3.1. Eastern Region Before the 2000s
The data indicated that biotypes E3, E6 and E7, from R3Y, had low and medium levels of resistance to propanil, compared with an intermediate susceptibility biotype (LCh02). Their RF values were statistically significant but lower than 10 (
Table 2). The GR
50 values of these biotypes were higher than (E7) or close to the label rate (E3 and E6). However, the GR
90 values were higher than the label rate for the three biotypes, especially in E7, at almost three times that amount. Conversely, the LCh01 biotype was susceptible, showing an RF of 1, which is not significant, and also a GR
50 and GR
90 much lower than the label rate (
Table 2). LCh01 susceptibility attracted our attention because it came from a field in which a rice–soybean-alternating rotation was in place from 1979 to 1988, followed by five years of continuous rice (R5Y). During this 10-year period, trifluralin alone or triflurain and imazaquin tank-mixes were used as preemergence in a non-glyphosate-resistant soybean crop. Possibly, this management contributed to lessening the selection pressure on the
E. crus-galli biotype, due to the fact that these herbicides present different modes of action than those used in rice.
With the range of doses used for quinclorac evaluation (
Table 2), it was not possible to estimate GR
50 for the susceptible biotype (LM04), and consequently, RF could not be estimated. However, it was observed that GR
50 values for biotype E3, E6, E7 and LCh01 were higher than the label rate (375 g ha
−1) and even higher than the maximum dose evaluated (3000 g ha
−1), especially in E3, E7 and LCh01. The same pattern was determined for GR
90, showing that quinclorac resistance had evolved. The biotype E5 was also quinclorac-resistant; in spite of GR
50 and GR
90 being below label rate, the coefficient for the “c” parameter was not near zero on the
x-axis. The four biotypes coming from the R3Y (E3, E5, E6 and E7) resulted in resistance to quinclorac, and also the one collected from R5Y (LCh01). The selection process in those cases occurred under just two sprayings of quinclorac tank-mixed with propanil in R3Y, and with at least three sprayings in R5Y. At that time, herbicide applications used to be performed with high propanil doses (2400 to 3360 g ai ha
−1) and quinclorac (320 g ai ha
−1) over
E. crus-galli populations with a high proportion of four-leaf plants, or even with tillers. Besides, water flushing after two weeks of herbicide application was usual, and the flooding date was established at around 50 to 60 days after seeding, on average. Under this management scheme, we can hypothesize that quinclorac was the herbicide that withstood the major selection pressure.
Concerning clomazone evaluation (
Table 2), all biotypes seemed to be susceptible, showing non-significant RF values (E3, E6, E7 and ZA03). For the LCh01 biotype, no model was adjusted, because it was very sensitive to the lowest dose used, and the ZA03 biotype needed very little clomazone to reach GR
50. Most of the GR
90 values were lower than the label rate, although E7 showed a slightly greater value. None of these
E. crus-galli biotypes had been exposed to clomazone before.
In summary, the E7 biotype showed a medium resistance level to propanil and a high resistance level to quinclorac, while E3 and E6 biotypes had a low resistance level to propanil and a high resistance level to quinclorac. Besides this, LCh01 and E5 biotypes showed a high level of resistance only to quinclorac. Similar results were reported by other authors [
28,
29,
30], in situations where propanil or quinclorac were used without rotation, in either crops or herbicides (modes of action).
3.2. Eastern Collection, 2006-2010
Twenty-one biotypes were evaluated primarily with propanil, quinclorac and clomazone. For propanil (
Table 3), statistically significant RF values ranged from 0.5 to 4.9, related to the GR
50 of 896 g ha
−1 corresponding to the susceptible biotype, LCh02. Some differences were observed in the degree of resistance between biotypes. For example, CL44, LM01 and RTb01, with RF values between 3 and 5, reached GR
50 and GR
90 values higher than the propanil label rate (1920 g ha
−1), denoting a noteworthy resistance level. Meanwhile, other biotypes such as CB02, RB281, RB282, ZA01, showed RF values below 1 and GR
50 values lower than the label rate, but GR
90 values higher than the label rate. The rest of the biotypes were susceptible, presenting GR
50 and GR
90 values lower than the label rate of propanil. The screening reflects differences in the evolutionary process, probably caused by variances in selection pressure, mainly due to rice being cropped many seasons in the same paddock, at times when few chemical options were available.
Data for the quinclorac dose–response assay (
Table 4) showed that it was not possible to estimate GR
50 values for the susceptible biotype (LM04) and two more, because no model was adjusted with the dose range utilized. All the other biotypes reached a GR
50 value higher than the label rate (375 g ha
−1), indicating resistance. Even though no model could be fitted for RB281, it turned out to be quinclorac-resistant, which is explained by the fact that its fresh weight decrease was not achived with 3000 g ha
−1 of quinclorac. In the second study carried out for quinclorac, with a more ample dose range, the GR
50 values for RB281 and ZA01 were much higher than the rate of quinclorac (375 g ha
−1) and their RF values were significant, while for E7 biotype, the RF was not significant (
Table 5). However, the “c” coefficient remained high for the E7 biotype and was not tangent to zero on the
x-axis, indicating that fresh weight was not depleted in the dose range used, confirming its high level of resistance.
For clomazone, RF values were not significant; thus, none of the evaluated biotypes showed resistance to this herbicide (
Table 6). Nevertheless, some biotypes such as LCh02, LM01 and LM03 showed a slightly greater GR
90 than the label rate (480 g ha
−1), perhaps indicating a selection process was occuring. During this period, most of the clomazone was used in a tank-mix with propanil and quinclorac, on early or late postemergence applications. In the last decade, clomazone use has changed, and more the 40% of rice acreage has begun to be treated with glyphosate and clomazone tank-mixed in preemergence [
31], which could explain why clomazone is still highly effective in
E. crus-galli control.
From this set of biotypes evaluated, LM01 and RTb01 presented a medium level of resistance to propanil and a high level of resistance to quinclorac, while RB281, RB282 and ZA01 presented a high level of resistance to quinclorac. Finally, biotype CL44 only showed medium-level resistance to propanil. The most used herbicide combination between 1995 and 2010 was a tank-mix with propanil + quinclorac, and plus clomazone (since 1995). Irrigation management in early years was not appropriately carried out, meaning that herbicide applications and the water lamina were concreted 45 to 60 days after seeding, with overgrown weeds. In that scenario, we hypothesize that selection pressure leaned more towards quinclorac, facilitating the evolution of resistance in many situations. In the last years of the mentioned period, early postemergence applications covered more acreage, and water management was also improved. In addition to this, more molecules appeared, enabling farmers to diversify in terms of chemical control.
Some of the above evaluated biotypes were also tested with acetolactate synthase-inhibitor herbicides (ALS, Group B-HRAC), along with some collections which originated from side-by-side fields with variable CL history (E0CL, E1CL, E2CL and E3CL). Seventeen biotypes were assessed, and only one showed resistance to IMIS (
Table 7). Biotype E0CL was considered the susceptible one for IMIS, although a model could not be adjusted. It was never previously exposed to those herbicides, and it did not produce any biomass with a small dose as 12.25 g ha
−1 of imazapyr + imazapic. On the other hand, the GR
50 and GR
90 values of E3CL were two and six times higher than the label rate, respectively. This biotype (E3CL) originated from a rice field that was cropped with CL rice three years in a row, and four IMIS applications were performed in total.
In the evaluation of bispyribac–sodium (
Table 8) and penoxsulam (
Table 9), all biotypes were susceptible, with the lowest dose enough to control
E. crus-galli. Therefore, no cross-resistance was detected to bispypribac–sodium or penoxsulam and IMIS in biotypes coming from CL fields. The average use of those herbicides before 2010 was not above 20% of the total area, mainly including bispyribac and tank-mixes with other modes of action. Therefore, it was expected not to find biotypes with resistance in such situations.
Obtained results are in accordance with what took place in other regions, where the high reliance on any molecule induces the selection of resistant biotypes [
13,
15,
17,
32,
33].
3.4. Northern Region, Since 2014
In the case of propanil, one biotype (TSF7) from the 12 tested had a significant RF of 6, although its GR
90 was below the label rate, possibly denoting a low level of resistance. Other biotypes reached an RF of about 6 to 7, but this was not significant. The TVA8 biotype had a GR
90 higher than the label dose (
Table 12), indicating an early alert sign that could be evolving resistance. Although propanil is the oldest rice herbicide in use in Uruguay, its use has significantly declined in the last decade, due to the availability of new molecules. Furthermore, it has been used mostly in tank-mixes with other modes of action, contributing to delaying the evolutionary process.
For quinclorac, the first series of assays yielded 100% resistance, evidenced by the fact that all biotypes were not controlled by this herbicide. Since there was no good reference for a susceptible biotype for this herbicide in the collection, a biotype from the eastern reference (LM04) was used. Resistance factors from 10 to more than 10 thousand were determined (
Table 13), and in all cases, the effective dose to control 90% of the biotype (GR
90) was much higher than the herbicide label rate. The biotype TSF7 was the exception because it showed no significant RF; nevertheless, eight times the label rate did not reduce its biomass production much. In some biotypes, such as TAP4, TEV9 and ARI11, even eight times the label rate did not reach any biomass reduction; therefore, no parameters could be calculated (GR
50 or RF). Such results are in accordance with the extended history of the widespread use of this herbicide in rice systems, alone or in tank-mixes with propanil and clomazone, among others. Quinclorac has been used with continuous increases of its recommended dose from 320 to 375 g ai ha
−1, to 450 to 500 g ai ha
−1 in the last 20 years, and complaints about its failures are usual among farmers.
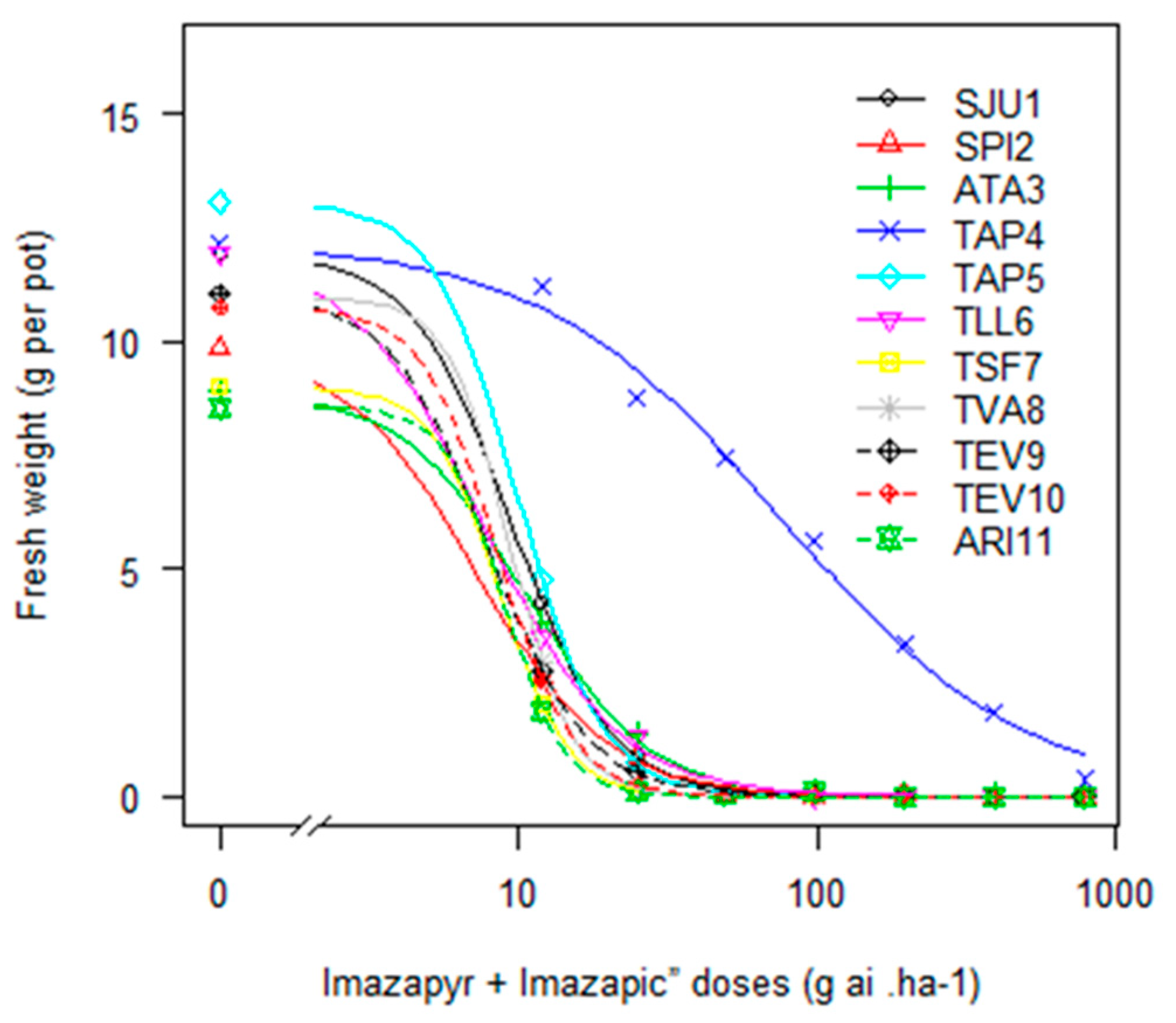
In the study carried out with IMIS, one of the 12 biotypes (TAP4) evidenced resistance (
Figure 1). This biotype originated from a CL rice system, presenting a significant RF of 11 ± 2.5, compared to the susceptible biotype (SPI2). The effective dose to control 90% of TAP4 biomass was six times greater than the label recommended rate, denoting the complicated situation for a farmer to deal with this problem. Furthermore, the preliminary results of response curves carried out with recently collected biotypes (four from eight) from CL systems show alarming levels of
E. crus-galli IMIS resistance (with GR
50 values five to 50 times the label recommended rate of 98 g ai ha
−1 (data not shown)).
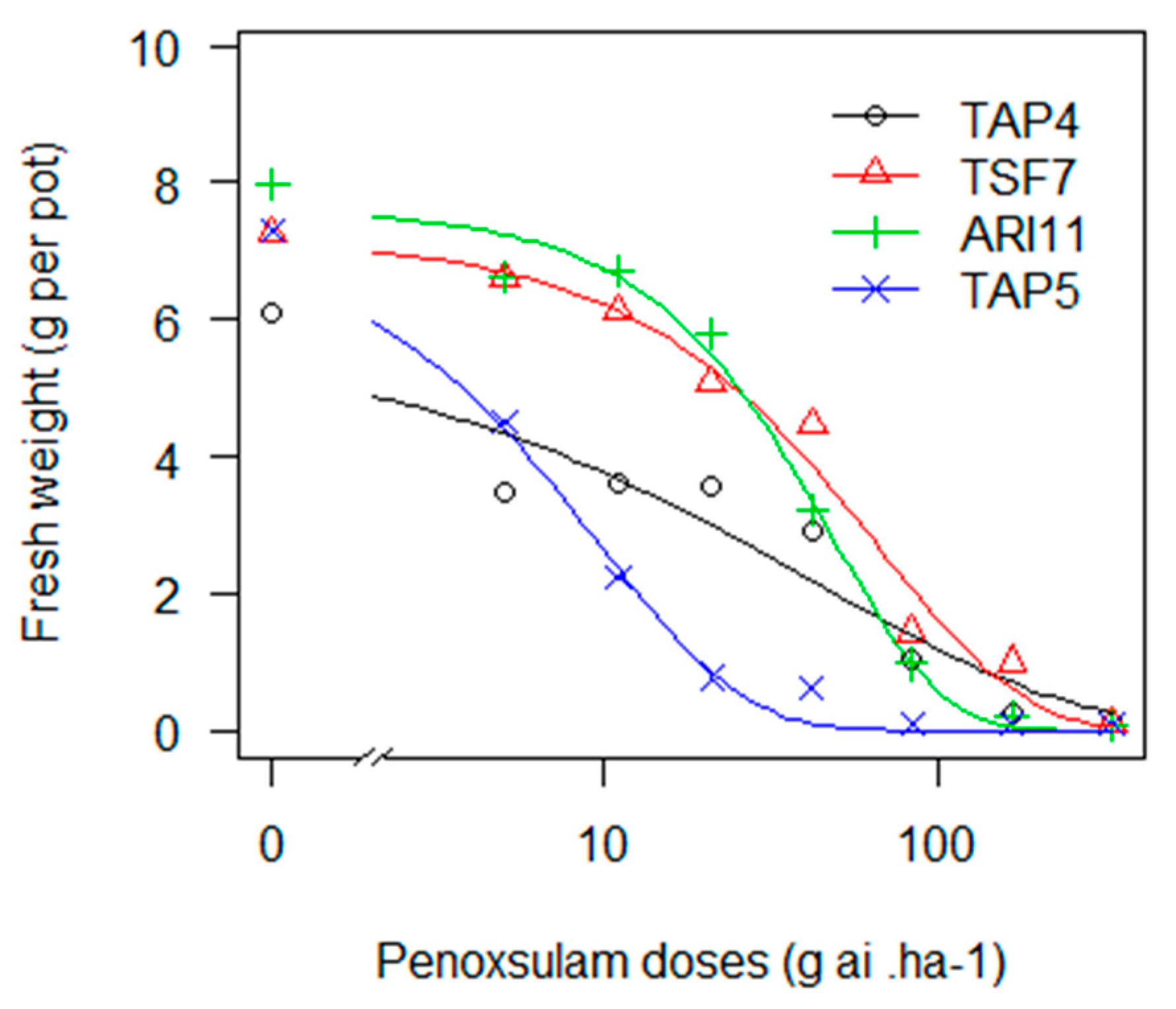
Despite most of the 12 biotypes exhibiting a low GR
50 with penoxsulam, TAP4, TSF7 and ARI11 showed a different behaviour (
Figure 2). Intermediate and significant RF values (3, 7 and 6, respectively) when compared to the susceptible biotype (TAP5) and a higher GR
90 than the label dose (
Table 14) may indicate that these biotypes have become resistant to this herbicide. Penoxsulam was introduced in 2004 and has become a relevant molecule in the last decade. In the early years, it was applied alone as a very highly efficient herbicide, but it endured high selection pressure. Moreover, as with IMIS, more emphasis should be placed on the higher risk resulting from the overuse of an ALS herbicide [
10]. Most recently, its use has become frequently in mixes with other modes of action.
Biotype TAP4, in summary, showed a wide-ranging ability to survive quinclorac, IMIS, and penoxsulam. This behaviour, characteristic of a multiple resistant biotype, could suggest an eventual metabolic resistance mechanism [
34,
35]. Such studies are underway, as well as the identification of target site mutations in the ALS enzyme in IMIS and penoxsulam-resistant biotypes, alongside the mechanism involved in quinclorac resistance [
36,
37,
38,
39].
Lastly, it is worth mentioning that, in this collection, fortunately, no resistance was found to have evolved against bispyribac–sodium and for both acetyl-CoA carboxylase-inhibitors (ACCase), profoxidim and cyhalofop (data not shown). The use of bispyribac has been reduced to 10% of the total area, mainly in mixes with other modes of action, while cyhalofop has increased its presence to 10%, with profoxidim used in a very small proportion.
3.5. Implications of Herbicide-Resistant E. crus-galli for Rice Production in Uruguay
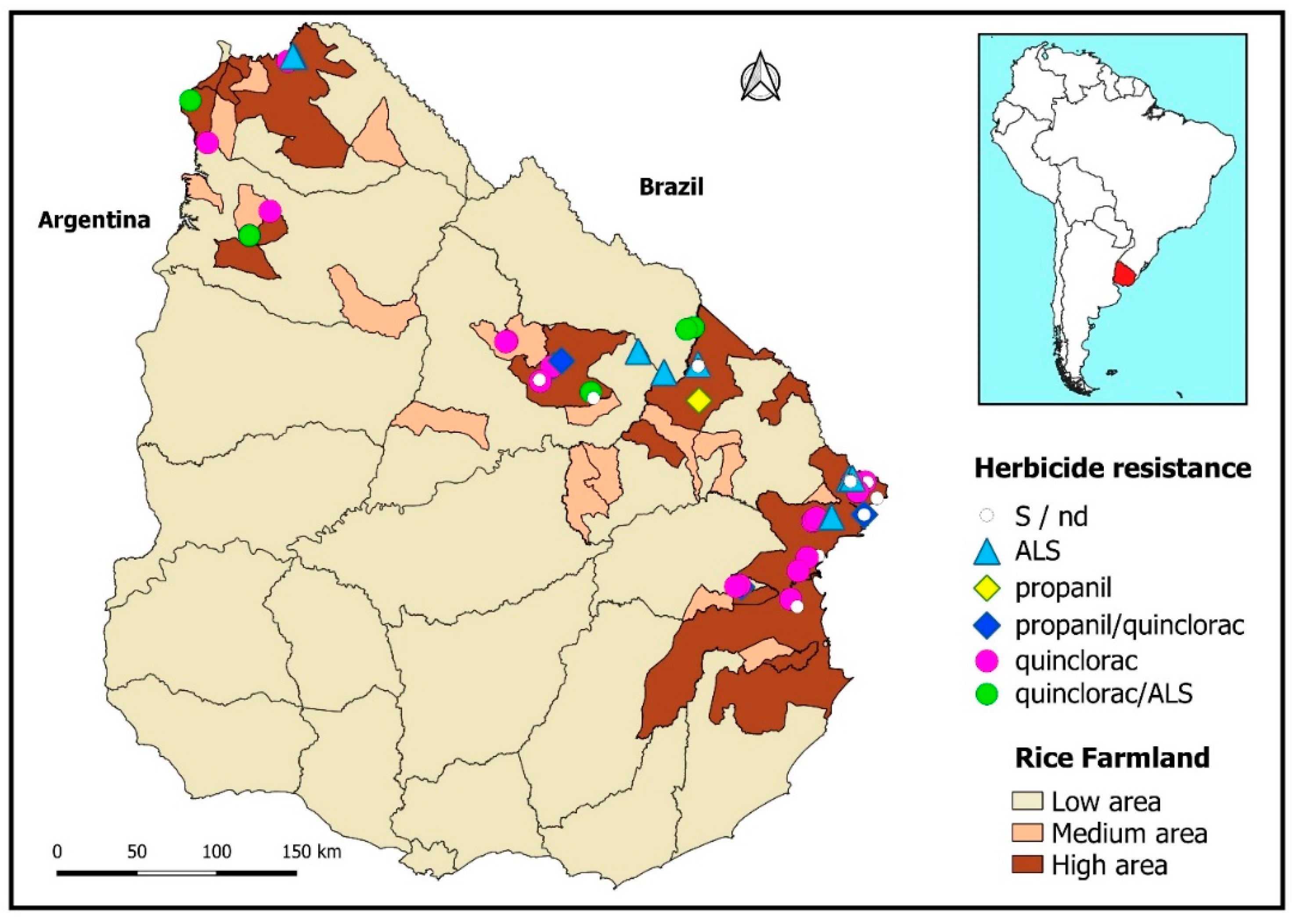
A synopsis of the different biotypes’ resistance statuses can be visualized as disseminated throughout the territory, along with the densest rice cropping areas (
Figure 3).
This situation denotes a serious problem for rice farmers because their main weed control management tool is at risk. Not only do they have reduced herbicide options, but also insufficient control leads to low yields and grain quality losses, increased production costs and farmland depreciation. Undoubtedly, a fundamental route to slowing down herbicide resistance is the application of integrated management practices [
13,
23,
40] in which chemical and cultural controls complement each other. These practices could contribute to reducing weed prevalence and dissemination, facilitating the crops’ competition ability or implying a change in the environmental conditions for weed growth through the reduction of their survival rate. Examples of these management actions are the reduction of weed seed entry to the soil seed bank (survivors from herbicide applications) to avoid an in-field increase of them and the use of clean machinery, especially the harvester, to reduce the dispersion of problematic weed biotypes, or the contamination from field to field.
In addition, the use of rotations with other crops could facilitate the use of different combinations of herbicide modes of actions than the ones currently used in rice and with perennial pastures that could break the weeds’ life cycles. This effort should be a collaborative compromise between farmers, researchers, industry and policymakers if the project is to succeed in maintaining sustainable commercial production systems. Moreover, and considering the dimension of the problem, our findings may be of assistance to further research elucidating the underlying resistance mechanism/s. Such information should be the base for developing management strategies to retard the evolution processes. Our paradigm should be to determine how it is possible to maintain both the productivity and sustainability of these highly productive rice systems.



{kind=link}
{kind=link}
{kind=link}


