
Progression of Watermelon Bud Necrosis Virus Infection in Its Vector, Thrips palmi




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Developing a Homogeneous Population of Thrips palmi
2.2. Establishing Watermelon Bud Necrosis Virus Culture
2.3. Generating a Viruliferous Thrips palmi Population
2.4. Confirming WBNV Infection by RT-PCR
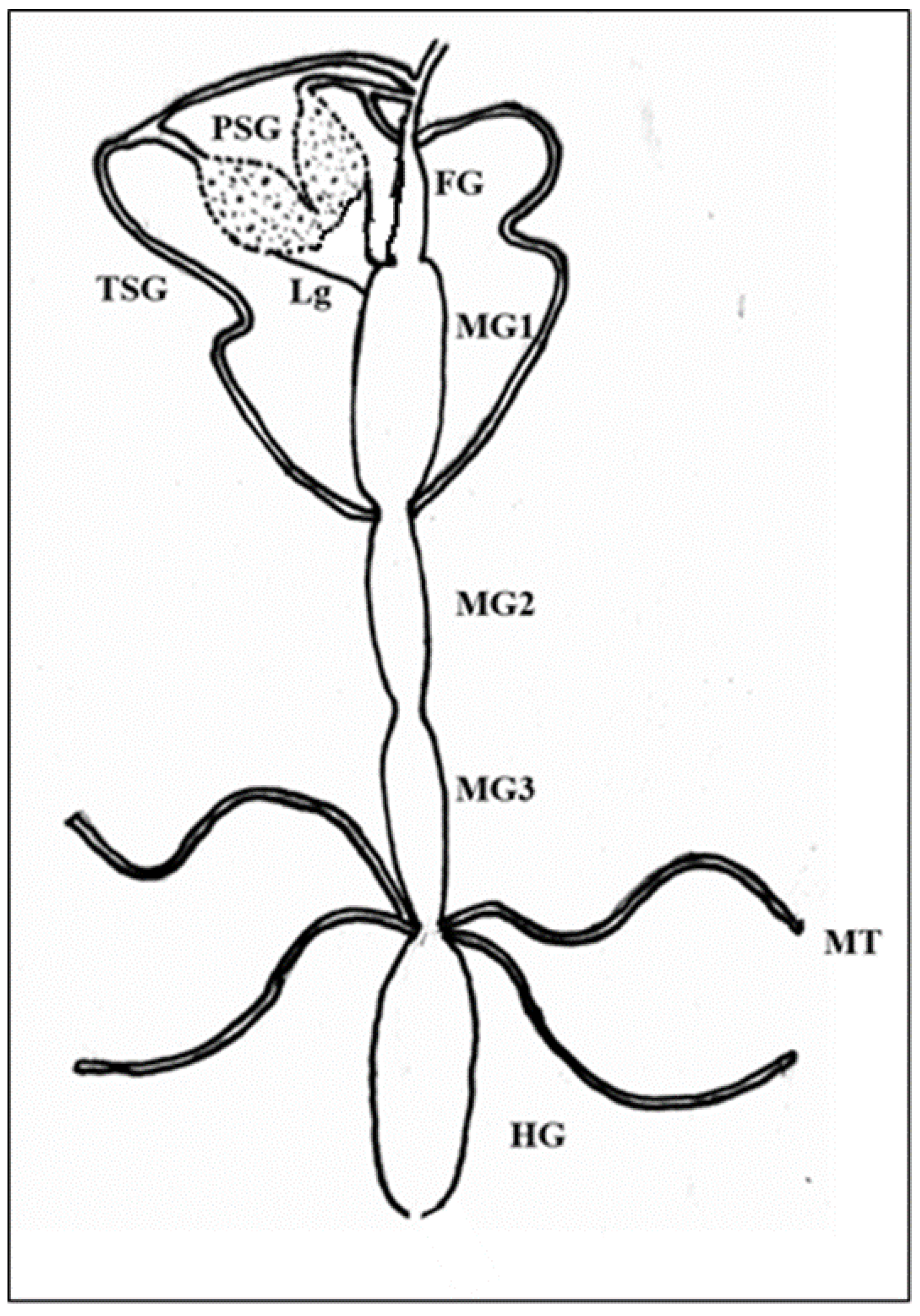
2.5. Dissection and Immunolabeling of Alimentary Canal and Salivary Glands of Thrips palmi
2.6. Localization of WBNV N Protein by Confocal Laser Scanning Microscopy
2.7. Quantification of the Fluorescence Signal
3. Results
3.1. Thrips palmi Population
3.2. Watermelon Bud Necrosis Virus Culture
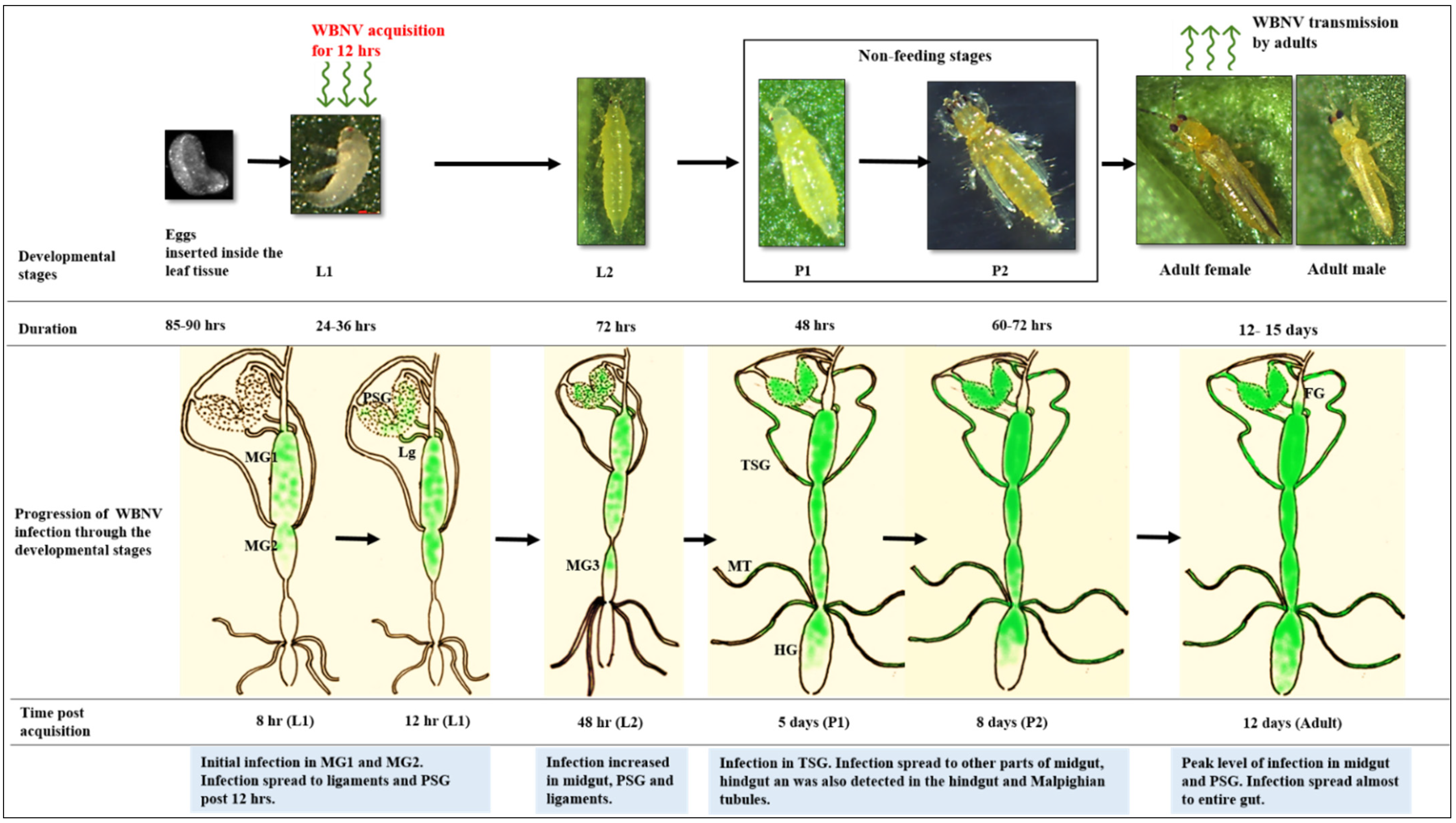
3.3. Developmental Stages and Viruliferous Population of Thrips palmi
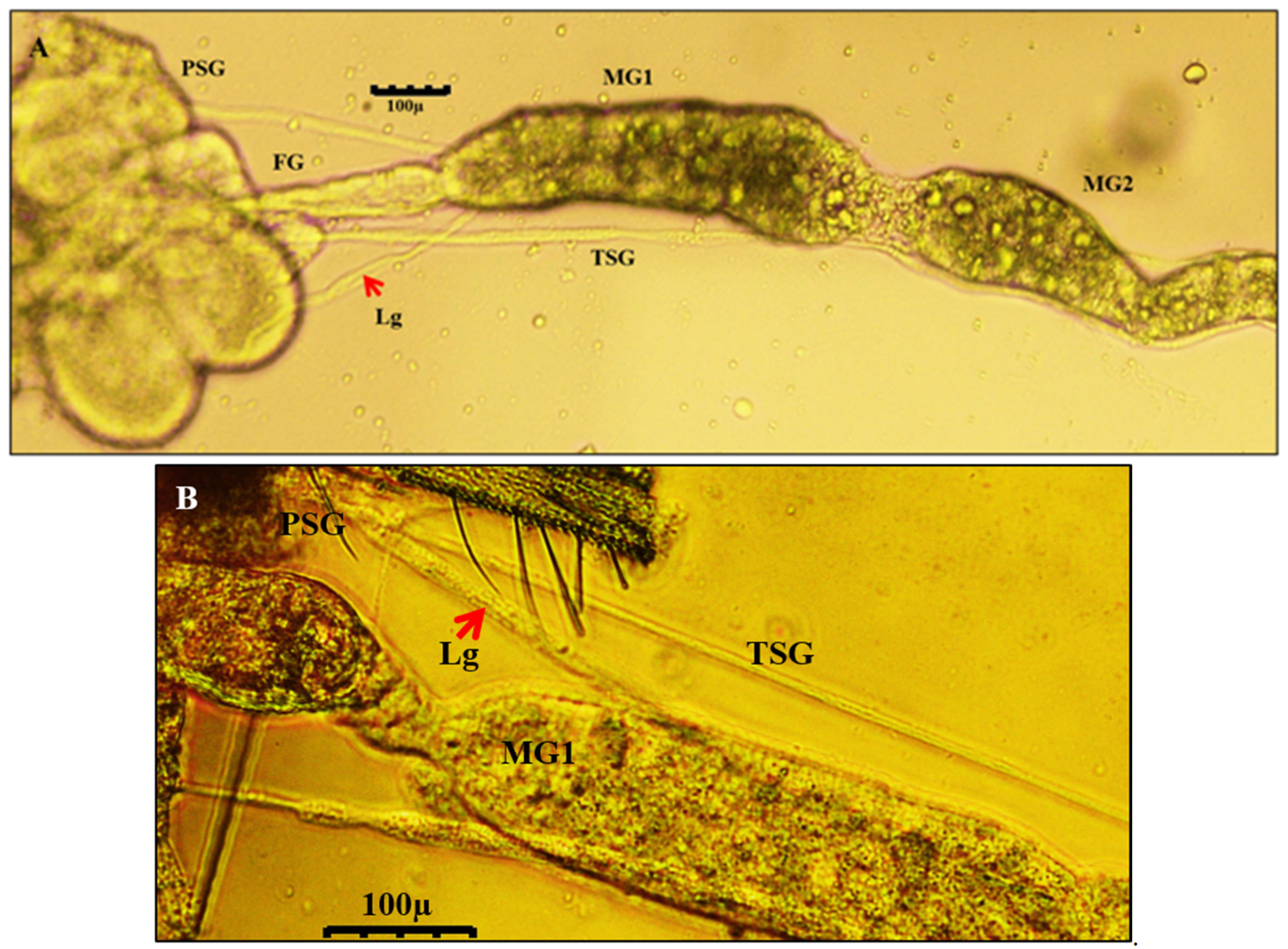
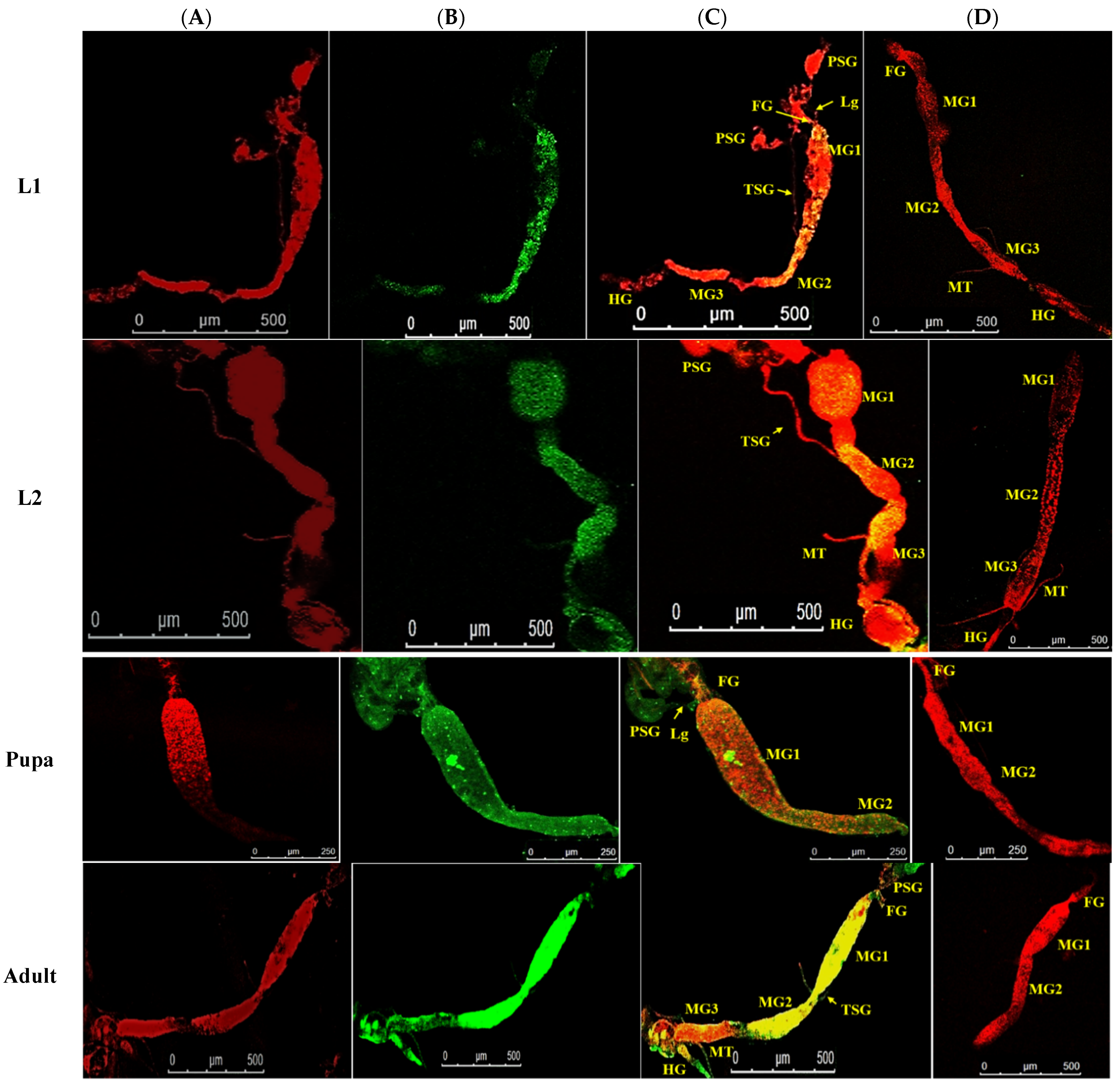
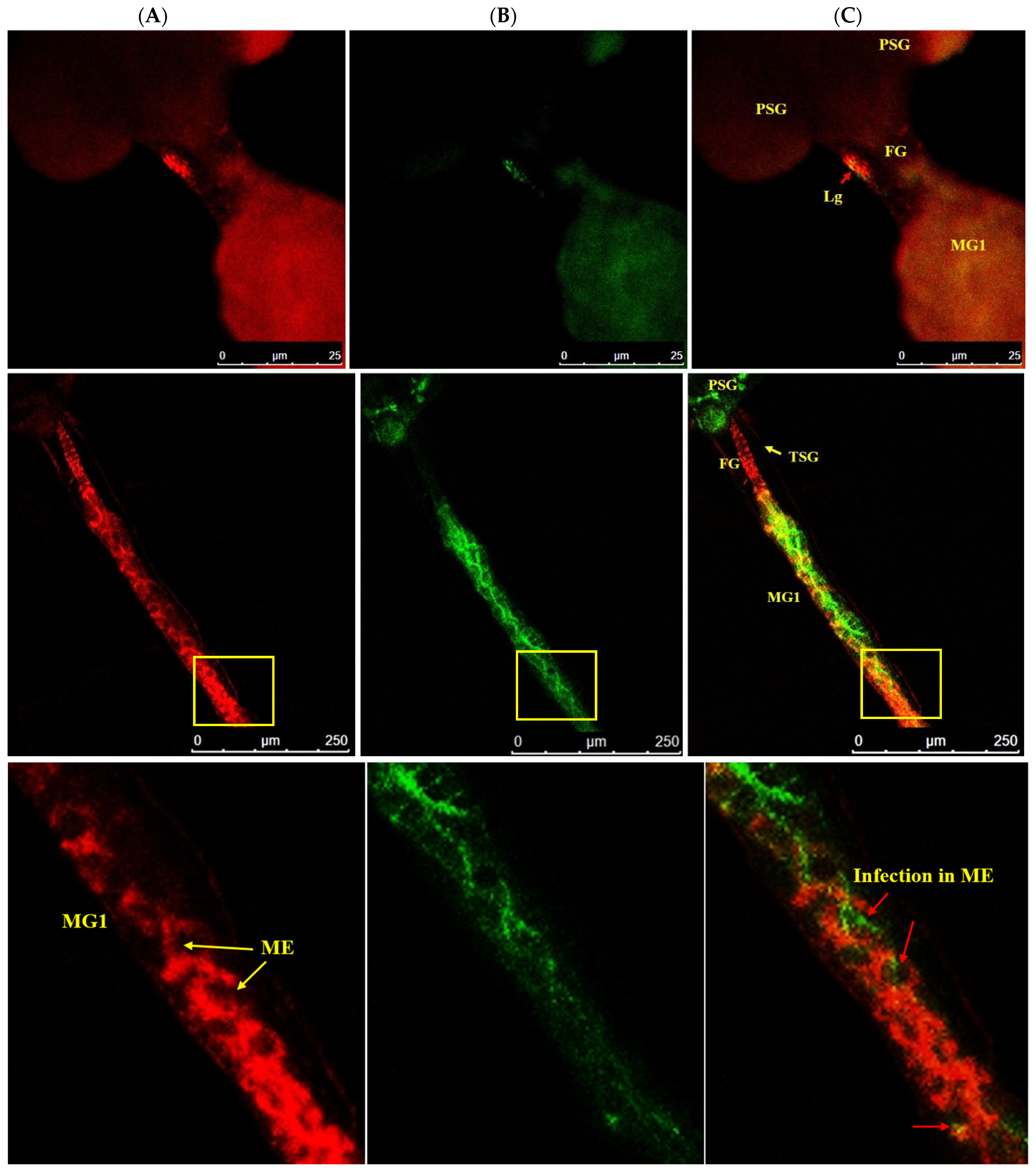
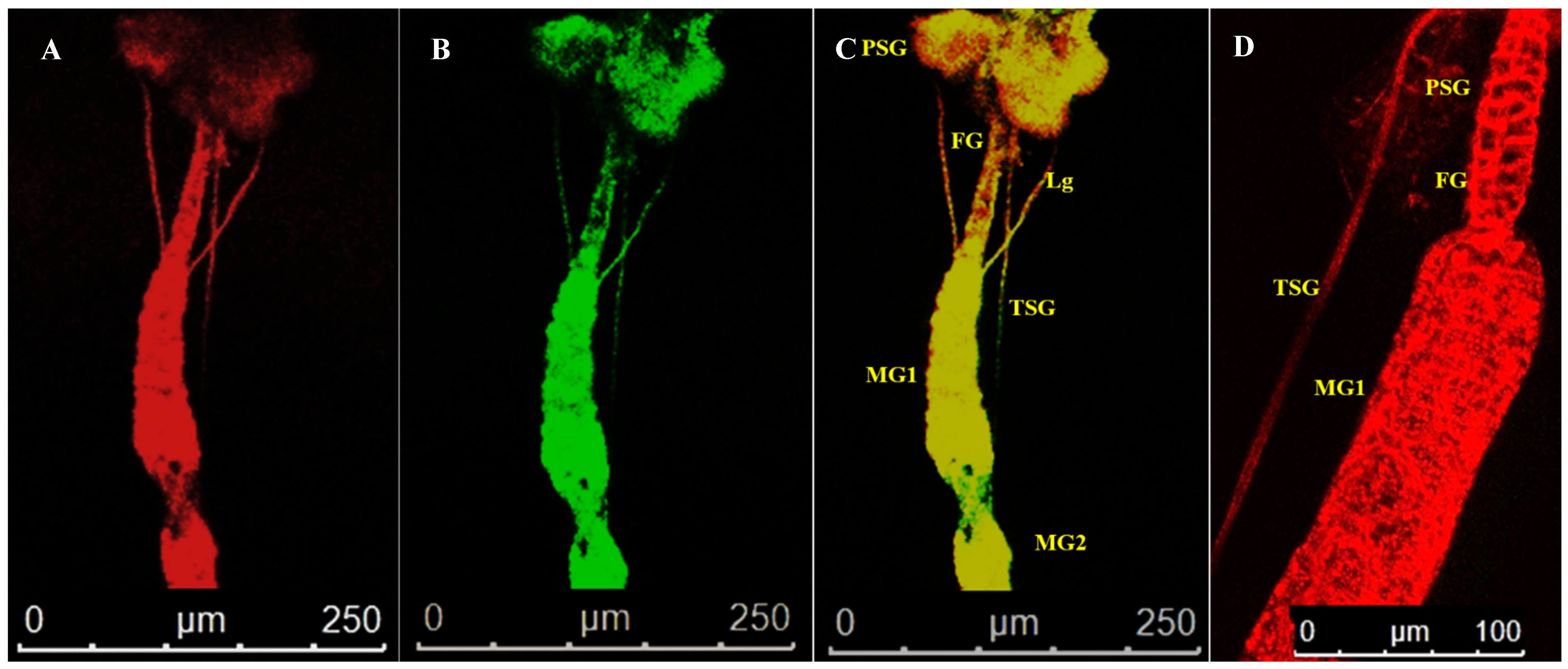
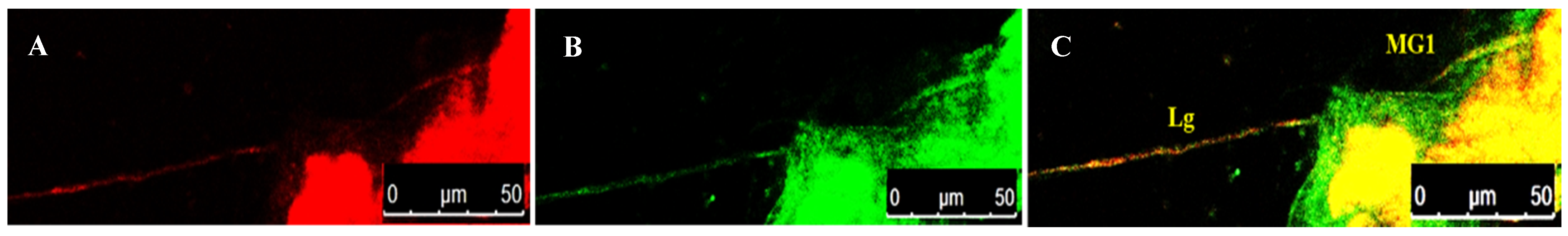
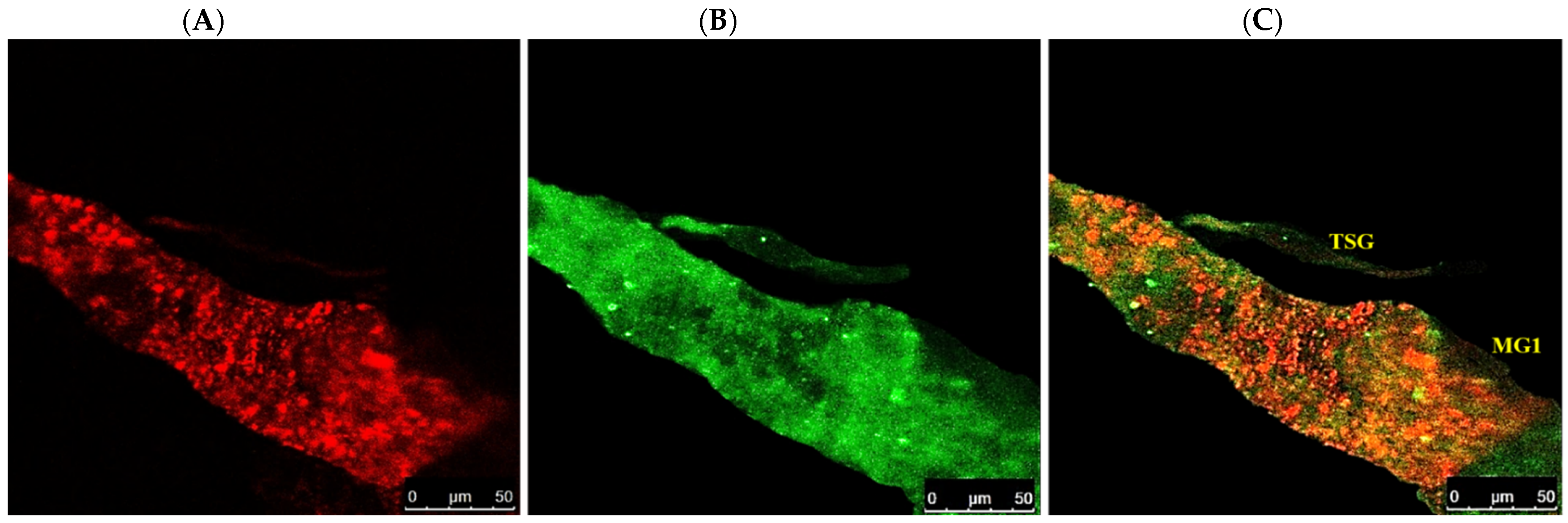
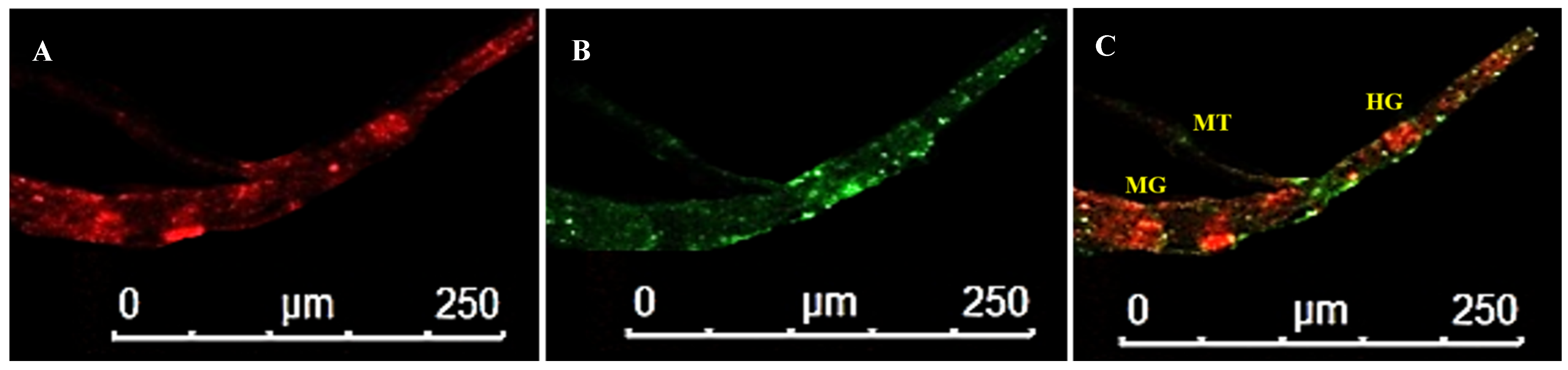
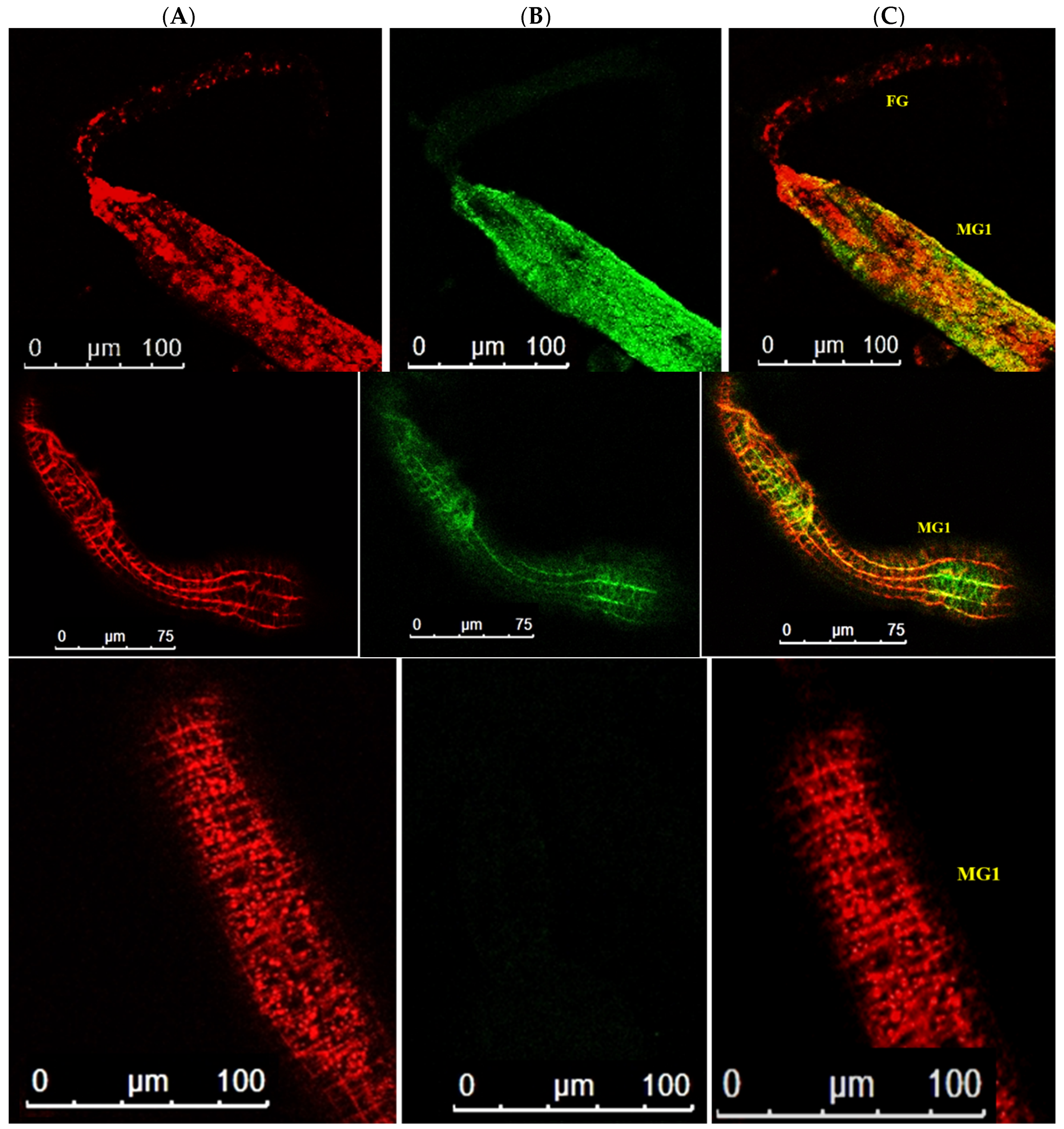
3.4. Localization of WBNV in Alimentary Canal and Salivary Glands of Thrips palmi during Different Developmental Stages
3.4.1. Localization of WBNV in L1 Stage
3.4.2. Localization of WBNV in L2 Stage
3.4.3. Localization of WBNV in Pupal Stages
3.4.4. Localization of WBNV in Adult Stage
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics Declaration
References
- Mound, L.A. Thysanoptera (Thrips) of the World a Checklist. 2015. Available online: http://www.ento.csiro.au/thysanoptera/worldthrips.php (accessed on 1 December 2020).
- Childers, C.C.; Achor, D.S. Thrips feeding and oviposition injuries to economic plants, subsequent damage and host responses to infestation. In Thrips Biology and Management; Parker, B.L., Skinner, M., Lewis, T., Eds.; Springer: New York, NY, USA, 1995; pp. 31–51. [Google Scholar]
- Whitfield, A.E.; Ullman, D.E.; German, T.L. Tospovirus-thrips interactions. Annu. Rev. Phytopathol. 2005, 43, 459–489. [Google Scholar] [CrossRef]
- Rotenberg, D.; Jacobson, A.L.; Schneweis, D.J.; Whitfield, A.E. Thrips transmission of tospoviruses. Curr. Opin. Virol. 2015, 15, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Dey, D.; Basavaraj, T.; Mandal, B.; Jain, R.K. Thrips as the vectors of tospoviruses in Indian agriculture. In A Century of Plant Virology in India; Mandal, B., Rao, G.P., Baranwal, V.K., Jain, R.K., Eds.; Springer: Singapore, 2017; pp. 537–556. [Google Scholar]
- Sharman, M.; Thomas, J.E.; Tree, D.; Persley, D.M. Natural host range and thrips transmission of capsicum chlorosis virus in Australia. Australas. Plant Pathol. 2020, 49, 45–51. [Google Scholar] [CrossRef]
- Nagata, T.; Inoue-Nagata, A.K.; Smid, H.M.; Goldbach, R.; Peters, D. Tissue tropism related to vector competence of Frankliniella occidentalis for tomato spotted wilt tospovirus. J. Gen. Virol. 1999, 80, 507–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, A.; Basavaraj, Y.B.; Jangra, S.; Das, A. Exposure to watermelon bud necrosis virus and groundnut bud necrosis virus alters the life history traits of their vector, Thrips palmi (Thysanoptera: Thripidae). Arch. Virol. 2019, 164, 2799–2804. [Google Scholar] [CrossRef]
- Montero-Astúa, M.; Ullman, D.E.; Whitfield, A.E. Salivary gland morphology, tissue tropism and the progression of tospovirus infection in Frankliniella occidentalis. Virology 2016, 493, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Nalam, V.J.; Yu, I.C.; Nachappa, P. Vector competence of thrips species to transmit soybean vein necrosis virus. Front. Microbiol. 2019, 10, 431. [Google Scholar] [CrossRef]
- Moritz, G.; Kumm, S.; Mound, L. Tospovirus transmission depends on thrips ontogeny. Virus Res. 2004, 100, 143–149. [Google Scholar] [CrossRef]
- Ghosh, A.; Jagdale, S.S.; Basavaraj; Dietzgen, R.G.; Jain, R.K. Genetics of Thrips palmi (Thysanoptera: Thripidae). J. Pest Sci. 2020, 93, 27–39. [Google Scholar] [CrossRef]
- Bhatti, J.S. Species of the genus Thrips from India (Thysanoptera). Syst. Entomol. 1980, 5, 109–166. [Google Scholar] [CrossRef]
- Johnson, M.W. Population Trends of a Newly Introduced Species, Thrips palmi (Thysanoptera: Thripidae), on commercial watermelon plantings in Hawaii. J. Econ. Entomol. 1986, 79, 718–720. [Google Scholar] [CrossRef]
- Guyot, J. Review and first observation in Guadeloupe on Thrips Palmi Karny. Agronomie 1998, 8, 565–575. [Google Scholar] [CrossRef] [Green Version]
- Pantoja, A.; Segarra, A.; Ruiz, H.; Medina-Gaud, S. Thrips palmi (Thysanoptera: Thripidae): A new insect pest for Puerto Rico. J. Agric. Univ. Puerto Rico. 1988, 2, 327–329. [Google Scholar]
- Cooper, B. Infestation of the southern yellow thrips Thrips palmi, in vegetables. J. Agric. Soc. Trinidad. Tobago 1991, 88, 37–38. [Google Scholar]
- Palmer, J.M. Thrips (Thysanoptera) from Pakistan to the Pacific: A review. Bull. Br. Mus. (Nat. Hist.) Entomol. Ser. 1992, 61, 76. [Google Scholar]
- Monteiro, R.C.; Zucchi, R.A.; Mound, L.A. Record of Thrips palmi Karny, 1925 (Thysanoptera, Thripidae) in the state of Sao Paulo, Brazil. Rev. Agric. 1995, 70, 53–55. [Google Scholar]
- EPPO. PQR Database; European and Mediterranean Plant Protection Organization: Paris, France, 2014; Available online: http://www.eppo.int/DATABASES/pqr/pqr.htm (accessed on 1 December 2020).
- Jain, R.K.; Pappu, H.R.; Pappu, S.S.; Reddy, M.K.; Vani, A. Watermelon bud necrosis tospovirus is a distinct virus species belonging to serogroup IV. Arch. Virol. 1998, 143, 1637–1644. [Google Scholar] [CrossRef]
- Kunkalikar, S.R.; Poojari, S.; Arun, B.M.; Rajagopalan, P.A.; Chen, T.C.; Yeh, S.D.; Naidu, R.A.; Zehr, U.B.; Ravi, K.S. Importance and genetic diversity of vegetable-infecting tospoviruses in India. Phytopathology 2011, 101, 367–376. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Krishnareddy, M. Watermelon bud necrosis: A new tospovirus disease. Acta Hortic. 1996, 431, 68–77. [Google Scholar] [CrossRef]
- Cluever, J.D.; Smith, H.A. A photo-based key of thrips (Thysanoptera) associated with horticultural crops in Florida. Fla. Entomol. 2017, 100, 454–467. [Google Scholar] [CrossRef]
- Jangra, S.; Mittal, A.; Dhall, H.; Jain, R.K.; Ghosh, A. A multiplex PCR assay for rapid identification of major tospovirus vectors reported in India. BMC Genom. 2020, 21, 4–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandal, B.; Pappu, H.R.; Culbreath, A.K. Factors affecting mechanical transmission of Tomato spotted wilt virus to peanut (Arachis hypogaea). Plant Dis. 2001, 85, 1259–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jangra, S.; Dhall, H.; Aggarwal, S.; Mandal, B.; Jain, R.K.; Ghosh, A. An observation on the embryonic development in Thrips palmi (Thysanoptera: Thripidae) eggs obtained by an artificial oviposition setup. J. Asia-Pac. Entomol. 2020, 23, 492–497. [Google Scholar] [CrossRef]
- Basavaraj; Siwach, J.; Pandey, R.; Kumar, A.; Mandal, B.; Jain, R.K. Multiplex RT-PCR assay system for simultaneous detection of capsicum chlorosis virus, groundnut bud necrosis orthotospovirus, and watermelon bud necrosis orthotospovirus, and identification of new natural host. In International Conference of Virology: Global Viral Epidemics a Challenging Threat; PGIMER: Chandigarh, India, 2018; pp. 82–83. [Google Scholar]
- Schneider, C.; Rasband, W.; Eliceiri, K. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- de Assis Filho, F.M.; Naidu, R.A.; Deom, C.M.; Sherwood, J.L. Dynamics of Tomato spotted wilt virus replication in the alimentary canal of two thrips species. Phytopathology 2002, 92, 729–733. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, S.; Fujisawa, I.; Ohnishi, J.; Hosokawa, D.; Tomaru, K. Localization of tomato spotted wilt tospovirus in larvae and pupae of the insect vector Thrips setosus. Phytopathology 1996, 86, 1199–1203. [Google Scholar] [CrossRef] [Green Version]
- Dhall, H.; Jangra, S.; Basavaraj, Y.B.; Ghosh, A. Host plant influences life cycle, reproduction, feeding, and vector competence of Thrips palmi (Thysanoptera: Thripidae), a vector of tospoviruses. Phytoparasitica 2021. [Google Scholar] [CrossRef]
- Ullman, D.E.; German, T.L.; Sherwood, J.L.; Westcot, D.M.; Cantone, F.A. Tospovirus replication in insect vector cells: Immunocytochemical evidence that the nonstructural protein encoded by the S RNA of tomato spotted wilt tospovirus is present in thrips vector cells. Phytopathology 1993, 83, 456. [Google Scholar] [CrossRef]
- Bandla, M.D.; Campbell, L.R.; Ullman, D.E.; Sherwood, J.L. Interaction of tomato spotted wilt tospovirus (TSWV) glycoproteins with a thrips midgut protein, a potential cellular receptor for TSWV. Phytopathology 1998, 88, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Buchon, N.; Osman, D.; David, F.P.A.; Yu Fang, H.; Boquete, J.-P.; Deplancke, B.; Lemaitre, B. Morphological and molecular characterization of adult midgut compartmentalization in Drosophila. Cell Rep. 2013, 3, 1725–1738. [Google Scholar] [CrossRef] [Green Version]
- Marianes, A.; Spradling, A.C. Physiological and stem cell compartmentalization within the Drosophila midgut. eLife 2013, 2, e00886. [Google Scholar] [CrossRef]
- O’Brien, L.E. Regional specificity in the Drosophila midgut: Setting boundaries with stem cells. Cell Stem Cell. 2013, 13, 375–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagata, T. Competence and Specificity of Thrips in the Transmission of Tomato Spotted Wilt Virus. Ph.D. thesis, Agricultural University of Wageningen, Wageningen, The Netherlands, 1999. [Google Scholar]
- Ullman, D.E.; Westcot, D.M.; Chenault, K.D.; Sherwood, J.L.; German, T.L.; Bandla, M.D.; Cantone, F.A.; Duer, H. Compartmentalization, intracellular transport, and autophagy of tomato spotted wilt tospovirus proteins in infected thrips cells. Phytopathology 1995, 85, 644–654. [Google Scholar] [CrossRef]
- Ullman, D.E.; Cho, J.J.; Mau, R.F.L.; Hunter, W.B.; Wescot, D.M.; Custer, D. Thrips-tomato spotted wilt virus interactions: Morphological, behavioral and cellular components influencing thrips transmission. Adv. Dis. Vector Res. 1992, 9, 195–240. [Google Scholar]
- Medeiros, R.B.; Ullman, D.E.; Sherwood, J.L.; German, T.L. Immunoprecipitation of a 50-kDa protein: A candidate receptor component for tomato spotted wilt tospovirus (Bunyaviridae) in its main vector, Frankliniella occidentalis. Virus Res. 2000, 67, 109–118. [Google Scholar] [CrossRef]
- Billingsley, P.F.; Lehane, M.J. Structure and ultrastructure of the insect midgut. In Biologyof the Insect Midgut; Billingsley, P.F., Lehane, M.J., Eds.; Springer: Dordrecht, The Netherlands, 1996; pp. 3–30. [Google Scholar]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghosh, A.; Priti; Mandal, B.; Dietzgen, R.G. Progression of Watermelon Bud Necrosis Virus Infection in Its Vector, Thrips palmi. Cells 2021, 10, 392. https://doi.org/10.3390/cells10020392
Ghosh A, Priti, Mandal B, Dietzgen RG. Progression of Watermelon Bud Necrosis Virus Infection in Its Vector, Thrips palmi. Cells. 2021; 10(2):392. https://doi.org/10.3390/cells10020392
Chicago/Turabian StyleGhosh, Amalendu, Priti, Bikash Mandal, and Ralf G. Dietzgen. 2021. "Progression of Watermelon Bud Necrosis Virus Infection in Its Vector, Thrips palmi" Cells 10, no. 2: 392. https://doi.org/10.3390/cells10020392