Root Distribution and Its Impacts on the Drought Tolerance Capacity of Hybrid Rice in the Sichuan Basin Area of China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Experimental Design
2.2. Samples and Measurements
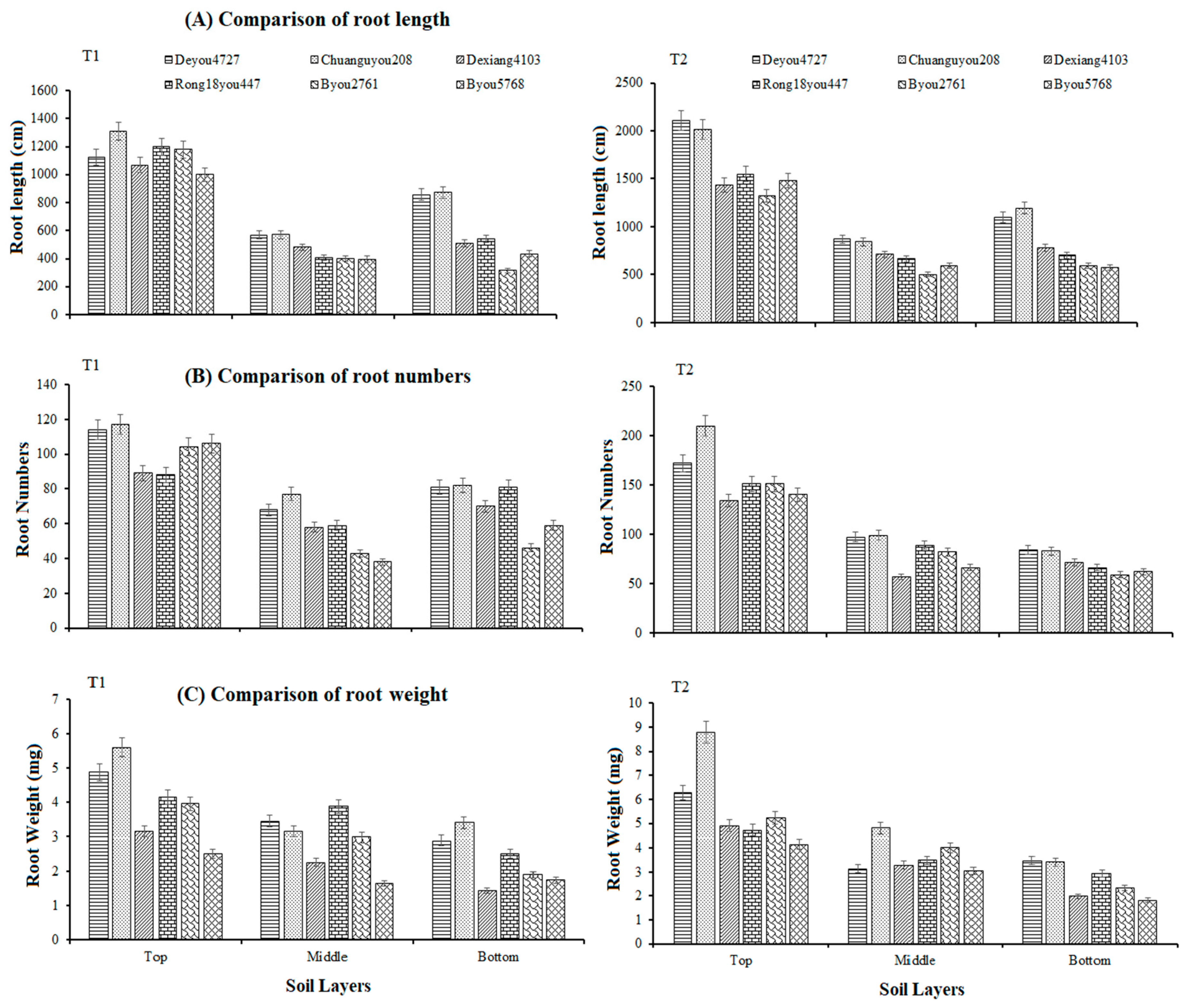
2.2.1. Root Distribution
2.2.2. Enzymes in Roots and Leaves
2.2.3. DEEPER ROOTING 1 (DRO1) Gene Sequencing
2.3. Statistical Methods
2.3.1. Method to Calculate the Drought Tolerance Capacity of Rice
2.3.2. Methods for Data Analysis
3. Results
3.1. Complex Tolerance Index of Rice under Drought Environment (CTIRDE)
3.2. Root Distribution Analysis
3.3. DEEPER ROOTING 1 (DRO1) Gene Sequence Analysis
3.4. Antioxidant Enzyme Activity under Drought Stress
3.5. Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, J.; Zhang, S.; Cheng, M.; Jiang, H.; Zhang, X.; Peng, C.; Lu, X.; Zhang, M.; Jin, J. Effect of drought on agronomic traits of rice and wheat: A meta-analysis. Int. J. Environ. Res. Public Health 2018, 15, 839. [Google Scholar] [CrossRef] [PubMed]
- Jing, R. Advances of research on drought resistance and water use efficiency in crop plants. Rev. China Agric. Sci. Technol. 2007, 9, 1–5. [Google Scholar]
- Reynolds, T.W.; Waddington, S.R.; Anderson, C.L.; Chew, A.; True, Z.; Cullen, A. Environmental impacts and constraints associated with the production of major food crops in Sub-Saharan Africa and South Asia. Food Secur. 2015. [Google Scholar] [CrossRef]
- Ndjiondjop, M.N.; Wambugu, P.W.; Sangare, J.R.; Gnikoua, K. The effects of drought on rice cultivation in sub-Saharan Africa and its mitigation: A review. Afr. J. Agric. Res. 2018, 13, 1257–1271. [Google Scholar] [CrossRef]
- Singh, S.; Prasad, S.; Yadav, V.; Kumar, A.; Jaiswal, B. Effect of Drought Stress on Yield and Yield Components of Rice (Oryza sativa L.) Genotypes. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 2752–2759. [Google Scholar]
- Singh, B.; Reddy, K.R.; Redoña, E.D.; Walker, T. Screening of Rice Cultivars for Morpho-Physiological Responses to Early-Season Soil Moisture Stress. Rice Sci. 2017, 24, 322–335. [Google Scholar] [CrossRef]
- Swapna, S.; Shylaraj, K.S. Screening for Osmotic Stress Responses in Rice Varieties under Drought Condition. Rice Sci. 2017, 24, 253–263. [Google Scholar] [CrossRef]
- Farooq, M.; Kobayashi, N.; Ito, O.; Wahid, A.; Serraj, R. Broader leaves result in better performance of indica rice under drought stress. J. Plant Physiol. 2010, 167, 1066–1075. [Google Scholar] [CrossRef]
- Fukai, S.; Cooper, M. Development of drought-resistant cultivars using physiomorphological traits in rice. Field Crop. Res. 1995, 40, 67–86. [Google Scholar] [CrossRef]
- Yue, B.; Xue, W.; Xiong, L.; Yu, X.; Luo, L.; Cui, K.; Jin, D.; Xing, Y.; Zhang, Q. Genetic basis of drought resistance at reproductive stage in rice: Separation of drought tolerance from drought avoidance. Genetics 2006, 172, 1213–1228. [Google Scholar] [CrossRef]
- Kumar, R.; Sreenu, K.; Singh, N.; Jain, N.; Singh, N.K.; Rai, V. Effect of drought stress on contrasting cultivars of rice. Int. J. Trop. Agric. 2015, 33, 1559–1564. [Google Scholar]
- Lipiec, J.; Doussan, C.; Nosalewicz, A.; Kondracka, K. Effect of drought and heat stresses on plant growth and yield: A review. Int. Agrophys. 2013, 27, 463–477. [Google Scholar] [CrossRef]
- Gowda, V.R.P.; Henry, A.; Yamauchi, A.; Shashidhar, H.E.; Serraj, R. Root biology and genetic improvement for drought avoidance in rice. Field Crop. Res. 2011, 122, 1–13. [Google Scholar] [CrossRef]
- Uga, Y.; Okuno, K.; Yano, M. Dro1, a major QTL involved in deep rooting of rice under upland field conditions. J. Exp. Bot. 2011, 62, 2485–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukai, S.; Pantuwan, G.; Jongdee, B.; Cooper, M. Screening for drought resistance in rainfed lowland rice. Field Crop. Res. 1999, 64, 61–74. [Google Scholar] [CrossRef]
- Uga, Y.; Sugimoto, K.; Ogawa, S.; Rane, J.; Ishitani, M.; Hara, N.; Kitomi, Y.; Inukai, Y.; Ono, K.; Kanno, N.; et al. Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions. Nat. Genet. 2013, 45, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant. Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Chutipaijit, S. Changes in physiological and antioxidant activity of indica rice seedlings in response to mannitol-induced osmotic stress. Chil. J. Agric. Res. 2016, 76, 455–462. [Google Scholar] [CrossRef] [Green Version]
- Bian, S.M.; Jiang, Y.W. Reactive oxygen species, antioxidant enzyme activities and gene expression patterns in leaves and roots of Kentucky bluegrass in response to drought stress and recovery. Sci. Hortic. (Amsterdam). 2009, 120, 264–270. [Google Scholar] [CrossRef]
- Nikolaeva, M.K.; Maevskaya, S.N.; Shugaev, A.G.; Bukhov, N.G. Effect of drought on chlorophyll content and antioxidant enzyme activities in leaves of three wheat cultivars varying in productivity. Russ. J. Plant. Physiol. 2010, 57, 87–95. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide Dismutase: Improved Assays and an Assay Applicable to Acrylamide Gels1. Anal. Biochem. 1971, 287, 276–287. [Google Scholar] [CrossRef]
- Kar, M.; Mishra, D. Catalase, Peroxidase, and Polyphenoloxidase Activities during. PLANT Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Jiang, Y.; Ye, S.; Karim, A.; Ling, Z.; He, Y.; Yang, S.; Luo, K. PtrWRKY73, a salicylic acid-inducible poplar WRKY transcription factor, is involved in disease resistance in Arabidopsis thaliana. Plant. Cell Rep. 2015, 34, 831–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nahar, S.; Kalita, J.; Sahoo, L.; Tanti, B. Morphophysiological and molecular effects of drought stress in rice. Ann. Plant. Sci. 2016, 5, 1409–1416. [Google Scholar] [CrossRef] [Green Version]
- Lou, Q.; Chen, L.; Mei, H.; Wei, H.; Feng, F.; Wang, P.; Xia, H.; Li, T.; Luo, L. Quantitative trait locus mapping of deep rooting by linkage and association analysis in rice. J. Exp. Bot. 2015, 66, 4749–4757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, Y.; Abe, J.; Kamoshita, A.; Yamagishi, J. Genotypic variation in root growth angle in rice (Oryza sativa L.) and its association with deep root development in upland fields with different water regimes. Plant. Soil 2006, 287, 117–129. [Google Scholar] [CrossRef]
- Asch, F.; Michael, D.; Abdoulaye, S.; Alain, A. Drought-induced changes in rooting patterns and assimilate partitioning between root and shoot in upland rice Drought-induced changes in rooting patterns and assimilate partitioning between root and shoot in upland rice. Field Crop. Res. 2005, 93, 223–236. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Somasundaram, R.; Panneerselvam, R. Drought Stress in Plants: A Review on Morphological Characteristics and Pigments Composition. Int. J. Agric. 2009, 11, 100–105. [Google Scholar]
- Allah, A.; Badawy, S.A.; Zayed, B.A.; Gohary, A.A. El The Role of Root System Traits in the Drought Tolerance of Rice (Oryza sativa L.). World Acad. Sci. Eng. Technol. 2010, 68, 1378–1382. [Google Scholar]
- Courtois, B.; Ahmadi, N.; Khowaja, F.; Price, A.H.; Rami, J.; Frouin, J.; Hamelin, C.; Ruiz, M. Rice Root Genetic Architecture: Meta-analysis from a Drought QTL Database. Rice 2009, 2, 115–128. [Google Scholar] [CrossRef] [Green Version]
- Uga, Y.; Yamamoto, E.; Kanno, N.; Kawai, S.; Mizubayashi, T.; Fukuoka, S. A major QTL controlling deep rooting on rice chromosome 4. Sci. Rep. 2013. [Google Scholar] [CrossRef]
- Uga, Y.; Kitomi, Y.; Yamamoto, E.; Kanno, N.; Kawai, S.; Mizubayashi, T.; Fukuoka, S. A QTL for root growth angle on rice chromosome 7 is involved in the genetic pathway of DEEPER ROOTING 1. Rice 2015. [Google Scholar] [CrossRef]
- Kitomi, Y.; Kanno, N.; Kawai, S.; Mizubayashi, T.; Fukuoka, S.; Uga, Y. QTLs underlying natural variation of root growth angle among rice cultivars with the same functional allele of DEEPER ROOTING 1. Rice 2015. [Google Scholar] [CrossRef] [PubMed]
- Nahar, S.; Sahoo, L.; Tanti, B. Screening of drought tolerant rice through morpho-physiological and biochemical approaches. Biocatal. Agric. Biotechnol. 2018, 15, 150–159. [Google Scholar] [CrossRef]
- Sgherri, C.L.M.; Pinzino, C.; Navari-Lzzo, F. Sunflower seedlings subjected to increasing stress by water deficit: Changes in O2~ production related to the composition of thylakoid membranes. Physiol. Plant. 1996. [Google Scholar] [CrossRef]
- Smirnoff, N. The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol. 1993. [Google Scholar] [CrossRef]
- Luna, C.M.; Pastori, G.M.; Driscoll, S.; Groten, K.; Bernard, S.; Foyer, C.H. Drought controls on H2O2accumulation, catalase (CAT) activity and CAT gene expression in wheat. J. Exp. Bot. 2005, 56, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-ríos, L.; Pinto, M. Effect of salt stress on antioxidant enzymes and lipid peroxidation in leaves in two contrasting corn, ‘Lluteño’ and ‘Jubilee’. Chil. J. Agric. Res. 2014, 74, 89–95. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Number | Genotype | Male × Female of F1 Hybrid | Number | Genotype | Male × Female of F1 Hybrid |
|---|---|---|---|---|---|
| 1 | Yixiang4245 | Yixiang1A × Yihui4245 | 24 | Tianlongyou1340 | Tianlong13A × Tianlonghui140 |
| 2 | Jiayou727 | Jialing1A × Chenghui727 | 25 | Tianlongyou140 | TianlongS × Tianlonghui140 |
| 3 | Chuannongyou498 | Chuannong1A × Shuhui498 | 26 | Tianlongyou872 | Tianlong8A × Tianlonghui72 |
| 4 | Deyou4923 | Dexiang074A × R4923 | 27 | jingyou781 | Jing7A × Dehui381 |
| 5 | Rong18you307 | Rong18A × Ronghui307 | 28 | Tianlongyou540 | Long5A × Tianlonghui140 |
| 6 | Dexiang4103 | Dexiang074A × LuhuiH103 | 29 | Byou5761 | B501A × Xikehui 4761 |
| 7 | Huaxiany357 | HuaxiangyouA × Mianhui357 | 30 | Longyou450 | Longxiang4A × Tianlonghui1250 |
| 8 | Guangyou2928 | Guangkang13A × xikehui2928 | 31 | Chuanyou5778 | Chuan106A × Nanhui5778 |
| 9 | Nei6you611 | Neixiang6A × Mianhui138 | 32 | Byou2727 | B213A × Chenghui727 |
| 10 | Chuanyou5727 | Chuan345A × Chenghui727 | 33 | Zhufengyou4720 | Zhufeng4A × Xikehui720 |
| 11 | Rong18you447 | Rong18A × Ronghui447 | 34 | Byou3446 | B395A × Xikehui3446 |
| 12 | Yixiangyou1108 | Yixiang1A × Yihui1108 | 35 | Chuanyou1288 | Chuan106A × Xikehui1288 |
| 13 | Fyou498 | FS3A × Shuhui498 | 36 | Jinyou1720 | Jin1A × Xikehui720 |
| 14 | Deyou4727 | Dexiang074A × Shuhui727 | 37 | Byou4446 | B4A × Xikehui3446 |
| 15 | Luyou257 | Lu98A × Shuhui257 | 38 | Rongyou3268 | Rong3A × Xikehui1288 |
| 16 | Chuanguyou208 | ChuanguA × Shuhui208 | 39 | Byou727 | B1A × Chenghui727 |
| 17 | IIyou498 | II-32A × Shuhui498 | 40 | Guangyou3446 | Guangkang13A × xikehui3446 |
| 18 | Nei6you538 | Neixiang6A × Shuhui538 | 41 | Byou2761 | B2A × Xikehui4761 |
| 19 | Chuannongyou298 | Chuannong2A × Shuhui498 | 42 | Byou2909 | B2A × Xikehui9909 |
| 20 | Chuannongyouhuazhan | Chuannong1A × Huazhan | 43 | Byou1288 | B1A × Xikehui1288 |
| 21 | Chuannongyou3203 | Neixiang6A × Luhui9 | 44 | Byou4928 | B4A × Xikehui2928 |
| 22 | Nei5you317 | Neixiang5A × Neihui3317 | 45 | Byou5768 | B501A × Xikehui768 |
| 23 | Neixiang7you317 | Neixiang7A × Neihui3317 | 46 | Gangyou725 | Gang46A × Mianhui725 |
| Materials | 5′UTR | 5′UTR | EXON 3 | EXON3 | EXON4 | EXON4 | 3′UTR | Intron5 |
|---|---|---|---|---|---|---|---|---|
| cDNA | 27 | 150 | 617 | 752 | 943 | 962 | 1133–1134 | -- |
| gDNA | 27 | 150 | 2096 | 2231 | 2513 | 2532 | 2909–2910 | 2815 |
| Seq (+) | A/G | C/T | T/C | A/C | A/- | C/A | CT/-- | A/T |
| NPB | A | C | T | A | A | C | CT | A |
| Chuanguyou208 | A | T | C | C | A | C | -- | A |
| Deyou4727 | A | T | C | C | A | C | -- | A |
| Dexiang4103 | H | T | C | H | A | C | -- | A |
| Rong18you447 | A | T | C | H | A | C | -- | H |
| Byou2761 | G | T | C | A | A | C | -- | A |
| Byou5768 | H | T | C | A | A | C | -- | A |
| Rice Varieties | SOD | CAT | POD | |||
|---|---|---|---|---|---|---|
| Leaf | Root | Leaf | Root | Leaf | Root | |
| Deyou 4727 | 69.46 c | 84.62 a | 342.12 ab | 494.62 c | 7.90 a | 32.02 a |
| Chuanguyou 208 | 85.44 ab | 87.51 a | 412.08 a | 815.18 a | 10.48 a | 22.13 b |
| Byou2761 | 53.43 d | 72.70 b | 304.13 b | 389.92 d | 7.75 a | 17.12 c |
| Byou5768 | 84.93 ab | 75.59 b | 175.06 c | 438.12 cd | 6.89 a | 19.44 c |
| Dexiang4103 | 81.04 b | 60.50 c | 242.17 bc | 449.56 cd | 5.79 a | 11.41 d |
| Rong18you447 | 91.54 a | 61.01 c | 207.39 c | 656.78 b | 7.98 a | 10.88 d |
| Trait. | Top Soil | Middle Soil | Bottom Soil | Antioxidant Enzymes | Leaf | Root |
|---|---|---|---|---|---|---|
| Root length | −0.114 | 0.936 ** | 0.956 ** | SOD | −0.338 | 0.983 ** |
| Root number | 0.369 | 0.96 ** | 0.874 * | CAT | 0.668 | 0.242 |
| Root weight | 0.767 * | 0.511 | 0.748 * | POD | 0.648 | 0.890 * |
| Trait | Top Soil | Middle Soil | Bottom Soil | Antioxidant Enzymes | Leaf | Root |
|---|---|---|---|---|---|---|
| Root length | 0.517 | 0.714 * | 0.857 * | SOD | 0.338 | 0.852 * |
| Root number | 0.266 | 0.496 | 0.825 * | CAT | 0.215 | 0.242 |
| Root weight | 0.309 | 0.684 | 0.774 * | POD | 0.548 | 0.763 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Samo, N.; Li, L.; Wang, M.; Qadir, M.; Jiang, K.; Qin, J.; Rasul, F.; Yang, G.; Hu, Y. Root Distribution and Its Impacts on the Drought Tolerance Capacity of Hybrid Rice in the Sichuan Basin Area of China. Agronomy 2019, 9, 79. https://doi.org/10.3390/agronomy9020079
Wang X, Samo N, Li L, Wang M, Qadir M, Jiang K, Qin J, Rasul F, Yang G, Hu Y. Root Distribution and Its Impacts on the Drought Tolerance Capacity of Hybrid Rice in the Sichuan Basin Area of China. Agronomy. 2019; 9(2):79. https://doi.org/10.3390/agronomy9020079
Chicago/Turabian StyleWang, Xuechun, Naseem Samo, Lamei Li, Mengran Wang, Muslim Qadir, Kaifeng Jiang, Jian Qin, Fahd Rasul, Guotao Yang, and Yungao Hu. 2019. "Root Distribution and Its Impacts on the Drought Tolerance Capacity of Hybrid Rice in the Sichuan Basin Area of China" Agronomy 9, no. 2: 79. https://doi.org/10.3390/agronomy9020079