Antioxidant Seasonal Changes in Soilless Greenhouse Sweet Peppers



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Agronomic Features
2.2. Greenhouse Facilities and Climatic Data
2.3. Fruit Quality Measurements
2.4. Statistical Analysis
3. Results
3.1. Greenhouse Microclimate and External Climatic Data
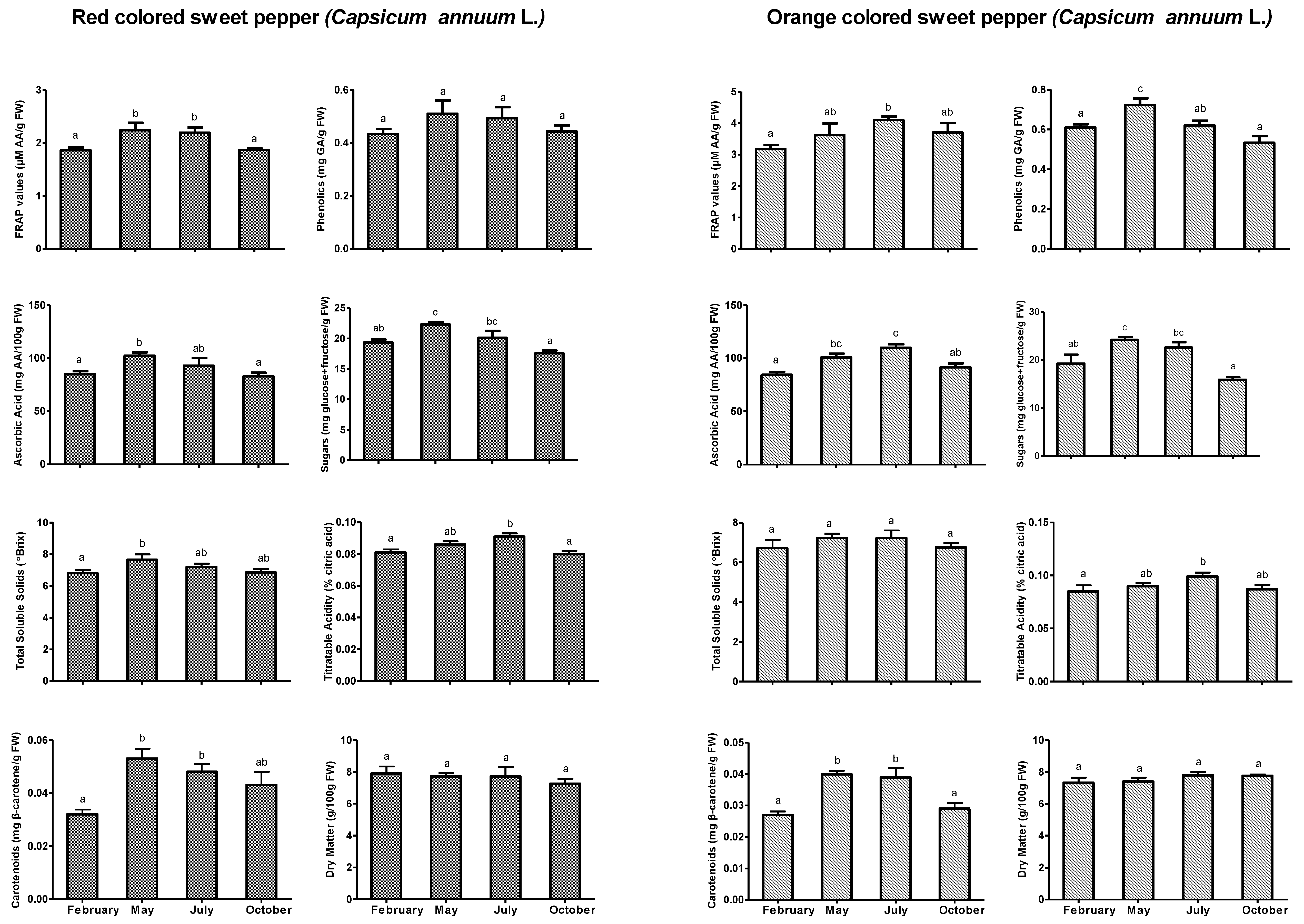
3.2. Antioxidants and Other Fruit Quality and Yield Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- USDA, United States Department of Agriculture, Economic Research Service. Vegetables and Pulses Yearbook Tables. Available online: https://www.ers.usda.gov/data-products/vegetables-and-pulses-data/ (accessed on 10 June 2019).
- Savvas, D.; Gianquinto, G.P.; Tuzel, Y.; Gruda, N. Soilless Culture. In Good Agricultural Practices for Greenhouse Vegetable Crops. Principles for Mediterranean Climate Areas; FAO, Ed.; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013; pp. 303–354. [Google Scholar]
- Gruda, N. Do soilless culture systems have an influence on product quality of vegetables? J. Appl. Bot. Food 2009, 82, 511–519. [Google Scholar]
- Gruda, N.; Qaryouti, M.M.; Leonardi, C. Growing Media. In Good Agricultural Practices for Greenhouse Vegetable Crops. Principles for Mediterranean Climate Areas; FAO, Ed.; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013; pp. 301–371. [Google Scholar]
- Gruda, N. Increasing Sustainability of Growing Media Constituents and Stand-Alone Subsstrates in Soilless Culture Systems. Agronomy 2019, 9, 298. [Google Scholar] [CrossRef]
- Gázquez, J.C.; López, J.C.; Pérez-Parra, J.J.; Baeza, E.J.; Saéz, M.; Parra, A. Greenhouse Cooling Strategies for Mediterranean Climate Areas. Acta Hortic. 2008, 801, 425–432. [Google Scholar] [CrossRef]
- EUR-Lex. Access to European Union Law, Commission Implementing Regulation (EU) No 543/2011. Available online: https://eur-lex.europa.eu/legal-content/GA/TXT/?uri=CELEX:32011R0543 (accessed on 10 June 2019).
- Gruda, N. Impact of Environmental Factors on Product Quality of Greenhouse Vegetables for Fresh Consumption. Crit. Rev. Plant Sci. 2015, 24, 227–247. [Google Scholar] [CrossRef]
- Santin, M.; Lucini, L.; Castagna, A.; Rocchetti, G.; Hauser, M.-T.; Ranieri, A. Comparative “Phenol-Omics” and Gene Expression Analyses in Peach (Prunus Persica) Skin in Response to Different Postharvest UV-B Treatments. Plant Physiol. Biochem. 2019, 135, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Prakash, D.; Gupta, K.R. The Antioxidant Phytochemicals of Nutraceutical Importance. Open Nutraceuticals 2009, 2, 20–35. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant Polyphenols as Dietary Antioxidants in Human Health and Disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Sullivan, A.M.; O’Callaghan, Y.C.; O’Grady, M.N.; Queguineur, B.; Hanniffy, D.; Troy, D.J.; Kerry, J.P.; O’Brien, N.M. In Vitro and Cellular Antioxidant Activities of Seaweed Extracts Prepared from Five Brown Seaweeds Harvested in Spring from the West Coast of Ireland. Food Chem. 2011, 126, 1064–1070. [Google Scholar] [CrossRef]
- Pellegrini, N.; Serafini, M.; Colombi, B.; Del Rio, D.; Salvatore, S.; Bianchi, M.; Brighenti, F. Total Antioxidant Capacity of Plant Foods, Beverages and Oils Consumed in Italy Assessed by Three Different In Vitro Assays. J. Nutr. 2003, 133, 2812–2819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toor, R.K.; Savage, G.P.; Lister, C.E. Seasonal Variations in the Antioxidant Composition of Greenhouse Grown Tomatoes. J. Food Compos. Anal. 2006, 19, 1–10. [Google Scholar] [CrossRef]
- Li, Q.; Kubota, C. Effects of Supplemental Light Quality on Growth and Phytochemicals of Baby Leaf Lettuce. Environ. Exp. Bot. 2009, 67, 59–64. [Google Scholar] [CrossRef]
- Blokhina, O. Antioxidants, Oxidative Damage and Oxygen Deprivation Stress: A Review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Shan, B.; Cai, Y.Z.; Sun, M.; Corke, H. Antioxidant Capacity of 26 Spice Extracts and Characterization of Their Phenolic Constituents. J. Agric. Food Chem. 2005, 53, 7749–7759. [Google Scholar] [CrossRef] [PubMed]
- Conte, A.; Conversa, G.; Scrocco, C.; Brescia, I.; Laverse, J.; Elia, A.; Del Nobile, M.A. Influence of Growing Periods on the Quality of Baby Spinach Leaves at Harvest and during Storage as Minimally Processed Produce. Postharvest Biol. Technol. 2008, 50, 190–196. [Google Scholar] [CrossRef]
- Sun, J.; Chu, Y.-F.; Wu, X.; Liu, R.H. Antioxidant and Antiproliferative Activities of Common Fruits. J. Agric. Food Chem. 2002, 50, 7449–7454. [Google Scholar] [CrossRef] [PubMed]
- Grifoni, D.; Agati, G.; Bussotti, F.; Michelozzi, M.; Pollastrini, M.; Zipoli, G. Different Responses of Arbutus Unedo and Vitis Vinifera Leaves to UV Filtration and Subsequent Exposure to Solar Radiation. Environ. Exp. Bot. 2016, 128, 1–10. [Google Scholar] [CrossRef]
- Del-Castillo-Alonso, M.-Á.; Diago, M.P.; Tomás-Las-Heras, R.; Monforte, L.; Soriano, G.; Martínez-Abaigar, J.; Núñez-Olivera, E. Effects of Ambient Solar UV Radiation on Grapevine Leaf Physiology and Berry Phenolic Composition along One Entire Season under Mediterranean Field Conditions. Plant Physiol. Biochem. 2016, 109, 374–386. [Google Scholar] [CrossRef] [PubMed]
- Escobedo, J.F.; Gomes, E.N.; Oliveira, A.P.; Soares, J. Modeling Hourly and Daily Fractions of UV, PAR and NIR to Global Solar Radiation under Various Sky Conditions at Botucatu, Brazil. Appl. Energy 2009, 86, 299–309. [Google Scholar] [CrossRef]
- León-Chan, R.G.; López-Meyer, M.; Osuna-Enciso, T.; Sañudo-Barajas, J.A.; Heredia, J.B.; León-Félix, J. Low Temperature and Ultraviolet-B Radiation Affect Chlorophyll Content and Induce the Accumulation of UV-B-Absorbing and Antioxidant Compounds in Bell Pepper (Capsicum Annuum) Plants. Environ. Exp. Bot. 2017, 139, 143–151. [Google Scholar] [CrossRef]
- Dáder, B.; Gwynn-Jones, D.; Moreno, A.; Winters, A.; Fereres, A. Impact of UV-A Radiation on the Performance of Aphids and Whiteflies and on the Leaf Chemistry of Their Host Plants. J. Photochem. Photobiol. B Biol. 2014, 138, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Olympios, C.M. The Cultivation Technique of Vegetables in Greenhouses (In Greek); Stamoulis Publications: Athens, Greece, 2001. [Google Scholar]
- Katsoulas, N.; Kittas, C.; Dimokas, G.; Lykas, C.H. Effect of Irrigation Frequency on Rose Flower Production and Quality. Biosyst. Eng. 2006, 93, 237–244. [Google Scholar] [CrossRef]
- Jacovides, C.; Assimakopoulos, V.; Tymvios, F.; Theophilou, K.; Asimakopoulos, D. Solar Global UV (280–380 nm) Radiation and Its Relationship with Solar Global Radiation Measured on the Island of Cyprus. Energy J. 2006, 31, 2728–2738. [Google Scholar] [CrossRef]
- Scalbert, A.; Monties, B.; Janin, G. Tannins in Wood: Comparison of Different Estimation Methods. J. Agric. Food Chem. 1989, 37, 1324–1329. [Google Scholar] [CrossRef]
- Neocleous, D.; Ntatsi, G. Seasonal Variations of Antioxidants and Other Agronomic Features in Soilless Production of Selected Fresh Aromatic Herbs. Sci. Hortic. 2018, 234, 290–299. [Google Scholar] [CrossRef]
- Benzie, F.F.I.; Strain, J.J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurements of total antioxidant power and ascorbic acid concentration. Methods Enzymol. 1999, 299, 15–27. [Google Scholar] [PubMed]
- Benzie, F.F.I.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “Antioxidant Power”. The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Amaya, D.B. A Guide to Carotenoid Analysis; ILDI Press: Washington, DC, USA, 2001. [Google Scholar]
- Gastilla, N.; Baeza, E. Greenhouse site selection. In Good Agricultural Practices for Greenhouse Vegetable Crops. Principles for Mediterranean Climate Areas; FAO, Ed.; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013; pp. 21–33. [Google Scholar]
- Zimmer, A.R.; Leonardi, B.; Miron, D.; Schapoval, E.; de Oliveira, J.R.; Gosmann, G. Antioxidant and Anti-Inflammatory Properties of Capsicum Baccatum: From Traditional Use to Scientific Approach. J. Ethnopharmacol. 2012, 139, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Ghasemnezhad, M.; Sherafati, M.; Payvast, G.A. Variation in Phenolic Compounds, Ascorbic Acid and Antioxidant Activity of Five Coloured Bell Pepper (Capsicum Annum) Fruits at Two Different Harvest Times. J. Funct. Foods 2011, 3, 44–49. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 3rd ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2002. [Google Scholar]
- Perla, V.; Nimmakayala, P.; Nadimi, M.; Alaparthi, S.; Hankins, G.R.; Ebert, A.W.; Reddy, U.K. Vitamin C and Reducing Sugars in the World Collection of Capsicum Baccatum, L. Genotypes. Food Chem. 2016, 202, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Mateos, R.M.; Jiménez, A.; Román, P.; Romojaro, F.; Bacarizo, S.; Leterrier, M.; Gómez, M.; Sevilla, F.; del Río, L.A.; Corpas, F.J.; et al. Antioxidant Systems from Pepper (Capsicum annuum L.): Involvement in the Response to Temperature Changes in Ripe Fruits. Int. J. Mol. Sci. 2013, 14, 9556–9580. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Pugliese, A.; Bonesi, M.; Menichini, F.; Tundis, R. Evaluation of chemical profile and antioxidant activity of twenty cultivars from Capsicum annuum, Capsicum baccatum, Capsicum chacoense and Capsicum chinense: A comparison between fresh and processed peppers. LWT—Food Sci. Technol. 2015, 64, 623–631. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized Methods for the Determination of Antioxidant Capacity and Phenolics in Foods and Dietary Supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef] [PubMed]
- Kittas, C.; Tchamitchian, M.; Katsoulas, N.; Karaiskou, P.; Papaioannou, C.H. Effect of Two UV-Absorbing Greenhouse-Covering Films on Growth and Yield of an Eggplant Soilless Crop. Sci. Hortic. 2006, 110, 30–37. [Google Scholar] [CrossRef]
- Schnitzler, W.; Gruda, N. Hydroponics and Product Quality. In Hydroponic Production of Vegetables and Ornamentalss; Savvas, D., Passam, H., Eds.; Embryo Publications: Athens, Greece, 2002; pp. 373–411. ISBN 960-8002-12-5. [Google Scholar]
- Neugart, S.; Schreiner, M. UVB and UVA as eustressors in horticultural and agricultural crops. Sci. Hortic. 2018, 234, 370–381. [Google Scholar] [CrossRef]
- Gruda, N.; Savvas, D.; Colla, G.; Rouphael, Y. Impacts of Genetic Material and Current Technologies on Product Quality of Selected Greenhouse Vegetables—A Review. Eur. J. Hortic. Sci. 2018, 83, 319–328. [Google Scholar] [CrossRef]
- Shaw, N.L.; Cantliffe, D.J. Brightly colored pepper cultivars for greenhouse production in Florida. Proc. Fla. State Hort. Soc. 2002, 115, 236–241. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| J | F | M | A | M | J | J | A | S | O | N | D | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gh | 1075 (810) | 1509 (1022) | 1801 (1096) | 2269 (1234) | 2528 (1211) | 2838 (1135) | 2746 (1144) | 2515 (1148) | 2151 (1095) | 1681 (987) | 1251 (857) | 972 (751) |
| Guv | 32 (24) | 45 (30) | 60 (37) | 81 (44) | 91 (44) | 92 (37) | 86 (36) | 76 (34) | 89 (45) | 63 (37) | 44 (30) | 31 (24) |
| To | 14.8 (4.2) | 20.5 (1.7) | 22.6 (4.2) | 26.0 (2.2) | 28.6 (2.5) | 31.7 (2.1) | 33.5 (3.1) | 30.0 (2.9) | 28.5 (3.0) | 22.7 (3.4) | 20.5 (2.6) | 12 (3.2) |
| RHo | 55.7 (14.7) | 49.3 (12.5) | 46.8 (8.4) | 48.4 (5.9) | 53.5 (9.5) | 65.8 (8.5) | 63.9 (14.8) | 72.0 (11.5) | 55.4 (11.0) | 47.2 (15.0) | 49.2 (11.9) | 53.0 (9.8) |
| Ti | 21.0 (4.9) | 23.7 (2.4) | 24.5 (2.9) | 25.4 (3.5) | 26.2 (2.9) | 27.9 (2.0) | 28.0 (2.0) | 27.9 (2.0) | 24.2 (2.2) | 23.8 (4.7) | 23.0 (3.6) | 21.8 (2.3) |
| RHi | 69.8 (11.8) | 59.2 (12.5) | 60.2 (11.3) | 61.5 (7.8) | 67.2 (6.6) | 69.8 (5.3) | 65.3 (9.1) | 75.1 (14.3) | 60.3 (12.8) | 62.9 (14.7) | 54.9 (11.7) | 61.5 (6.3) |
| VPD | 0.9 (0.5) | 1.2 (0.4) | 1.1 (0.2) | 1.1 (0.3) | 1.1 (0.3) | 1.3 (0.4) | 1.4 (0.2) | 1.1 (0.3) | 1.2 (0.3) | 1.2 (0.2) | 1.3 (0.2) | 0.9 (0.3) |
| F Probability | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Source | FRAP | Ph | AA | Sug | TSS | TA | Car | Chl | DM |
| Season | 0.0010 | 0.6085 | <0.0001 | <0.0001 | 0.0012 | <0.0001 | <0.0001 | 0.1736 | 0.1724 |
| Cultivar | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| S × C | 0.0305 | 0.0042 | 0.0534 | 0.1100 | 0.6260 | 0.9321 | 0.0035 | <0.0001 | 0.0016 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neocleous, D.; Nikolaou, G. Antioxidant Seasonal Changes in Soilless Greenhouse Sweet Peppers. Agronomy 2019, 9, 730. https://doi.org/10.3390/agronomy9110730
Neocleous D, Nikolaou G. Antioxidant Seasonal Changes in Soilless Greenhouse Sweet Peppers. Agronomy. 2019; 9(11):730. https://doi.org/10.3390/agronomy9110730
Chicago/Turabian StyleNeocleous, Damianos, and Georgios Nikolaou. 2019. "Antioxidant Seasonal Changes in Soilless Greenhouse Sweet Peppers" Agronomy 9, no. 11: 730. https://doi.org/10.3390/agronomy9110730