Effect of Plant Growth-Promoting Rhizobacteria on Salicornia ramosissima Seed Germination under Salinity, CO2 and Temperature Stress

 , , , and
, , , and
Abstract
:1. Introduction
- (1)
- the isolation and characterization of the cultivable rhizobacteria of several halophytes from southwestern Spain salt marshes;
- (2)
- the selection of PGPR consortia;
- (3)
- the analysis of the impact of PGPR inoculants on seed germination of Salicornia ramosissima under NaCl, CO2 and temperature stressors.
2. Materials and Methods
2.1. Isolation of Cultivable Rhizobacteria From Halophytes
2.2. Rhizobacteria Characterization, Identification and Consortia Design
2.3. Rhizobacteria Used for Inoculation in This Study
2.4. Salicornia ramosissima Seeds Source and Experimental Treatments
2.5. Preparation of Bacterial Inoculants and Inoculation of Salicornia ramosissima Seeds
2.6. Salicornia ramosissima Gnobiotic Seed Germination Assay
2.7. Statistical Analysis
3. Results
3.1. Isolation, Characterization and Identification of Cultivable Bacteria from H. portulacoides and S. ramosissima Rhizosphere in Southwestern Spain Salt Marshes
3.2. Design of Two Rhizobacteria Consortia from H. portulacoides and S. ramosissima Cultivable Isolates
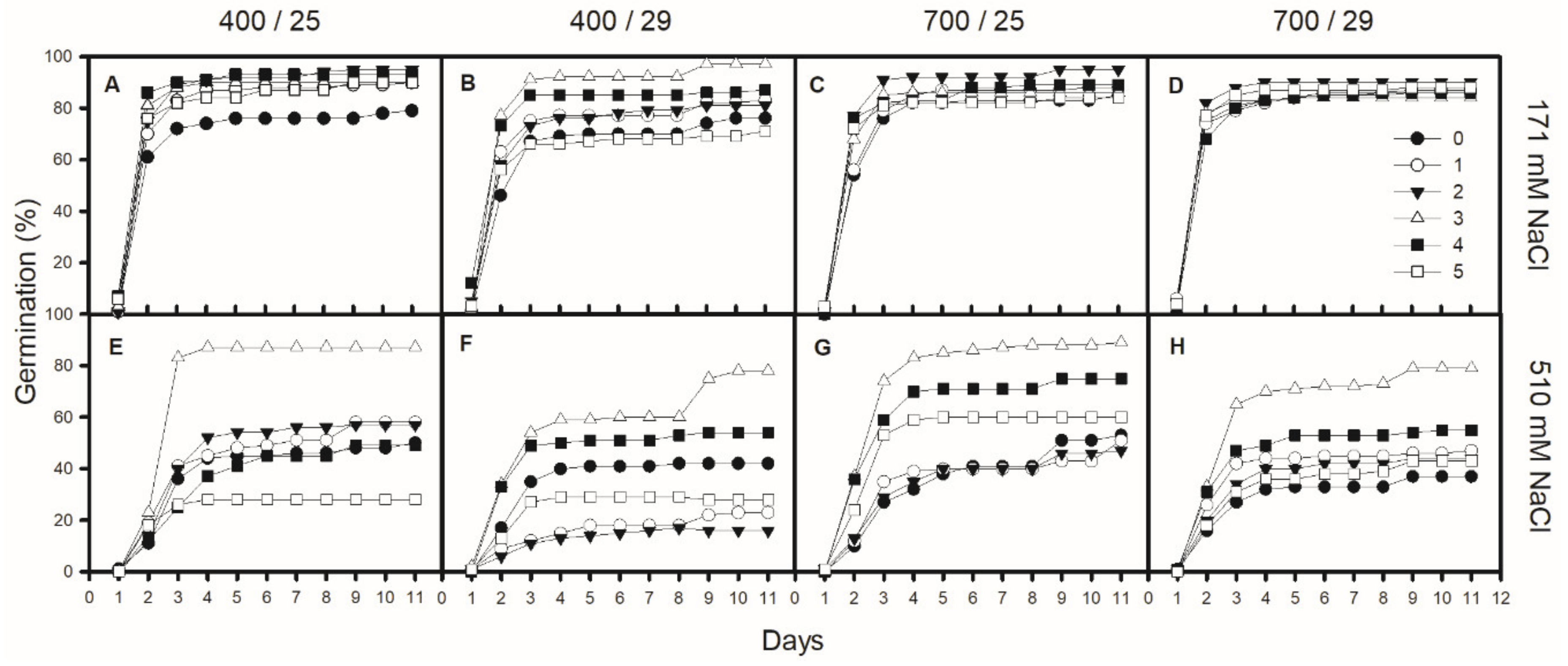
3.3. Effect of CO2, Temperature, NaCl and PGPR Inoculation on Salicornia ramosissima Seed Germination Parameters
3.4. Effect of CO2, Temperature, NaCl and PGPR Inoculation on Seedling Biomass
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014. [Google Scholar]
- Ventura, Y.; Eshel, A.; Pasternak, D.; Sagi, M. The development of halophyte-based agriculture: Past and present. Ann. Bot. 2015, 115, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Loconsole, D.; Cristiano, G.; De Lucia, B. Glassworts: From wild salt marsh species to sustainable edible crops. Agriculture 2019, 9, 14. [Google Scholar] [CrossRef]
- Ventura, Y.; Sagi, M. Halophyte crop cultivation: The case for Salicornia and Sarcocornia. Environ. Exp. Bot. 2013, 92, 144–153. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Buhmann, A.; Papenbrock, J. An economic point of view of secondary compounds in halophytes. Funct. Plant Biol. 2013, 40, 952–967. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Plant salt tolerance: Adaptations in halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef]
- Benjamin, J.J.; Lucini, L.; Jothiramshekar, S.; Parida, A. Metabolomic insights into the mechanisms underlying tolerance to salinity in different halophytes. Plant Physiol. Biochem. 2018, 135, 528–545. [Google Scholar] [CrossRef]
- Yensen, N.P. Halophyte uses for the twenty-first century. In Ecophysiology of High Salinity Tolerant Plants; Khan, M.A., Weber, D.J., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 367–397. ISBN 9781402040177. [Google Scholar]
- Grigore, M.; Oprica, L. Halophytes as Possible Source of Antioxidant Compounds, in a Scenario Based On Threatened Agriculture and Food Crisis. Iran. J. Public Health 2015, 44, 1153–1155. [Google Scholar]
- Petropoulos, S.A.; Karkanis, A.; Martins, N.; Ferreira, I.C.F.R. Edible halophytes of the Mediterranean basin: Potential candidates for novel food products. Trends Food Sci. Technol. 2018, 74, 69–84. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Sharma, S.; Shah, M.T. Successful Cultivation of Salicornia brachiata—A Sea Asparagus Utilizing RO Reject Water: A Sustainable Solution. Int. J. Waste Resour. 2018, 8, 322. [Google Scholar] [CrossRef]
- Lu, D.; Zhang, M.; Wang, S.; Cai, J.; Zhou, X.; Zhu, C. Nutritional characterization and changes in quality of Salicornia bigelovii Torr. during storage. LWT Food Sci. Technol. 2010, 43, 519–524. [Google Scholar] [CrossRef]
- Urbano, M.; Tomaselli, V.; Bisignano, V.; Veronico, G.; Hammer, K.; Laghetti, G. Salicornia patula Duval-Jouve: From gathering of wild plants to some attempts of cultivation in Apulia region (southern Italy). Genet. Resour. Crop Evol. 2017, 64, 1465–1472. [Google Scholar] [CrossRef]
- Patel, S. Salicornia: Evaluating the halophytic extremophile as a food and a pharmaceutical candidate. 3 Biotech 2016, 6, 104. [Google Scholar] [CrossRef] [PubMed]
- Barreira, L.; Resek, E.; Rodrigues, M.J.; Rocha, M.I.; Pereira, H.; Bandarra, N.; da Silva, M.M.; Varela, J.; Custódio, L. Halophytes: Gourmet food with nutritional health benefits? J. Food Compos. Anal. 2017, 59, 35–42. [Google Scholar] [CrossRef]
- Maciel, E.; Lillebø, A.; Domingues, P.; da Costa, E.; Calado, R.; Domingues, M.R.M. Polar lipidome profiling of Salicornia ramosissima and Halimione portulacoides and the relevance of lipidomics for the valorization of halophytes. Phytochemistry 2018, 153, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Plant Growth-Promoting Bacteria: Mechanisms and Applications. Scientifica (Cairo) 2012, 2012, 963401. [Google Scholar] [CrossRef]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef]
- Komaresofla, B.R.; Alikhani, H.A.; Etesami, H.; Khoshkholgh-Sima, N.A. Improved growth and salinity tolerance of the halophyte Salicornia sp. by co–inoculation with endophytic and rhizosphere bacteria. Appl. Soil Ecol. 2019, 138, 160–170. [Google Scholar] [CrossRef]
- Teixeira, C.; Almeida, C.M.R.; da Silva, M.N.; Bordalo, A.A.; Mucha, A.P. Development of autochthonous microbial consortia for enhanced phytoremediation of salt-marsh sediments contaminated with cadmium. Sci. Total Environ. 2014, 493, 757–765. [Google Scholar] [CrossRef]
- De Souza, M.P.; Huang, C.P.A.; Chee, N.; Terry, N. Rhizosphere bacteria enhance the accumulation of selenium and mercury in wetland plants. Planta 1999, 209, 259–263. [Google Scholar] [CrossRef]
- Szymańska, S.; Płociniczak, T.; Piotrowska-Seget, Z.; Hrynkiewicz, K. Endophytic and rhizosphere bacteria associated with the roots of the halophyte Salicornia europaea L.—Community structure and metabolic potential. Microbiol. Res. 2016, 192, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Bibi, F.; Strobel, G.A.; Naseer, M.I.; Yasir, M.; Al-Ghamdi, A.A.K.; Azhar, E.I. Microbial flora associated with the Halophyte-Salsola imbricate and its biotechnical potential. Front. Microbiol. 2018, 9, 65. [Google Scholar] [CrossRef]
- Navarro-Torre, S.; Mateos-Naranjo, E.; Caviedes, M.A.; Pajuelo, E.; Rodríguez-Llorente, I.D. Isolation of plant-growth-promoting and metal-resistant cultivable bacteria from Arthrocnemum macrostachyum in the Odiel marshes with potential use in phytoremediation. Mar. Pollut. Bull. 2016, 110, 133–142. [Google Scholar] [CrossRef]
- Yuan, Z.; Druzhinina, I.S.; Labbé, J.; Redman, R.; Qin, Y.; Rodriguez, R.; Zhang, C.; Tuskan, G.A.; Lin, F. Specialized Microbiome of a Halophyte and its Role in Helping Non-Host Plants to Withstand Salinity. Sci. Rep. 2016, 6, 32467. [Google Scholar] [CrossRef] [PubMed]
- Mesa, J.; Mateos-Naranjo, E.; Caviedes, M.A.; Redondo-Gómez, S.; Pajuelo, E.; Rodríguez-Llorente, I.D. Scouting contaminated estuaries: Heavy metal resistant and plant growth promoting rhizobacteria in the native metal rhizoaccumulator Spartina maritima. Mar. Pollut. Bull. 2015, 90, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Andrades-Moreno, L.; Del Castillo, I.; Parra, R.; Doukkali, B.; Redondo-Gómez, S.; Pérez-Palacios, P.; Caviedes, M.A.; Pajuelo, E.; Rodríguez-Llorente, I.D. Prospecting metal-resistant plant-growth promoting rhizobacteria for rhizoremediation of metal contaminated estuaries using Spartina densiflora. Environ. Sci. Pollut. Res. Int. 2014, 21, 3713–3721. [Google Scholar] [CrossRef]
- Paredes-Páliz, K.I.; Caviedes, M.A.; Doukkali, B.; Mateos-Naranjo, E.; Rodríguez-Llorente, I.D.; Pajuelo, E. Screening beneficial rhizobacteria from Spartina maritima for phytoremediation of metal polluted salt marshes: Comparison of gram-positive and gram-negative strains. Environ. Sci. Pollut. Res. 2016, 23, 19825–19837. [Google Scholar] [CrossRef]
- Mesa, J.; Mateos-naranjo, E.; Caviedes, M.A.; Redondo-gómez, S.; Pajuelo, E.; Rodríguez-llorente, I.D. Endophytic Cultivable Bacteria of the Metal Bioaccumulator Spartina maritima Improve Plant Growth but Not Metal Uptake in Polluted Marshes Soils. Front. Microbiol. 2015, 6, 1450. [Google Scholar] [CrossRef]
- Mesa, J.; Rodríguez-Llorente, I.D.; Pajuelo, E.; Piedras, J.M.B.; Caviedes, M.A.; Redondo-Gómez, S.; Mateos-Naranjo, E. Moving closer towards restoration of contaminated estuaries: Bioaugmentation with autochthonous rhizobacteria improves metal rhizoaccumulation in native Spartina maritima. J. Hazard. Mater. 2015, 300, 263–271. [Google Scholar] [CrossRef]
- Dobereiner, J. Isolation and identification of aerobic nitrogen-fixing bacteria from soil and plants. In Methods in Applied Soil Microbiology and Biochemistry; Alef, K., Nannipieri, P., Eds.; Academic Press: London, UK, 1995; pp. 134–141. [Google Scholar]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophore. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Penrose, D.M.; Glick, B.R. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol. Plant. 2003, 118, 10–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Pérez-Romero, J.A.; Idaszkin, Y.L.; Barcia-Piedras, J.M.; Duarte, B.; Redondo-Gómez, S.; Caçador, I.; Mateos-Naranjo, E. Disentangling the effect of atmospheric CO2 enrichment on the halophyte Salicornia ramosissima J. Woods physiological performance under optimal and suboptimal saline conditions. Plant Physiol. Biochem. 2018, 127, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Redondo, S.; Rubio-Casal, A.E.; Castillo, J.M.; Luque, C.J.; Álvarez, A.A.; Luque, T.; Figueroa, M.E. Influences of salinity and light on germination of three Sarcocornia taxa with contrasted habitats. Aquat. Bot. 2004, 78, 255–264. [Google Scholar] [CrossRef]
- Mateos-Naranjo, E.; Andrades-Moreno, L.; Redondo-Gómez, S. Comparison of germination, growth, photosynthetic responses and metal uptake between three populations of Spartina densiflora under different soil pollution conditions. Ecotoxicol. Environ. Saf. 2011, 74, 2040–2049. [Google Scholar] [CrossRef] [PubMed]
- Gaskins, M.H.; Albrecht, S.L.; Hubbell, D.H. Rhizosphere bacteria and their use to increase plant productivity: A review. Agric. Ecosyst. Environ. 1985, 12, 99–116. [Google Scholar] [CrossRef]
- Olivares, F.L.; Busato, J.G.; de Paula, A.M.; da Silva Lima, L.; Aguiar, N.O.; Canellas, L.P. Plant growth promoting bacteria and humic substances: Crop promotion and mechanisms of action. Chem. Biol. Technol. Agric. 2017, 4, 30. [Google Scholar] [CrossRef]
- Zaidi, A.; Ahmad, E.; Khan, M.S.; Saif, S.; Rizvi, A. Role of plant growth promoting rhizobacteria in sustainable production of vegetables: Current perspective. Sci. Hortic. (Amst.) 2015, 193, 231–239. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Ahmad, M.; Zahir, Z.A.; Javaid, A.; Ashraf, M. The role of mycorrhizae and plant growth promoting rhizobacteria (PGPR) in improving crop productivity under stressful environments. Biotechnol. Adv. 2014, 32, 429–448. [Google Scholar] [CrossRef]
- Paul, D.; Lade, H. Plant-growth-promoting rhizobacteria to improve crop growth in saline soils: A review. Agron. Sustain. Dev. 2014, 34, 737–752. [Google Scholar] [CrossRef]
- Mahanty, T.; Bhattacharjee, S.; Goswami, M.; Bhattacharyya, P.; Das, B.; Ghosh, A.; Tribedi, P. Biofertilizers: A potential approach for sustainable agriculture development. Environ. Sci. Pollut. Res. 2017, 24, 3315–3335. [Google Scholar] [CrossRef] [PubMed]
- Timmusk, S.; Behers, L.; Muthoni, J.; Muraya, A.; Aronsson, A.-C. Perspectives and Challenges of Microbial Application for Crop Improvement. Front. Plant Sci. 2017, 8, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric. 2016, 2, 1127500. [Google Scholar] [CrossRef]
- Mateos-Naranjo, E.; Mesa, J.; Pajuelo, E.; Perez-Martin, A.; Caviedes, M.A.; Rodríguez-Llorente, I.D. Deciphering the role of plant growth-promoting rhizobacteria in the tolerance of the invasive cordgrass Spartina densiflora to physicochemical properties of salt-marsh soils. Plant Soil 2015, 394, 45–55. [Google Scholar] [CrossRef]
- Navarro-Torre, S.; Barcia-Piedras, J.M.; Caviedes, M.A.; Pajuelo, E.; Redondo-Gómez, S.; Rodríguez-Llorente, I.D.; Mateos-Naranjo, E. Bioaugmentation with bacteria selected from the microbiome enhances Arthrocnemum macrostachyum metal accumulation and tolerance. Mar. Pollut. Bull. 2017, 117, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Páliz, K.I.; Mateos-Naranjo, E.; Doukkali, B.; Caviedes, M.A.; Redondo-Gómez, S.; Rodríguez-Llorente, I.D.; Pajuelo, E. Modulation of Spartina densiflora plant growth and metal accumulation upon selective inoculation treatments: A comparison of gram negative and gram positive rhizobacteria. Mar. Pollut. Bull. 2017, 125, 77–85. [Google Scholar] [CrossRef]
- Nies, D.; Silver, S. Molecular Microbiology of Heavy Metals; Springer: Berlin/Heidelberg, Germany, 2007; ISBN 9783540697701. [Google Scholar]
- Roane, T.M.; Kellogg, S.T. Characterization of bacterial communities in heavy metal contaminated soils. Can. J. Microbiol. 1996, 42, 593–603. [Google Scholar] [CrossRef]
- Kamilova, F.; Okon, Y.; de Weert, S.; Hora, K. Commercialization of microbes: Manufacturing, inoculation, best practice for objective field testing, and registration. In Principles of Plant-Microbe Interactions; Springer: Berlin/Heidelberg, 2015; pp. 319–327. [Google Scholar]
- Ameixa, O.M.C.C.; Marques, B.; Fernandes, V.S.; Soares, A.M.V.M.; Calado, R.; Lillebø, A.I. Dimorphic seeds of Salicornia ramosissima display contrasting germination responses under different salinities. Ecol. Eng. 2016, 87, 120–123. [Google Scholar] [CrossRef]
- Duca, D.; Lorv, J.; Patten, C.L.; Rose, D.; Glick, B.R. Indole-3-acetic acid in plant–microbe interactions. Antonie Leeuwenhoek 2014, 106, 85–125. [Google Scholar] [CrossRef]
- Hara, Y.; Toriyama, K. Seed nitrogen accelerates the rates of germination, emergence, and establishment of rice plants. Soil Sci. Plant Nutr. 1998, 44, 359–366. [Google Scholar] [CrossRef]
- Agenbac, G.A.; Villiers, O.T. The effect of nitrogen fertilizers on the germination and seedling emergence of wild oat (A. fatua L.) seed in different soil types. Weed Res. 1989, 29, 239–245. [Google Scholar] [CrossRef]
- Gamalero, E.; Glick, B.R. Bacterial Modulation of Plant Ethylene Levels. Plant Physiol. 2015, 169, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Consortia Number | Hosting Halophyte | Sampling Location | Bacterial Strains | Reference |
|---|---|---|---|---|
| 1 | Spartina densiflora | Tinto estuary, SW Spain | Pseudomonas composti SDT3 Aeromonas aquariorum SDT13 Bacillus thuringiensis SDT14 | [30] |
| 2 | Arthrocnemun machrostachyum | Odiel estuary, SW Spain | Vibrio kanaloae RA1 Pseudoalteromonas prydzensis RA15 Staphylococcus warneri RA18 | [27] |
| 3 | Spartina maritima | Tinto and Odiel estuaries, SW Spain | Bacillus methylotrophicus SMT38 Bacillus aryabhattai SMT48 Bacillus licheniformis SMT51 Pantoea sp. RSO7 | [29,31] |
| 4 | Halimione portulacoides | Piedras estuary, SW Spain | Described in Section 3.2. | Present study |
| 5 | Salicornia ramosissima | Tinto and Piedras estuaries, SW Spain | Described in Section 3.2. | Present study |
| Strain | 16S rDNA Sequenced Fragment (bp) | Accession n. | Related Species | Identity (%) |
|---|---|---|---|---|
| HPJ2 | 1377 | MH304385 | Vibrio spartinae | 99.85 |
| HPJ9 | 1368 | MH304386 | Bacillus siamensis | 99.93 |
| HPJ15 | 1400 | MH304387 | Marinobacter sediminum | 99.21 |
| HPJ21 | 1397 | MH304388 | Bacillus aryabhattai | 100 |
| HPJ40 | 1398 | MH304389 | Bacillus zhangzhouensis | 99.79 |
| HPJ43 | 1410 | MH304390 | Bacillus zhangzhouensis | 99.79 |
| HPJ49 | 1328 | MH304391 | Bacillus subtilis | 100 |
| HPJ50 | 1390 | MH304392 | Vibrio parahaemolyticus | 99.78 |
| SRP14 | 1396 | MH304393 | Bacillus paralicheniformis | 100 |
| SRP15 | 1420 | MH304394 | Bacillus aryabhattai | 100 |
| SRT1 | 1397 | MH304395 | Vibrio neocaledonicus | 99.93 |
| SRT8 | 1334 | MH304396 | Thalassospira australica | 99.55 |
| SRT12 | 1071 | MH917124 | Halomonas taeanensis | 99.81 |
| SRT13 | 1411 | MH304397 | Vibrio alginolyticus | 99.65 |
| SRT14 | 1405 | MH304398 | Vibrio alginolyticus | 99.57 |
| SRT15 | 1377 | MH304399 | Pseudarthrobacter oxydans | 99.49 |
| Hosting Halophyte | Strain | NaCl Tolerance a | PGP Properties | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 28 ℃ | 40 ℃ | Nitrogen Fixation b | Phosphate Solubilization c | Siderophores Production c | IAA Production (mg/mL) | Biofilm Production b | ACC Deaminase Activity (µmol α-cetog h−1 mg prot−1) | ||
| Halimione portulacoides | HPJ2 | 1.2 | 1.2 | + | 12 | 30 | 4.12 | + | 1.86 |
| HPJ15 | 2 | 2 | - | - | 12 | 15.41 | - | - | |
| HPJ50 | 1.2 | 1.2 | + | 9 | 20 | 7.48 | + | - | |
| Salicornia ramosissima | SRT1 | 2 | 1.2 | + | 10 | 20 | 5.65 | + | - |
| SRT8 | 1.2 | 1.2 | - | - | - | - | + - | 1.24 | |
| SRT15 | 0.2 | 0.2 | + | 9 | - | 20.99 | - | - | |
| Parameter | Biomass | % Germination | FGD | MTG |
|---|---|---|---|---|
| CO2 | 0.01 ** | 0.00 ** | 0.87 | 0.00 ** |
| T | 0.12 | 0.00 ** | 0.08 | 0.00 ** |
| NaCl | 0.00 ** | 0.00 ** | 0.00 ** | 0.00 ** |
| PGPR | 0.00 ** | 0.00 ** | 0.19 | 0.00 ** |
| CO2 x T | 0.00 ** | 0.20 | 0.26 | 0.02 * |
| CO2 x NaCl | 0.64 | 0.00 ** | 0.63 | 0.03 * |
| CO2 x PGPR | 0.47 | 0.00 ** | 0.63 | 0.00 ** |
| T x NaCl | 0.71 | 0.01 ** | 0.17 | 0.03 * |
| T x PGPR | 0.00 ** | 0.28 | 0.62 | 0.41 |
| NaCl x PGPR | 0.00 ** | 0.00 ** | 0.81 | 0.00 ** |
| CO2 x T x NaCl | 0.05 * | 0.26 | 0.08 | 0.45 |
| CO2 x T x PGPR | 0.06 | 0.00 ** | 0.62 | 0.00 ** |
| CO2 x NaCl x PGPR | 0.02 * | 0.79 | 0.12 | 0.10 |
| T x NaCl x PGPR | 0.00 ** | 0.16 | 0.28 | 0.61 |
| CO2 x T x NaCl x PGPR | 0.01 ** | 0.00 ** | 0.89 | 0.00 ** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mesa-Marín, J.; Pérez-Romero, J.A.; Mateos-Naranjo, E.; Bernabeu-Meana, M.; Pajuelo, E.; Rodríguez-Llorente, I.D.; Redondo-Gómez, S. Effect of Plant Growth-Promoting Rhizobacteria on Salicornia ramosissima Seed Germination under Salinity, CO2 and Temperature Stress. Agronomy 2019, 9, 655. https://doi.org/10.3390/agronomy9100655
Mesa-Marín J, Pérez-Romero JA, Mateos-Naranjo E, Bernabeu-Meana M, Pajuelo E, Rodríguez-Llorente ID, Redondo-Gómez S. Effect of Plant Growth-Promoting Rhizobacteria on Salicornia ramosissima Seed Germination under Salinity, CO2 and Temperature Stress. Agronomy. 2019; 9(10):655. https://doi.org/10.3390/agronomy9100655
Chicago/Turabian StyleMesa-Marín, Jennifer, Jesús A. Pérez-Romero, Enrique Mateos-Naranjo, Miguel Bernabeu-Meana, Eloísa Pajuelo, Ignacio D. Rodríguez-Llorente, and Susana Redondo-Gómez. 2019. "Effect of Plant Growth-Promoting Rhizobacteria on Salicornia ramosissima Seed Germination under Salinity, CO2 and Temperature Stress" Agronomy 9, no. 10: 655. https://doi.org/10.3390/agronomy9100655