Impacts of Tamarix (L.) Litter and Mycorrhizal Amendments on Baccharis salicifolia (Ruiz & Pav.) Pers. Competitiveness and Mycorrhizal Colonization

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil and Plant Material Collection and Preparation
2.2. Experimental Setup
2.3. Growing Conditions
2.4. Measurements
2.5. Mycorrhizal Root Preparation and Analysis
2.6. Data Analysis
3. Results
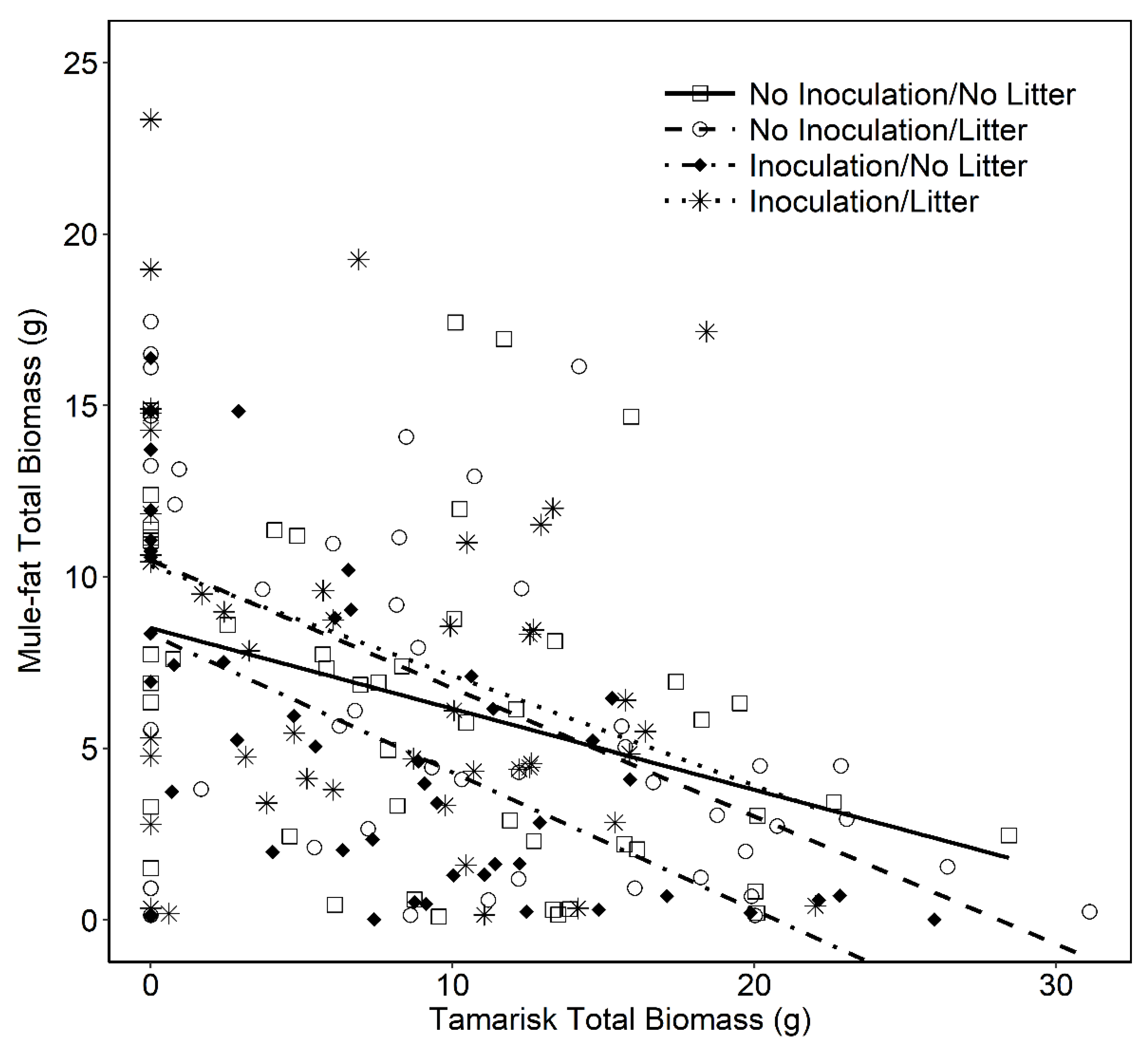
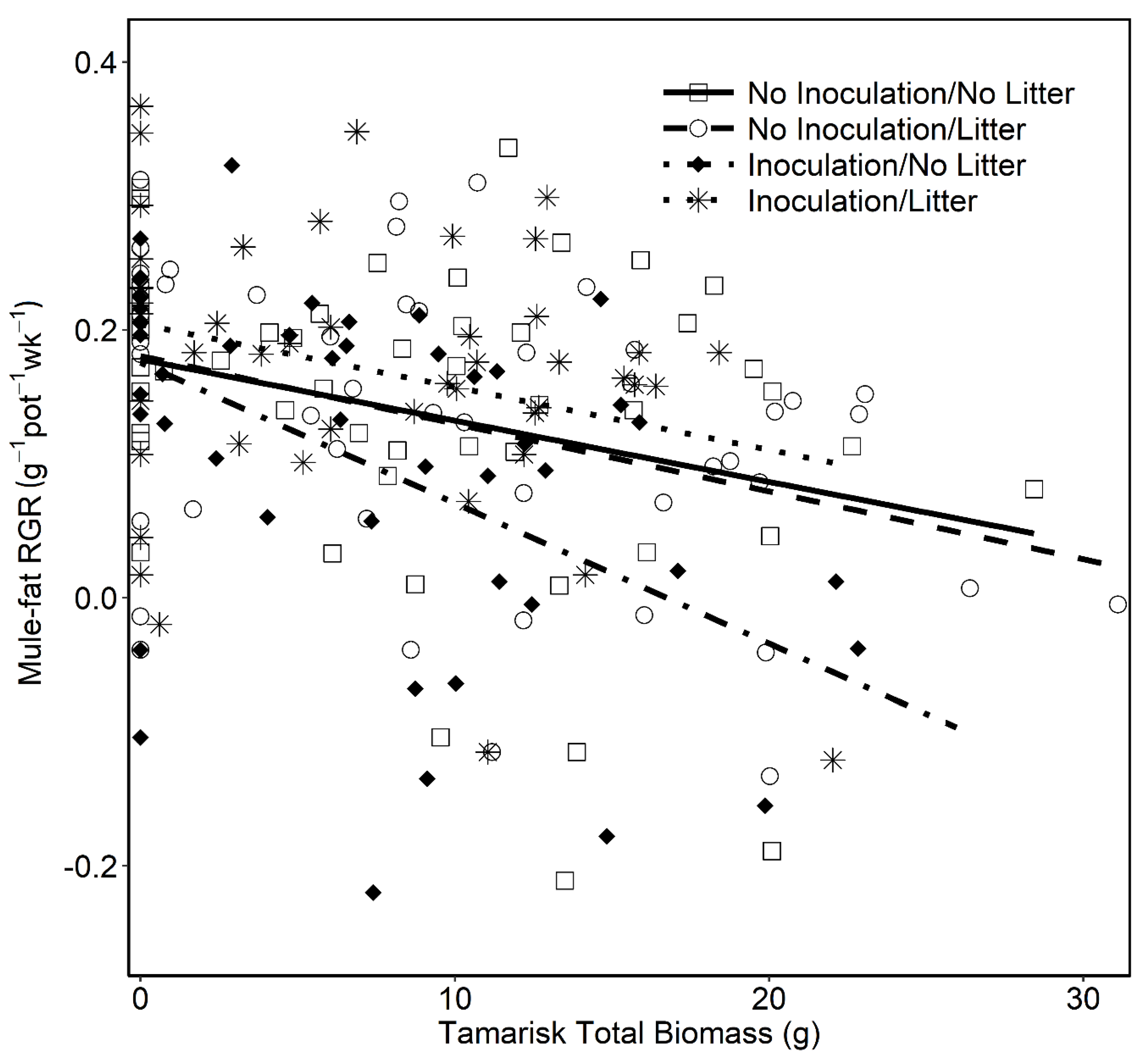
3.1. Mule-fat Growth under Different Soil-Litter Treatments in Competition with Tamarisk
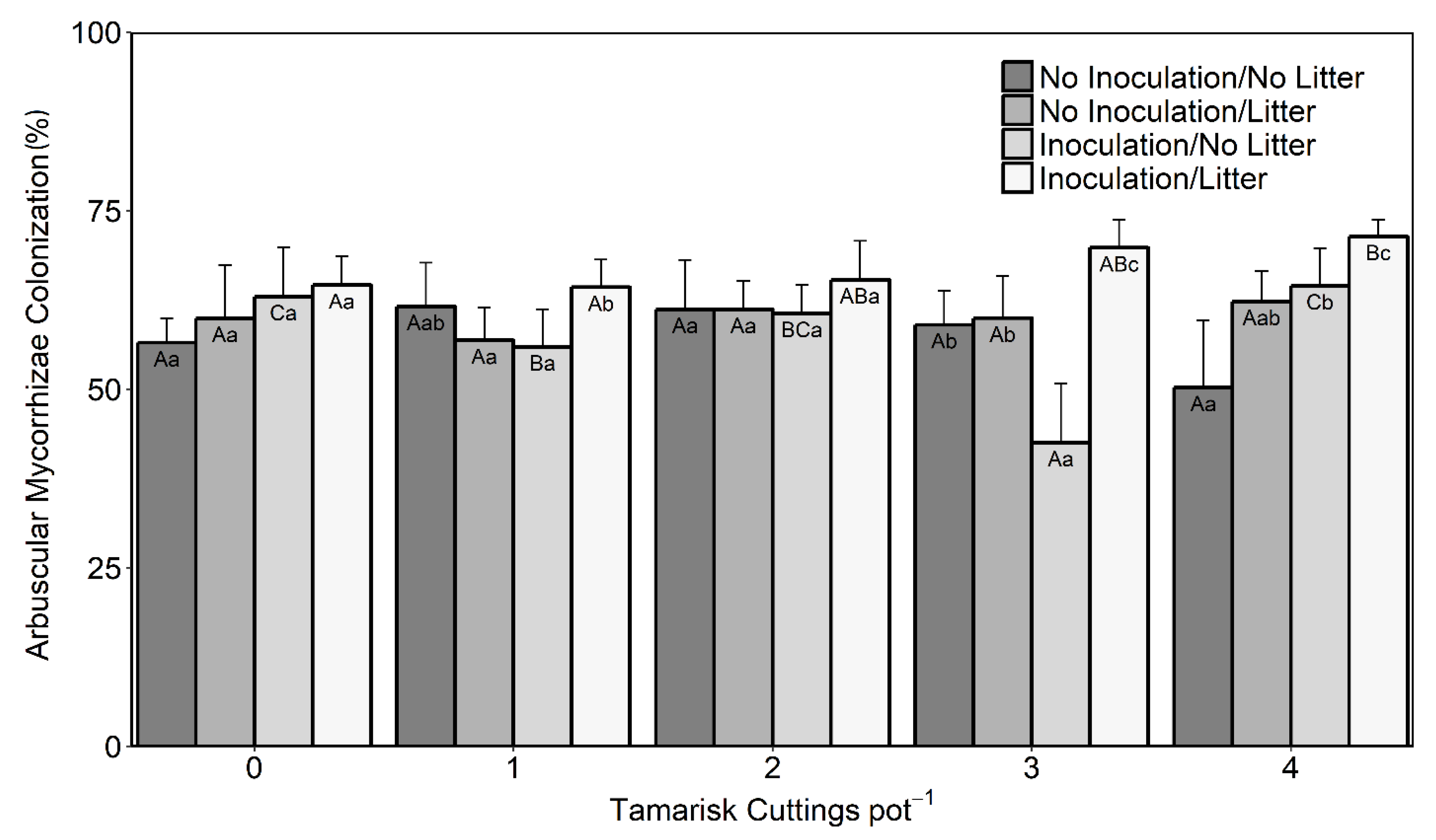
3.2. Arbuscular Mycorrhizal Fungi Colonization of Mule-Fat
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stromberg, J.C.; Lite, S.J.; Marler, R.; Paradzick, C.; Shafroth, P.B.; Shorrock, D.; White, J.M.; White, M.S. Altered stream-flow regimes and invasive plant species: The Tamarix case. Glob. Ecol. Biogeogr. 2007, 16, 381–393. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Arico, S.; Aronson, J.; Baron, J.S.; Bridgewater, P.; Cramer, V.A.; Epstein, P.R.; Ewel, J.J.; Klink, C.A.; Lugo, A.E.; et al. Novel ecosystems: Theoretical and management aspects of the new ecological world order. Glob. Ecol. Biogeogr. 2006, 15, 1–7. [Google Scholar] [CrossRef]
- Robinson, T.W. Introduction, Spread, and Areal Extent of Saltcedar (Tamarix) in the Western States; Geological Survey Professional Paper 491-A; United States Geological Survey: Washington, DC, USA, 1965.
- Zavaleta, E. The Economic Value of Controlling an Invasive Shrub. AMBIO J. Hum. Environ. 2000, 29, 462–467. [Google Scholar] [CrossRef]
- Drus, G.M.; Dudley, T.L.; D’Antonio, C.M.; Even, T.J.; Brooks, M.L.; Matchett, J.R. Synergistic interactions between leaf beetle herbivory and fire enhance tamarisk (Tamarix spp.) mortality. Biol. Control 2014, 77, 29–40. [Google Scholar] [CrossRef]
- Ohmart, R.D.; Anderson, B.W.; Hunter, W.C. The Ecology of the Lower Colorado River from Davis Dam to the Mexico-United States International Boundary: A Community Profile; U.S. Fish and Wildlife Service Biological Report 85(7.19); U.S. Fish and Wildlife Service: Springfield, VA, USA, 1988.
- van Riper, C.; Paxton, K.L.; O’Brien, C.; Shafroth, P.B.; McGrath, L.J. Rethinking avian response to Tamarix on the lower Colorado River: A Threshold Hypothesis. Restor. Ecol. 2008, 16, 155–167. [Google Scholar] [CrossRef]
- Bateman, H.L.; Ostoja, S.M. Invasive woody plants affect the composition of native lizard and small mammal communities in riparian woodlands. Anim. Conserv. 2012, 15, 294–304. [Google Scholar] [CrossRef]
- Bean, D.; Dudley, T.; Hultine, K.R. Bring on the Beetles: The history and impact of tamarisk biological control. In Tamarix: A Case Study of Ecological Change in the American West; Sher, A., Quigley, M.F., Eds.; Oxford University Press: New York, NY, USA, 2013; pp. 377–403. ISBN 978-0-19-989820-6. [Google Scholar]
- Harms, R.S.; Hiebert, R.D. Vegetation response following invasive tamarisk (Tamarix spp.) Removal and Implications for Riparian Restoration. Restor. Ecol. 2006, 14, 461–472. [Google Scholar] [CrossRef]
- Kennard, D.; Louden, N.; Gemoets, D.; Ortega, S.; González, E.; Bean, D.; Cunningham, P.; Johnson, T.; Rosen, K.; Stahlke, A. Tamarix dieback and vegetation patterns following release of the northern tamarisk beetle (Diorhabda carinulata) in western Colorado. Biol. Control 2016, 101, 114–122. [Google Scholar] [CrossRef]
- Ohrtman, M.K.; Sher, A.A.; Lair, K.D. Quantifying soil salinity in areas invaded by Tamarix spp. J. Arid Environ. 2012, 85, 114–121. [Google Scholar] [CrossRef]
- Ladenburger, C.G.; Hild, A.L.; Kazmer, D.J.; Munn, L.C. Soil salinity patterns in Tamarix invasions in the Bighorn Basin, Wyoming, USA. J. Arid Environ. 2006, 65, 111–128. [Google Scholar] [CrossRef]
- Glenn, E.P.; Morino, K.; Nagler, P.L.; Murray, R.S.; Pearlstein, S.; Hultine, K.R. Roles of saltcedar (Tamarix spp.) and capillary rise in salinizing a non-flooding terrace on a flow-regulated desert river. J. Arid Environ. 2012, 79, 56–65. [Google Scholar] [CrossRef]
- Beauchamp, V.B.; Stromberg, J.C.; Stutz, J.C. Interactions between Tamarix ramosissima (saltcedar), Populus fremontii (cottonwood), and mycorrhizal fungi: effects on seedling growth and plant species coexistence. Plant Soil 2005, 275, 221–231. [Google Scholar] [CrossRef]
- Uselman, S.M.; Snyder, K.A.; Blank, R.R. Insect biological control accelerates leaf litter decomposition and alters short-term nutrient dynamics in a Tamarix-invaded riparian ecosystem. Oikos 2011, 120, 409–417. [Google Scholar] [CrossRef]
- Shafroth, P.B.; Beauchamp, V.B.; Briggs, M.K.; Lair, K.; Scott, M.L.; Sher, A. Planning riparian restoration in the context of Tamarix control in western North America. Restor. Ecol. 2008, 16, 97–112. [Google Scholar] [CrossRef]
- Taylor, J.P.; Mcdaniel, K.C. Restoration of Saltcedar (Tamarix Sp.)-infested floodplains on the Bosque Del Apache National Wildlife Refuge. Weed Technol. 1998, 12, 345–352. [Google Scholar] [CrossRef]
- Sherry, R.A.; Shafroth, P.B.; Belnap, J.; Ostoja, S.; Reed, S.C. Germination and growth of native and invasive plants on soil associated with biological control of tamarisk ( Tamarix spp.). Invasive Plant Sci. Manag. 2016, 9, 290–307. [Google Scholar]
- Dreesen, D.; Harrington, J.; Subirge, T.; Stewart, P.; Fenchel, G. Riparian Restoration in the Southwest—Species Selection, Propagation, Planting Methods, and Case Studies. In National Proceedings: Forest and Conservation Nursery Associations-1999, 2000, and 2001. Proceedings RMRS-P-24; Dumroese, R.K., Riley, L.E., Landis, T.D., Eds.; U.S. Department of Agriculture-Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2002; pp. 253–272. [Google Scholar]
- Beauchamp, V.B.; Walz, C.; Shafroth, P.B. Salinity tolerance and mycorrhizal responsiveness of native xeroriparian plants in semi-arid western USA. Appl. Soil Ecol. 2009, 43, 175–184. [Google Scholar] [CrossRef]
- Younginger, B.; Barnouti, J.; Moon, D.C. Interactive effects of mycorrhizal fungi, salt stress, and competition on the herbivores of Baccharis halimifolia. Ecol. Entomol. 2009, 34, 580–587. [Google Scholar] [CrossRef]
- Miyamoto, S.; Martinez, I.; Padilla, M.; Portillo, A.; Ornelas, D. Landscape Plant Lists for Salt Toler-Ance Assessment. Available online: http://www.epwu.org/reclaimed_water/plant_list.pdf (accessed on 27 October 2012).
- Szaro, R.C.; Arizona, U.; Arboretum., B.T.S. Riparian forest and scrubland community types of Arizona and New Mexico. Desert Plants 1989, 9, 70–138. [Google Scholar]
- Bagstad, K.J.; Lite, S.J.; Stromberg, J.C. Vegetation, soils, and hydrogeomorphology of riparian patch types of a dryland river. West. N. Am. Nat. 2006, 66, 23–44. [Google Scholar]
- Glenn, E.P.; Tanner, R.; Mendez, S.; Kehret, T.; Moore, D.; Garcia, J.; Valdes, C. Growth rates, salt tolerance and water use characteristics of native and invasive riparian plants from the delta of the Colorado River, Mexico. J. Arid Environ. 1998, 40, 281–294. [Google Scholar] [CrossRef]
- Vandersande, M.W.; Glenn, E.P.; Walworth, J.L. Tolerance of five riparian plants from the lower Colorado River to salinity drought and inundation. J. Arid Environ. 2001, 49, 147–159. [Google Scholar] [CrossRef]
- Horton, J.S.; Mounts, F.C.; Kraft, J.M. Seed germination and seedling establishment of phreatophyte species. Stn. Pap. Rocky Mt. For. Range Exp. Stn. 1960, 48, 26. [Google Scholar]
- Boldt, P.E.; Robbins, T.O. Phytophagous and Flower-Visiting Insect Fauna of Baccharis salicifolia (Asteraceae) in the Southwestern United States and Northern Mexico. Environ. Entomol. 1990, 19, 515–523. [Google Scholar] [CrossRef]
- Levine, C.M.; Stromberg, J.C. Effects of flooding on native and exotic plant seedlings: Implications for restoring south-western riparian forests by manipulating water and sediment flows. J. Arid Environ. 2001, 49, 111–131. [Google Scholar]
- Lesica, P.; DeLuca, T.H. Is tamarisk allelopathic? Plant Soil 2004, 267, 357–365. [Google Scholar]
- Meinhardt, K.A.; Gehring, C.A. Disrupting mycorrhizal mutualisms: A potential mechanism by which exotic tamarisk outcompetes native cottonwoods. Ecol. Appl. 2012, 22, 532–549. [Google Scholar] [CrossRef]
- Hultine, K.R.; Bean, D.W.; Dudley, T.L.; Gehring, C.A. Species introductions and their cascading impacts on biotic interactions in desert riparian ecosystems. Integr. Comp. Biol. 2015, 55, 587–601. [Google Scholar] [CrossRef]
- Frew, A.; Powell, J.R.; Allsopp, P.G.; Sallam, N.; Johnson, S.N. Arbuscular mycorrhizal fungi promote silicon accumulation in plant roots, reducing the impacts of root herbivory. Plant Soil 2017, 419, 423–433. [Google Scholar] [CrossRef]
- Schwartz, M.W.; Hoeksema, J.D.; Gehring, C.A.; Johnson, N.C.; Klironomos, J.N.; Abbott, L.K.; Pringle, A. The promise and the potential consequences of the global transport of mycorrhizal fungal inoculum. Ecol. Lett. 2006, 9, 501–515. [Google Scholar] [CrossRef] [PubMed]
- Richter, B.S.; Stutz, J.C. Mycorrhizal inoculation of big sacaton: Implications for grassland restoration of abandoned agricultural fields. Restor. Ecol. 2002, 10, 607–616. [Google Scholar] [CrossRef]
- Maltz, M.R.; Treseder, K.K. Sources of inocula influence mycorrhizal colonization of plants in restoration projects: A meta-analysis. Restor. Ecol. 2015, 23, 625–634. [Google Scholar] [CrossRef]
- White, J.A.; Tallaksen, J.; Charvat, I. The effects of arbuscular mycorrhizal fungal inoculation at a roadside prairie restoration site. Mycologia 2008, 100, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Abdala-Roberts, L.; Pratt, R.; Pratt, J.D.; Mooney, K.A. Traits underlying community consequences of plant intra-specific diversity. PLoS ONE 2017, 12, e0183493. [Google Scholar] [CrossRef] [PubMed]
- Koske, R.E.; Gemma, J.N. A modified procedure for staining roots to detect VA mycorrhizas. Mycol. Res. 1989, 92, 486–488. [Google Scholar] [CrossRef]
- McGonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Bruun Brockhoff, P.; Haubo Bojesen Christensen, R. lmerTest: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82. [Google Scholar] [CrossRef]
- Lenth, R.V. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef]
- Bay, R.F.; Sher, A. Success of active revegetation after Tamarix removal in riparian ecosystems of the Southwestern United States: A quantitative assessment of past restoration projects. Restor. Ecol. 2008, 16, 113–128. [Google Scholar] [CrossRef]
- Ellis, L.M.; Crawford, C.S.; Molles, M.C., Jr. Comparison of litter dynamics in native and exotic riparian vegetation along the Middle Rio Grande of central New Mexico, U.S.A. J. Arid Environ. 1998, 38, 283–296. [Google Scholar] [CrossRef]
- Shi, Z.; Zhang, L.; Feng, G.; Christie, P.; Tian, C.; Li, X. Diversity of arbuscular mycorrhizal fungi associated with desert ephemerals growing under and beyond the canopies of tamarisk shrubs. Chin. Sci. Bull. 2006, 51, 132–139. [Google Scholar] [CrossRef]
- Lehnhoff, E.A.; Rew, L.J.; Zabinski, C.A.; Menalled, F.D. Reduced impacts or a longer lag phase? Tamarix in the Northwestern U.S.A. Wetlands 2012, 32, 497–508. [Google Scholar] [CrossRef]
- Taniguchi, T.; Acharya, K.; Imada, S.; Iwanaga, F.; Yamanaka, N. Arbuscular mycorrhizal colonization of Tamarix ramosissima along a salinity gradient in the southwestern United States. Landsc. Ecol. Eng. 2015, 11, 221–225. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, Y.; Li, W. Arbuscular mycorrhizal fungi infection in desert riparian forest and its environmental implications: A case study in the lower reach of Tarim River. Prog. Nat. Sci. 2008, 18, 983–991. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murray, L.; Schutte, B.J.; Ganguli, A.C.; Lehnhoff, E.A. Impacts of Tamarix (L.) Litter and Mycorrhizal Amendments on Baccharis salicifolia (Ruiz & Pav.) Pers. Competitiveness and Mycorrhizal Colonization. Agronomy 2019, 9, 453. https://doi.org/10.3390/agronomy9080453
Murray L, Schutte BJ, Ganguli AC, Lehnhoff EA. Impacts of Tamarix (L.) Litter and Mycorrhizal Amendments on Baccharis salicifolia (Ruiz & Pav.) Pers. Competitiveness and Mycorrhizal Colonization. Agronomy. 2019; 9(8):453. https://doi.org/10.3390/agronomy9080453
Chicago/Turabian StyleMurray, Leeland, Brian J. Schutte, Amy C. Ganguli, and Erik A. Lehnhoff. 2019. "Impacts of Tamarix (L.) Litter and Mycorrhizal Amendments on Baccharis salicifolia (Ruiz & Pav.) Pers. Competitiveness and Mycorrhizal Colonization" Agronomy 9, no. 8: 453. https://doi.org/10.3390/agronomy9080453