1. Introduction
Biotic stress remains one of the greatest challenges in agriculture today. It is a so-called complex trait, meaning that it is conferred by a large network of interacting genes. Plant pathogens, such as fungi, can be divided into necrotrophs, biotrophs, and hemi-biothrophs. Biotrophic pathogens exploit living host cells to continue their life cycle, whereas necrotrophs kill the cells of their hosts to feed on the cellular nutrients. However, most of the plant pathogens are hemi-biotrophic, including both a biotrophic and a necrotrophic phase in their life cycle, often in the early and late phase, respectively [
1,
2].
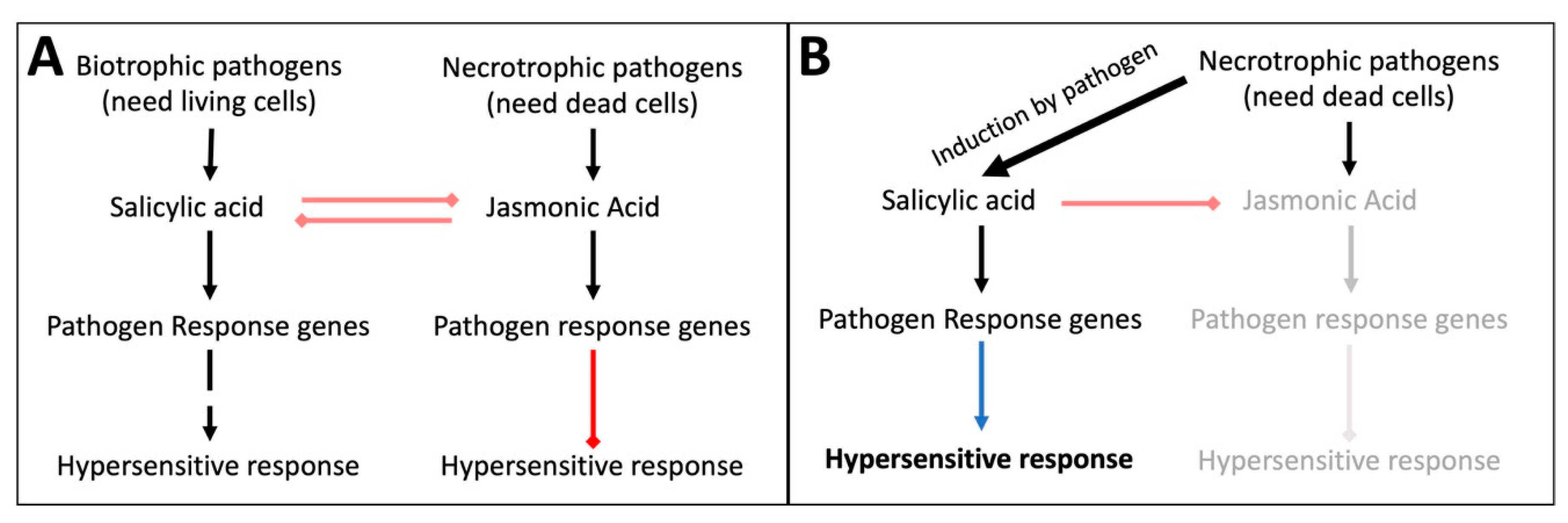
Plant innate immunity to fungi has two major pathways, the salicylic acid (SA) and the jasmonic acid (JA) pathway, induced by biotrophs and necrotrophs respectively [
3,
4]. Biotrophic fungi need alive host cells to feed on. When infected, the host plant activates the SA pathway, leading to the expression of specific pathogen response genes and, eventually, to hypersensitive response (HR) and local apoptosis that prevent the biotrophic fungus from spreading (
Figure 1A). The central player in the SA pathway is NPR1 [
5] that physically interacts with the kinases NIMIN1 and NIMIN2 [
4,
6] and induces transcription of transcription factors, such as WRKY70 [
6,
7]. The final step is the activation of pathogenesis-related (PR) genes, such as
PR1 and
PR2 [
8,
9] (
Figure 2). All the genes involved are only transiently expressed, because they put a great stress on the plant. Necrotrophic pathogens will activate the JA pathway, triggering the expression of pathway-specific pathogen response genes, such as
PR12 [
10,
11,
12,
13]) that lead to the suppression of HR [
2] (
Figure 1A).
Whereas the pathways have some mildly overlapping components, they generally repress each other to facilitate more rigid responses [
7,
14]. Some necrotrophic pathogens exploit the mutual exclusivity of these pathways by active induction of the SA pathway [
15,
16]. Thereby, they suppress the pathway that is supposed to fight them and kill host cells via HR to feed on (
Figure 1B). These fungi, such as
Botrytis cinerea and
Sclerotinia sclerotiorum, pose a significant threat to agriculture, for instance, in canola (
Brassica napus). During the infection process by these necrotrophic fungi, reactive oxygen species (ROS) are produced by the plant at the infection site. The ROS contribute to the host cell death, and as a consequence facilitate the infection and further growth of the fungi. By reducing the ROS production, the infection by necrotrophic pathogens can be inhibited [
17,
18,
19]. Rossi et al. [
19] increased the resistance of tobacco plants against the necrotrophic fungus
Botrytis cinerea by overexpressing the chloroplast-targeted antioxidant protein flavodoxin. Here we describe an alternative method to combat necrotrophic fungi in
Brassica napus by selecting plants with a reduced reaction to SA. In this manner, the plants become less sensitive to their own SA pathway.
To this end, we employed epigenetic rather than mutagenic selection, as previously established [
20,
21,
22,
23]. In this selection method, isogenic lines are used as starting material. From a population of about 200 plants, individual plants are selected for a specific trait as reduced respiration. The self-fertilized progenies of these plants are submitted to the same selection. These steps are repeated three times with the progenies obtained in the previous selection cycle. In this way epilines are obtained with distinct transcriptome, methylome and histone modifications. In rice epilines selected for higher energy use efficiency, the differentially expressed genes could be correlated with differential methylation and histone H3 lysine-4 trimethylation (H3K4me3) patterns [
23]. In canola epilines selected for drought tolerance, the drought-responsive genes were specifically differentially expressed and had a H3K4me3 pattern facilitating the transcription of the drought-responsive genes [
22]. In the three published cases, the selected properties remained stable over several generations: at least eight generations in the canola epilines [
20,
22] and four generations in the rice epilines [
23]. Considering that defense response networks are very interconnected, mutations often are either too disruptive or are compensated by the network. An epigenetic approach is more holistic and allows the discovery of new involved genes. Additionally, this selection method can be utilized on very young seedlings, eliminating the need for a lengthy growth process.
3. Results
3.1. Repeated Selection Leads to Canola Lines with Higher Tolerance to SA
To select lines tolerant to SA, a repeated selection approach was used as described [
20,
21,
22,
23]. When 12-day-old seedlings of canola are treated with 50 mg/L SA by watering, approximately 16 h after treatment the fresh weight of the cotyledons drops by approximately 15% (
Figure 3A), mainly due to water loss (lower turgor of cotyledons). To increase tolerance to SA seedlings that after SA treatment have cotyledons with a high fresh weight were selected. As a second parameter ascorbate concentration in the cotyledons was used as a measure for vitality. Ascorbate is abundantly present in leaves and has diverse biological roles in the live cycle of a plant: plant growth and development, photosynthesis, senescence, protectant against both abiotic and biotic stresses, … [
25,
26]. Although it was noticed that the ascorbate concentration of cotyledons increases when treated with SA (
Figure 3B), probably as a defense to oxidative stress induced by SA, seedlings were selected with a high ascorbate concentration as it was observed that seedlings with a low ascorbate concentration have a weak performance and had a very low seed set in the greenhouse (
Figure S3).
A population of approximately 200 canola seedlings was treated with SA by watering. Approximately 16 h after treatment, the seedlings were selected with a high cotyledon weight and a high ascorbate concentration in the cotyledons and falling above the 90% prediction interval of the regression line of weight versus ascorbate concentration (
Figure S1). These seedlings were transferred to the greenhouse for seed production by self-fertilization. The progenies of the first generation were subjected to the same selection, focusing on high cotyledon weight and ascorbate concentration, and the selected seedlings were transferred to the greenhouse for seed production. The selections and seed productions were done recurrently for three generations (
Figure S2). Finally, eight selected lines were maintained of which the ascorbate concentration and weight after SA treatment were similar to those of the untreated control line. Two lines derived from two different plants from the original population were chosen for in-depth analysis. Seed upscalings were done from 50 plants for each selected line and for the original line from which the selection had been started. Selected lines are indicated further by ‘Line 1’ and ‘Line 2’, and the original line is indicated as ‘control’.
3.2. SA Treatment Induces Water Loss in Canola Seedlings
Twelve-day-old seedlings of the control and the selected Lines 1 and 2 were treated with 0 and 50 mg/L SA and both the ascorbate concentration and fresh weight of the cotyledons were measured. The results of this experiment are summarized in
Table 1 and
Table 2, showing respectively the weights and the ascorbate concentrations of the cotyledons of the same seedlings. The percent weight loss of the cotyledons by the SA treatment was significant in the control (minus 13.3%), whereas the weight losses in both Line 1 and Line 2 were lower and not significant (minus 8.6% and minus 9.2%, respectively) (
Table 1). The ascorbate concentration per unit fresh weight had increased significantly in the control (12.3%) while the increase was less in the selected lines (6.3%) (
Table 2). The increased ascorbate concentration in the control can, at least partly, be explained by the enhanced water loss.
3.3. The SA-Tolerant Lines are More Tolerant to BOTRYTIS Cinerea
The sensitivity to Botrytis cinerea was evaluated on plants at different developmental stage and by different methods, more specifically by either spraying 4-week-old plants as a qualitative assay or by spotting a high density of spores on both cotyledons of 12-day-old seedlings as a quantitative assay.
In the spray assay the leaf damage by necrosis and chlorosis was scored by quantifying the non-green array versus the total area of the leaf. The control line infected by spraying with
Botrytis cinerea showed approximately 65% leaf chlorosis four days after treatment (
Figure 4C), indicative of heavy infection. In Line 1, this leaf chlorosis was reduced to 20% (
Figure 4A), whereas in Line 2 it was by 45% (in between control and Line 1) (
Figure 4B).
In spot assays on cotyledons, the lesion diameter was compared to that of the control. Line 1 was found to be significantly more tolerant to
Botrytis cinerea, but not Line 2 that showed an even slightly increased susceptibility to
Botrytis cinerea when compared to the control line (
Figure 4D).
3.4. Seed Yield of the Selected Lines are Similar to Those of the Control
Tolerance to fungi may have a negative impact on yield, especially when the defense mechanism is constitutive or when the induced level is high or maintained for a long period. The selected lines and control were evaluated in field trials in Canada on three locations with four repetitions per line and per location for one year. Diseases were controlled by standard agronomic practices. The seed yield data showed no yield drag in the selected lines (
Figure S4).
3.5. Known Genes in the SA Pathway Exhibit Changed Expression Levels
To understand the mechanism of the SA-tolerance, the expression of the genes of the SA pathway were studied in the control and the selected Lines 1 and 2, after induction by SA. The genes of the SA pathway are expressed transiently and at different time points after induction by SA, for this the transcription of the genes of the SA pathway were studied in time series after induction with SA for 0, 2, 3, 6, 9, 24, and 48 h. The genes studied were
NIMIN1,
NIMIN2,
WRKY70,
PR1, and
PR2. Because
NPR1 is regulated post-translationally [
4,
27] the transcription of NPR1 was not studied. When the transient expression of the SA response genes was checked by means of RT-qPCR, a strong transient induction was found in all tested previously reported genes, namely
PR1,
PR2,
WRKY70,
NIMIN1, and
NIMIN2 that all reached a maximum after six to nine hours post induction (
Figure 5). This expression was mostly lost within 48 h (
Figure 5). In the selected populations, the genes were still induced, but at a much lower rate, with over 10-fold reduction in relative expression increase, indicating the holistic suppression without the loss of the pathway.
3.6. The Gene Expression with Unknown Connection to the SA Pathway Is Modified
ChIP-Seq experiments with an antibody against polymerase II (Pol2) were performed on uninduced and SA-induced seedlings. The cotyledons of the induced seedlings were harvested six hours after induction. 1713 genes were induced by the SA treatment, while 1123 genes were repressed. From the 1713 induced genes 1070 could be annotated and from the 1123 repressed genes 697 could be annotated.
Figure S5 shows a summary of the gene ontology analyses. A set of differentially expressed genes previously unknown to be regulated directly or indirectly by SA were tested for their transient induction with RT-qPCR. Six new target genes were identified with altered transcription (
Figure 6).
CDPK5 (CALMODULIN-DOMAIN PROTEIN KINASE 5) has a calmodulin-dependent protein kinase activity and is associated with the abscisic acid (ABA) pathway. The control showed an increased expression 6–9 h post induction, whereas no such an expression occurred in the selected lines.
LRL12(LEAF RUST 10 DISEASE-RESISTANCE LOCUS RECEPTOR-LIKE PROTEIN KINASE-like 1.2) is a serine/threonine kinase reportedly involved in cellular response to ABA stimulus, cellular response to water deprivation, photoperiodism and flowering. Whereas the expression was strong in the control 6 and 24 h post induction, the response in selected lines was barely visible.
WAKLG (Wall-associated receptor kinase-like 8) again displayed a strong increase in transcription approximately 9 h post induction, but the SA-tolerant line showed no significant response.
GAUTE (Probable galacturonosyltransferase 14) is a transferase associated with carbohydrate synthesis and pollen development. In the control the expression increased in two waves: a fast response 2 h post induction and a second approximately 24 h post induction with SA. The SA-tolerant line had a similar pattern, but greatly diminished, at the limit of detection.
TOM1 (AT4G21790) encodes a host factor that is required for TMV virus multiplication. The transcription was overall higher in the selected lines than that in the control.
M4E813 is a predicted protein with possible association to stress response. Its expression strongly increased 9–24 h post induction with SA, whereas the SA-tolerant lines did exhibit such an induction.
3.7. Changes in H3K4me3
To connect the altered expression profiles of the SA-induced genes in the selected lines to a possible epigenetic marker correlated with activation of transcription, the H3K4me3 histone modification was studied in the control and the selected Lines 1 and 2.
After induction by SA the overall H3K4me3 changes were subtler than the Pol2 distributions, often showing a mark elongation from the transcriptional start to the entire gene. Only in genes high in the cascade, such as
WRKY70 and
NPR3, the H3K4me3 deposition increased 6 h post induction, but in genes at the end of the induction cascade, such as
PR1, the epigenetic mark did not change. Interestingly, the amounts of H3K4me3 deposition were lower in the selected Line 1 than those in the control (
Figure 7). This selected Line 1 showed also a greater decrease in transcription and had a significantly higher tolerance to
Botrytis cinerea than the control.
4. Discussion
The pathogen response network in plants is very complex and contains redundancies to compensate for inactivation by the pathogen interaction. The epigenetic selection method as used here was not only very rapid and limited to three selection cycles on very young plants, it also holistically shifted the entire SA response network without complete disruption.
As selection is based on a reduced reaction to SA treatments, the genes involved in the pathway are expected to be less activated than in the control. This effect occurred and was consistent over several SA pathway genes (
Figure 5). Over 10-fold less induction was observed in the selected lines.
Variation in PR gene expression between individuals, especially in the early response, is unsurprising. The PR genes are located at the end of a long signaling cascade. Mild fluctuations in expression timing can have a large influence on the transcription level when sampled at a particular time point. Indeed, the variation seen (6 h post induction;
Figure 5) is mostly due to one biological replicate.
Additionally, RNA-seq was used to identify genes, that after induction by SA, show the same abolished induction pattern as the known genes of the SA pathway. Two genes associated with the ABA pathway and dehydration were found. The ABA pathway has been reported to have some connections to the SA pathway [
28,
29]. Interestingly, one gene was identified that is involved in pollen development (
GAUTE). This gene has a unique expression pattern, but its induction was clearly reduced in the selected lines. Only one predicted gene activated by SA stimulus (
M4E813) has been computer annotated and, although it has been associated with a general response to stress (
UniProt: M4E813_BRARP), including both biotic and abiotic stresses (GO:0006950), structural annotations range from the Usp domain-containing (
InterPro: M4E813_BRARP) to the U-box proteins (
Genebank XM_013841965.2). All the newly identified genes could be further investigated for possible involvement in SA signaling.
In the list of genes induced by SA generated by the Pol2 ChIP-Seq, a GO enrichment for receptor binding and stress response was found. Interestingly within the suppressed genes the energy pathways are enriched, possibly indicating cellular prioritizing of pathogen defense over metabolism (
Figure S5). Additionally, stress response genes are enriched in repression, which hints at a mutual repression between pathways.
The selection method starting from an isogenic doubled haploid population and the altered H3K4me3 pattern in the selected lines demonstrate that the tolerance to SA is regulated at the epigenetic level. The epigenetic regulation had also been detected for H3K4me3 in the drought-tolerant canola lines [
22] and the methylome in the energy use efficient rice lines [
23], that had been selected in a manner similar to that used here to select the SA tolerance.
Although Lines 1 and 2 are selected likewise, Line 1 is more tolerant to Botrytis cinerea than Line 2 that shows only some tolerance in the spray assay. This result is in agreement with the more pronounced altered expression patterns of the SA-induced genes after SA treatment in Line 1. Also, after SA induction, the H3K4me3 deposition in Line 1 is lower than that in Line 2, which is similar to that in the control.
The use of epigenetics in agriculture is still in a developmental phase. During the life cycle of the crop, epigenetics is important for adaptation to the environment and strategies are being generated to use epigenetics to improve plant performance in the field [
30]. Recently, Raju et al. [
31] have demonstrated the potential utility of epilines in breeding. They had selected transgene-null epilines in soybean with the MSH1 system. The progenies obtained by crossing the epilines to the wild type had improved yield traits, namely in numbers of pods per plant, seed weight and seed yield.
Keeping in mind that the SA tolerance has an epigenetic origin, the question remains how stable the phenotype is over multiple generations. A high epigenetic stability is observed when plants are vegetatively propagated [
32]. Hauben et al. [
20] described the epigenetic stability in canola of energy use efficiency over eight generations of self-fertilizations. Raju et al. [
31] reported a loss of the improved yield traits after four generations of self-fertilization in the F2 lines that had been obtained by crossing transgene-null MSH1-derived epilines to the wild type. Here, only two generations were studied after selection without loss of the SA tolerance in the two generations obtained by self-fertilization under non-selective conditions. By means of an analogous selection scheme for other traits in rice and canola, we observed a low to very high stability, depending on the selected trait. For the SA tolerance more generations made by both self-fertilizations and crossings have to be examined to get some insight into the stability of the SA tolerance.
It can be argued that the SA tolerant lines are more susceptible to biotrophic pathogens that require the SA pathway for efficient resistance. However, the SA-tolerant lines were produced to combat the specific threat of necrotrophic fungi, such as Botrytis cinerea. Further experiments are needed to assay the sensitivity of the SA-tolerant lines to biotrophic pathogens and to other necrotrophic pathogens such as Sclerotinia sclerotiorum and Leptospharia maculans which are currently a large agronomic concern.
We demonstrated that epigenetic selection methods based on performance after a mimicked pathogen attack can produce plants with reduced reaction to that perceived threat. These lines show an altered expression in pathway-specific genes without complete break-down of the pathway. In laboratory and greenhouse tests, these lines had an increased tolerance to Botrytis cinerea, but further field trials have to evaluate the performance of the SA-tolerant lines under pathogen pressure.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}