pancreatic Cancer. N. Engl. J. Med. 2018, 379, 2395–2406. [Google Scholar] [CrossRef]
- Wyatt, L.C.; Lewis, J.S.; Andreev, O.A.; Reshetnyak, Y.K.; Engelman, D.M. Applications of pHLIP Technology for Cancer Imaging and Therapy. Trends Biotechnol. 2017, 35, 653–664. [Google Scholar] [CrossRef]
- Golijanin, J.; Amin, A.; Moshnikova, A.; Brito, J.M.; Tran, T.Y.; Adochite, R.C.; Andreev, G.O.; Crawford, T.; Engelman, D.M.; Andreev, O.A.; et al. Targeted imaging of urothelium carcinoma in human bladders by an ICG pHLIP peptide ex vivo. Proc. Natl. Acad. Sci. USA 2016, 113, 11829–11834. [Google Scholar] [CrossRef] [Green Version]
- Moore, R.M.; Greenberg, M.M. Acid production in the functioning heart under conditions of ischemia and of congestion. Am. J. Physiol. Leg. Content 1937, 118, 217–224. [Google Scholar] [CrossRef]
- Sosunov, E.A.; Anyukhovsky, E.P.; Sosunov, A.A.; Moshnikova, A.; Wijesinghe, D.; Engelman, D.M.; Reshetnyak, Y.K.; Andreev, O.A. pH (low) insertion peptide (pHLIP) targets ischemic myocardium. Proc. Natl. Acad. Sci. USA 2013, 110, 82–86. [Google Scholar] [CrossRef] [Green Version]
- Price, N.L.; Miguel, V.; Ding, W.; Singh, A.K.; Malik, S.; Rotllan, N.; Moshnikova, A.; Toczek, J.; Zeiss, C.; Sadeghi, M.M.; et al. Genetic deficiency or pharmacological inhibition of miR-33 protects from kidney fibrosis. JCI Insight 2019, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Cargo Type | Cargo | Tumor Model | Vehicle |
|---|---|---|---|
| Small Molecules | |||
| Antiproliferative agents | Phallodin [19] | HeLa, JC, M4A4, HT1080 * | WT pHLIP-K(rho) |
| α-Amanitin [20] | HeLa, U2OS, M4A4, MDA-MB-231 * | WT pHLIP | |
| Doxorubicin [21] | MCF-7, MCF-7/ADR * | WT pHLIP | |
| microtubule inhibitor monomethyl auristatin E (MMAE) [22] | HeLa, A431, MDA-MB-23 * NCRr nu/nu mice bearing HeLa or A431 tumors ** | WT pHLIP pHLIP-D25E pHLIP-P20G pHLIP-D14Gla:D25Aad | |
| Paclitaxel [13] | A549 * | WT pHLIP pHLIP-D14Gla:D25Aad | |
| Imaging agents | Alexa750, Cy5.5 [6,7] | BALB/c nude mice bearing HeLa and NM2C5 tumors, TRAMP mice bearing M4A4 tumors ** CRL-2116 tumor bearing C3D2F1 mice ** | WT pHLIP |
| Alexa488, Alexa546, Alexa647, IR680 [23] | 4T1 * BALB/c mice bearing 4T1 tumors and MMTV-PyMT mice ** | WT pHLIP Var3 pHLIP Var7 pHLIP | |
| Indocyanin green (ICG) [24] | HMEpC * BALB/c mice bearing 4T1, MDA-MB-231, A549, LLC, M4A4, HeLa, UM-UC3 and LNCaP tumors ** | Var3 pHLIP | |
| 64Cu, 18F-AlF [9] | BALB/c mice bearing 4T1 tumors ** | WT pHLIP Var3 pHLIP Var7 pHLIP | |
| 99mTC [25] | LLC tumor bearing C57BL/6 mice, PC-3 and LNCaP tumor bearing SCID mice ** | WT pHLIP | |
| Nanoparticles | |||
| Antiproliferative agents | Doxorubicin encapsulated MCM-41 nanoparticles [26] | MCF-7, MCF-7/ADR * | WT pHLIP |
| Ceramide encapsulated liposomes [27] | A549, HeLa * | WT pHLIP | |
| Gramicidin A encapsulated liposomes [28] | A549, HeLa, M4A4 * | WT pHLIP | |
| Genetic material | Plasmid DNA encapsulated dendrigraft poly lysine nanoparticles [29] | Bel-7402 * BALB/c nude mice bearing Bel-7402 tumors ** | WT pHLIP |
| Metallic nanoparticles | Gold nanoparticles [30,31,32] | HeLa, A549, JC BALB/c nude mice bearing HeLa tumors and BALB/c mice bearing JC tumors ** | WT pHLIP |
| Gadolinium nanoparticles [33] | A549 * EMT6 tumors in BALB/c mice ** | WT pHLIP | |
| Peptide Nucleic Acids (PNAs) | |||
| Anti-miRNA | Anti-miR-155 [34] | A549, KB, DLBCL * Nude mice bearing neoplastic B cells derived from enlarged spleens of miR-155LSLtTA mice and mice bearing KB tumors ** | WT pHLIP |
| Anti-miR-21 [34,35] | A549 * Orthotopic and heterotopic LLC tumors in C57BL6 mice ** | WT pHLIP Var3 pHLIP | |
| Anti-lncRNA | HOX transcript antisense RNA targeting PNA [36] | A2780P, A2780_CR5, KURAMOCHI, SKBR-3, MCF-7, MDA-MB-231 * BALB/c-nu/nu mice bearing A2780_CR5 tumors ** | WT pHLIP |
| Antisense | αKU80 [37] | A549, DLD1-BRCA2KO, EMT-6 * BALBc/Rw mice bearing EMT-6 tumors, athymic nu/nu mice bearing DLD1-BRCA2KO tumors ** | WT pHLIP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dharmaratne, N.U.; Kaplan, A.R.; Glazer, P.M. Targeting the Hypoxic and Acidic Tumor Microenvironment with pH-Sensitive Peptides. Cells 2021, 10, 541. https://doi.org/10.3390/cells10030541
Dharmaratne NU, Kaplan AR, Glazer PM. Targeting the Hypoxic and Acidic Tumor Microenvironment with pH-Sensitive Peptides. Cells. 2021; 10(3):541. https://doi.org/10.3390/cells10030541
Chicago/Turabian StyleDharmaratne, Nayanthara U., Alanna R. Kaplan, and Peter M. Glazer. 2021. "Targeting the Hypoxic and Acidic Tumor Microenvironment with pH-Sensitive Peptides" Cells 10, no. 3: 541. https://doi.org/10.3390/cells10030541