Impact of Genetic Background on the Leaf-Protective Enzyme Activity and Hormone Levels of Maize

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Conditions
2.2. Experimental Designs
2.3. Sampling and Measurements
2.4. Activity of SOD, CAT and POD, and the MDA Content
2.5. Content of Endogenous Hormones
2.6. Statistical Analysis
3. Results
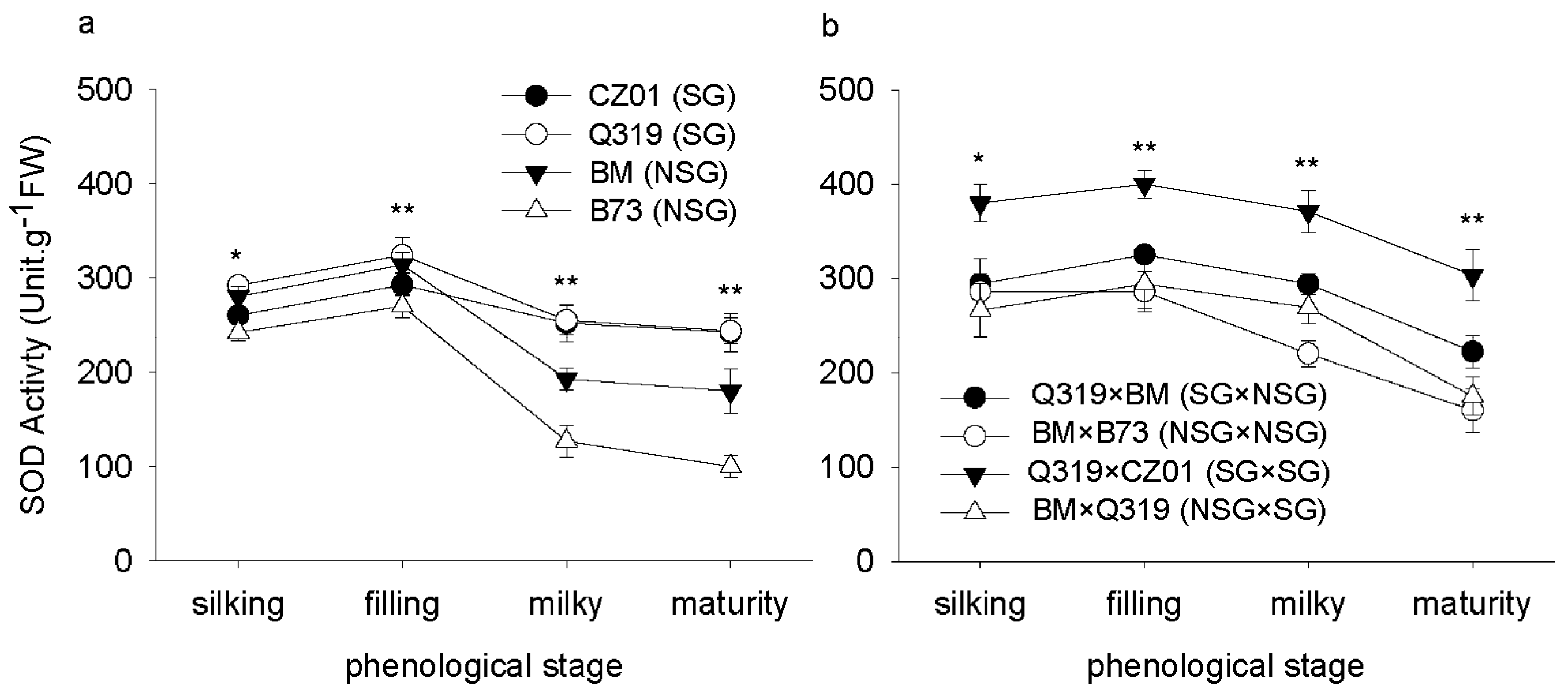
3.1. Superoxide Dismutase (SOD) Activity after Silking
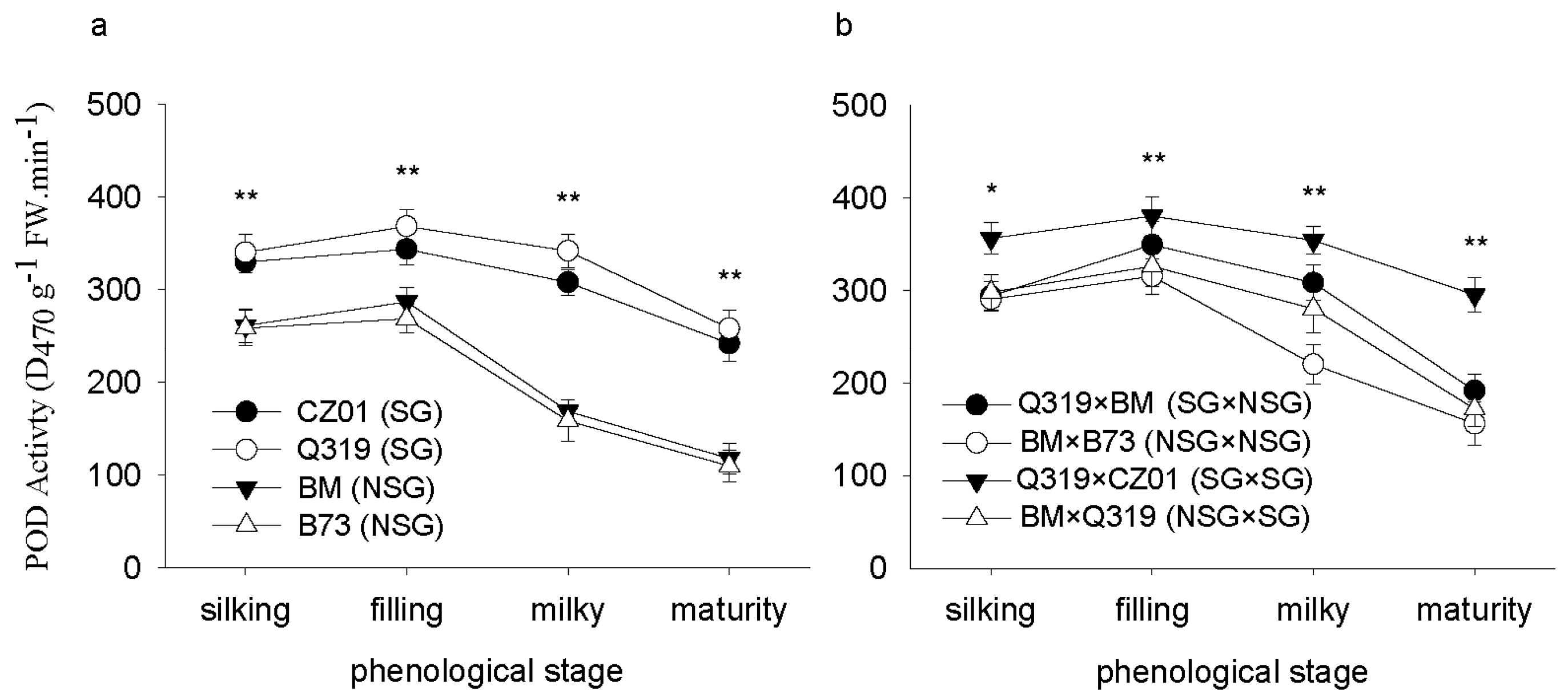
3.2. Peroxidase (POD) Activity after Silking
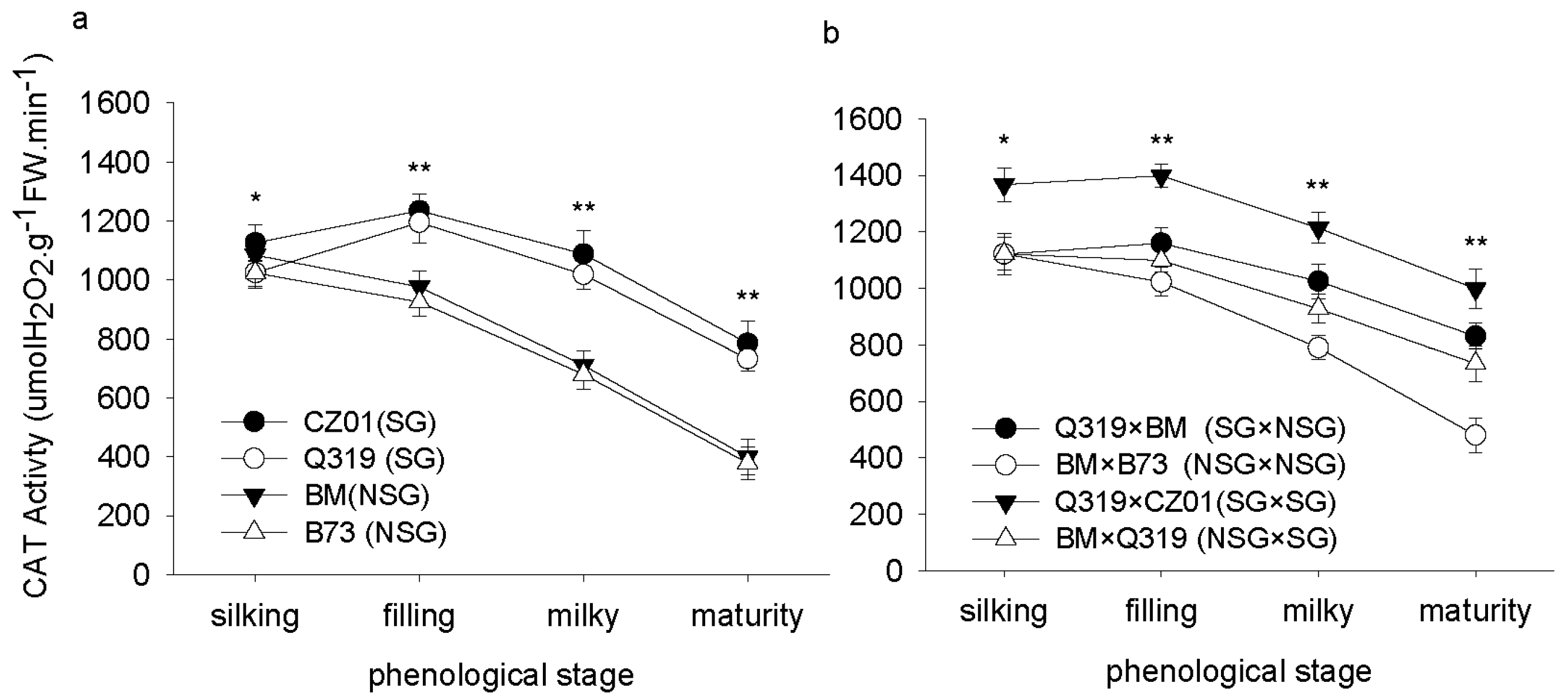
3.3. Catalase (CAT) Activity after Silking
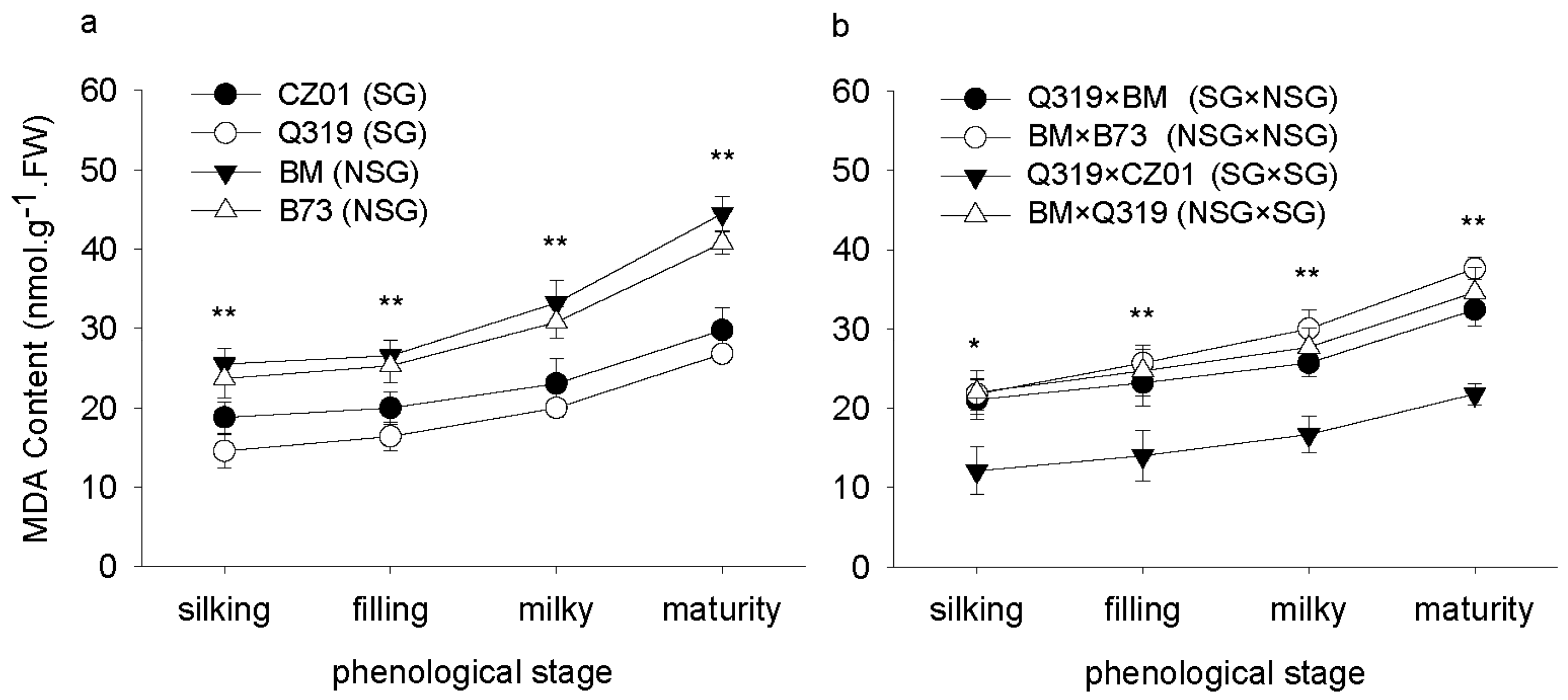
3.4. Malondialdehyde (MDA) Content after Silking
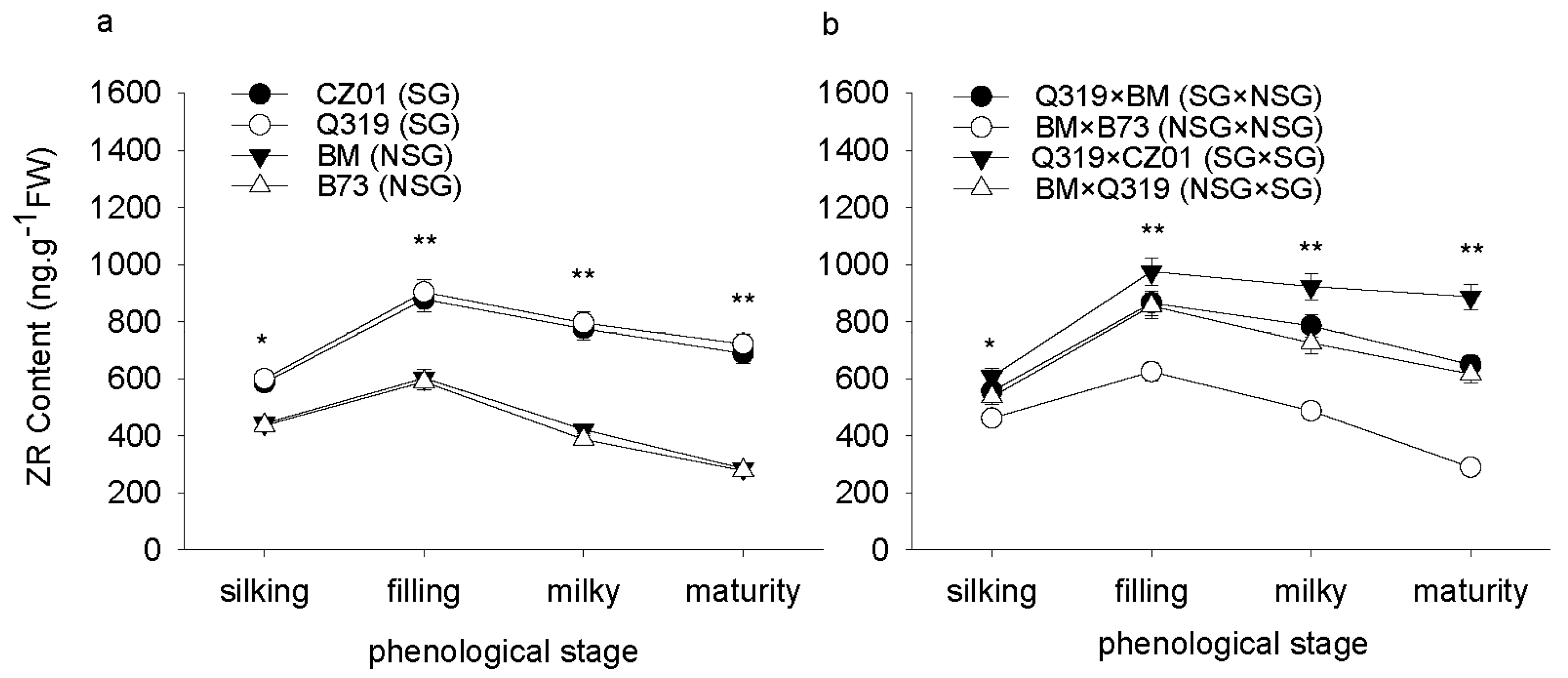
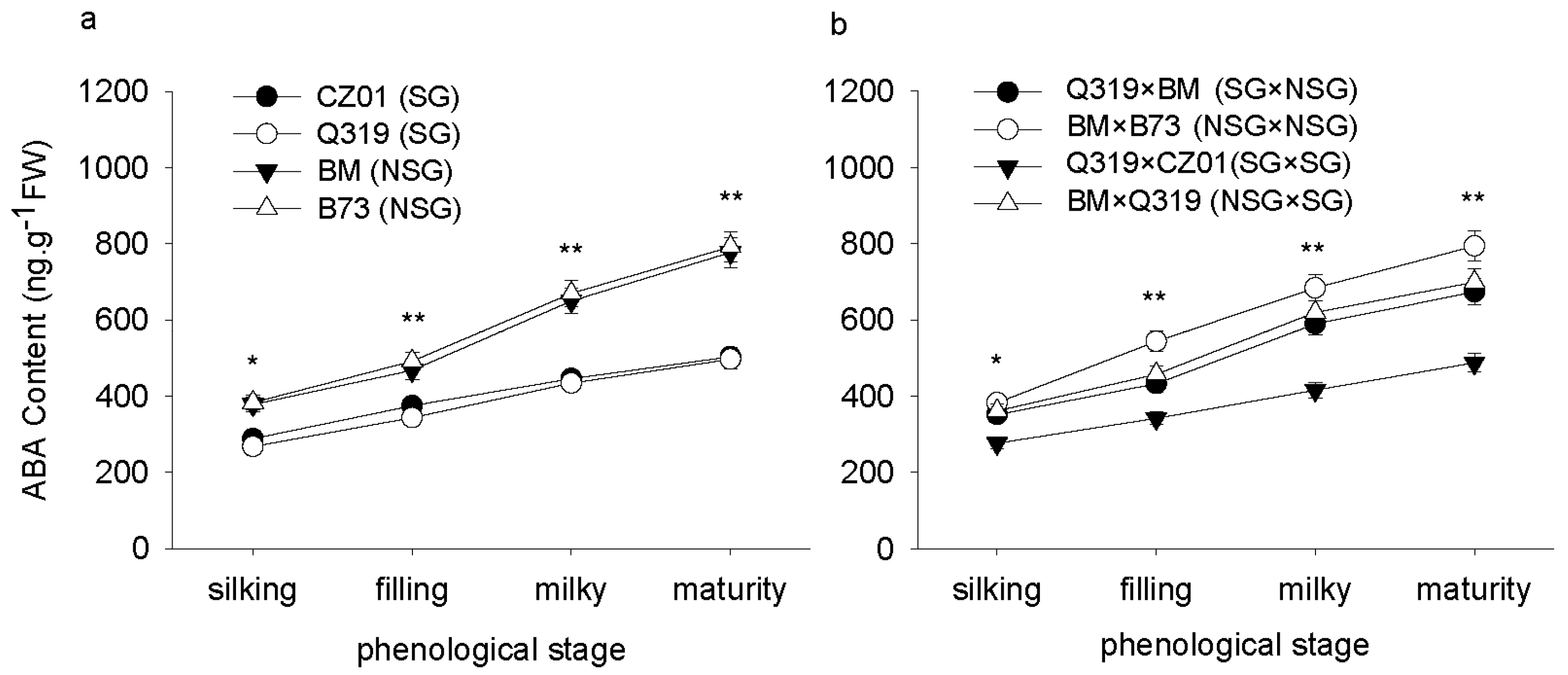
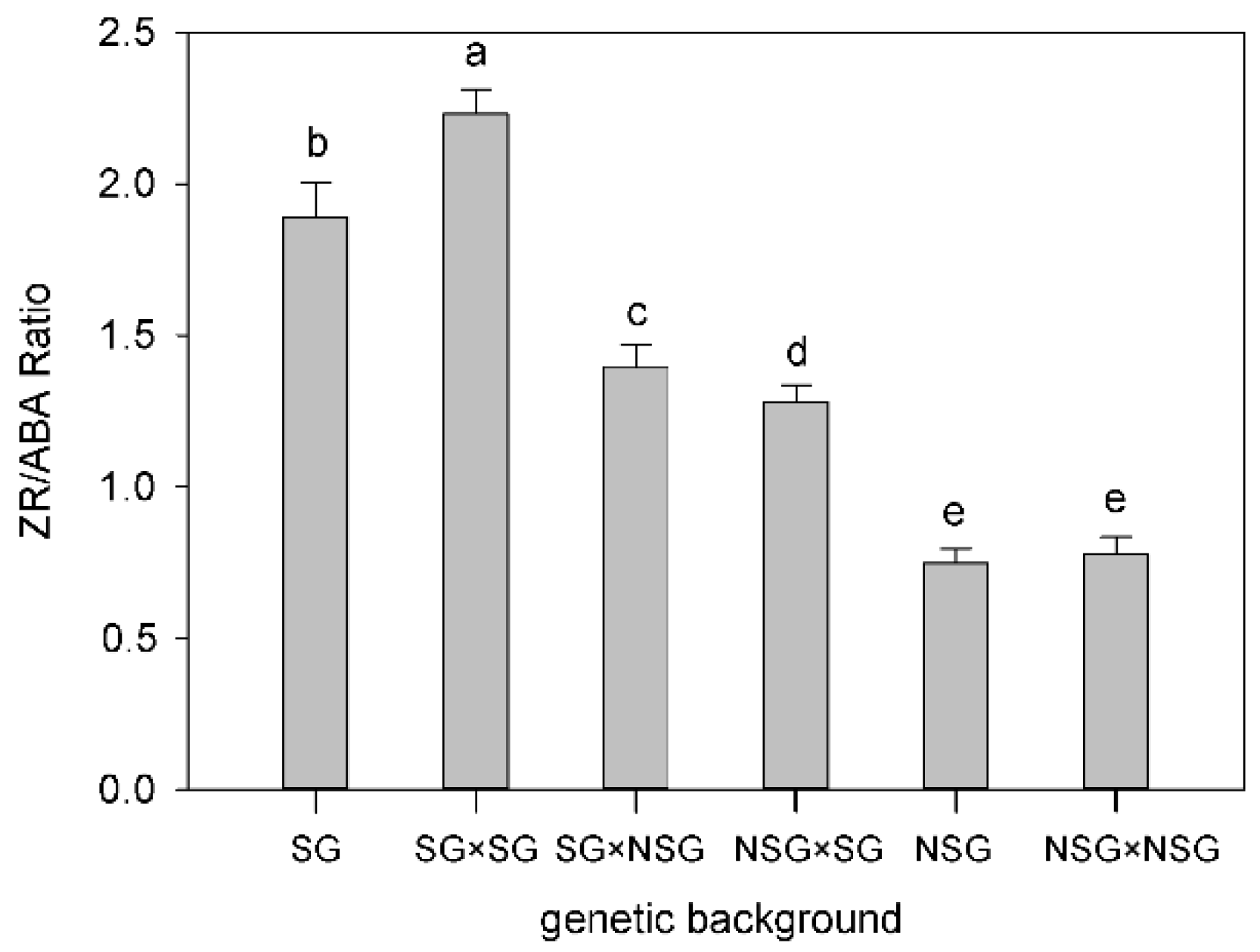
3.5. Hormone Content in Leaves after Silking
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wang, K.J.; Dong, S.T.; Hu, C.H.; Liu, K.; Zhang, J.W. The evolution of physiological characteristics of maize root during varietal replace in China, 1905s to 1990s II. Changes of the protective enzyme activities and lipid peroxidation. Acta Agron. Sin. 2002, 28, 384–388. [Google Scholar]
- Jagadish, K.S.V.; Kavi, K.P.B.; Bahuguna, R.N.; Nicolaus, V.W.; Nese, S. Staying alive or going to die during terminal senescence-an enigma surrounding yield stability. Front. Plant Sci. 2015, 6, 1070. [Google Scholar] [CrossRef] [PubMed]
- Antonietta, M.; Fanello, D.D.; Acciaresi, H.A.; Guiamet, J.J. Senescence and yield responses to plant density in stay green and earlier-senescing maize hybrids from Argentina. Field Crops Res. 2014, 155, 111–119. [Google Scholar] [CrossRef]
- Gregersen, P.L.; Culetic, A.; Boschian, L.; Krupinska, K. Plant Senescence and Crop Productivity. Plant Mol. Biol. 2013, 82, 603–622. [Google Scholar] [CrossRef] [PubMed]
- Belícuas, P.R.; Aguiar, A.M.; Bento, D.A.V.; Câmara, T.M.M.; Junior, C.L.S. Inheritance of the Stay-Green Trait in Tropical Maize. Euphytica 2014, 198, 163–173. [Google Scholar] [CrossRef]
- Renu, K. Leaf senescence and abiotic stresses share reactive oxygen species-mediated chloroplast degradation. Protoplasma 2012, 249, 469–481. [Google Scholar]
- Smith, K.; Craven, M.; Mclaren, N.W.; Bemer, J.M. Effects on fungicide application on sorghum yield, leaf senescence and photosynthetic efficiency in South Africa. S. Afr. J. Bot. 2015, 121, 201. [Google Scholar] [CrossRef]
- Vietor, D.M.; Rhodes, G.A.; Rooney, W.L. Relationship of phenotypic variation in sorghum to nutritive value of crop residues. Field Crops Res. 2010, 118, 243–250. [Google Scholar] [CrossRef]
- Spano, G.; Di, F.N.; Perrotta, C.; Platani, C.; Ronga, G.; Lawlor, D.W.; Napier, J.A.; Shewry, P.R. Physiological characterization of ‘stay green’ mutants in durum wheat. J. Exp. Bot. 2003, 54, 1415–1420. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.L.; Xie, R.Z.; Zhang, F.L.; Li, J.; Li, S.M.; Long, H.L.; Liu, Y.; Guo, Y.Q.; Li, S.K. Genetic contribution to maize yield gain among different locations in China. Maydice 2016, 6, 1070. [Google Scholar]
- Zhang, X.X.; Wang, M.; Wu, T.T.; Wu, C.X.; Jiang, B.J.; Guo, C.H.; Han, T.F. Physiological and molecular studies of stay-green caused by pod removal and seed injury in soybean. Crop J. 2016, 6, 435–443. [Google Scholar] [CrossRef]
- Wingler, A.; Von, S.A.; Leegood, R.C.; Lea, P.J.; Quick, W.P. Regulation of leaf senescence by cytokinin, sugars, and light. Effects on NADH-dependent hydroxypyruvate reductase. Plant Physiol. 1998, 116, 329–335. [Google Scholar] [CrossRef]
- Hu, T.; Yuan, L.; Wang, J.; Kang, S.; Li, F. Antioxidation responses of maize roots and leaves to partial root-zone irrigation. Agric. Water Manag. 2010, 98, 164–171. [Google Scholar] [CrossRef]
- Blumwald, E. Sodium transport and salt tolerance in plants. Curr. Opin. Cell Biol. 2000, 12, 431–434. [Google Scholar] [CrossRef]
- Suzuki, N.; Shai, K.; Ron, M.; Gad, M. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ. 2012, 35, 259–270. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Jin, J.Y. Effect of N and K nutrition on changes of endogenous hormone and metabolism of active oxygen during leaf senescence in spring maize. J. Plant Nutr. Fertil. 1999, 4, 289–296. [Google Scholar]
- Humbeck, K.; Quast, S.; Krupinska, K. Functional and molecular changes in the photosynthetic apparatus during senescence of flag leaves from field-grown barley plants. Plant Cell Environ. 1996, 19, 337–344. [Google Scholar] [CrossRef]
- Markovich, O.; Steiner, E.; Kouřil, Š.; Tarkowski, P.; Aharoni, A.; Elbaum, R. Silicon promotes cytokinin biosynthesis and delays senescence in Arabidopsis and Sorghum. Plant Cell Environ. 2017, 40, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Raines, T.; Shanks, C.; Cheng, C.Y.; McPherson, D.; Argueso, C.T.; Kim, H.J.; Franco-Zorrilla, J.M.; López-Vidriero, I.; Solano, R.; Vaňková, R.; et al. The cytokinin response factors modulate root and shoot growth and promote leaf senescence in Arabidopsis. Plant J. 2016, 85, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Borrell, A.K.; Hammer, G.L.; Henzell, R.G. Does maintaining green leaf area in sorghum improve yield under drought? II. Dry matter production and yield. Crop Sci. 2000, 40, 1037–1048. [Google Scholar] [CrossRef]
- Liu, K.C.; Dong, S.T.; Zhao, H.J.; Wang, Q.C.; Li, Z.X.; Liu, X.; Zhang, H. Leaf stay-green traits in Chinese maize inbred lines and their relationship with grain yield. Acta Agron. Sin. 2009, 35, 1662–1671. [Google Scholar] [CrossRef]
- Wang, K.J.; Hu, C.H.; Dong, S.T.; Liu, K.C.; Sun, Q.Q. Changes of the protective enzyme activities and lipid peroxidation after anthesis among maize varieties planted in different years. Acta Agron. Sin. 1999, 25, 700–706. [Google Scholar]
- Zhang, Z.; Li, G.; Gao, H.; Zhang, L.; Yang, C.; Liu, P.; Meng, Q. Characterization of Photosynthetic Performance during Senescence in Stay-Green and Quick-Leaf-Senescence Zea mays L. Inbred Lines. PLoS ONE 2012, 7, e42936. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhao, H.; Dong, S.; Wang, Q.; Li, Z.; Liu, K. Study on Characteristics of Nitrogen Metabolism in Diallel Cross Generation of Different Maize Genotypes After Silking. Sci. Agric. Sin. 2014, 47, 33–42. [Google Scholar]
- Winterbourn, C.; Hawkins, R.; Brian, M.; Carrell, R. The estimation of red cell superoxide dismutase activity. J. Lab. Clin. Med. 1975, 85, 337–341. [Google Scholar] [PubMed]
- Zhang, Z.O.; Pang, X.Q.; Xue, W.D.; Ji, Z.L.; Jiang, Y.M. Role of peroxidase in anthocyanin degradation in litchi fruit pericarp. Food Chem. 2005, 90, 47–52. [Google Scholar] [CrossRef]
- Schmedes, A.; Hølmer, G. A new thiobarbituric acid (TBA) method for determining free malondialdehyde (MDA) and hydroperoxides selectively as a measure of lipid peroxidation. J. Am. Oil Chem. Soc. 1989, 66, 813–817. [Google Scholar] [CrossRef]
- Liu, K.C. Study on Physiological Mechanism and Inheritance of Leaf Stay-Green Traits of Different Maize (Zea mays L.) Genotypes; Shandong Agricultural University: Tai’an, China, 2003. [Google Scholar]
- Fridovich, I. Superoxide dismutases. Annu. Rev. Biochem. 1975, 44, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Prochazkova, D.; Sairam, R.K.; Srivastava, G.C.; Singh, D.V. Oxidative stress and antioxidant activity as the basis of senescence in maize leaves. Plant Sci. 2001, 161, 765–771. [Google Scholar] [CrossRef]
- Xue, H.; Jia, L.; Gong, Y.H.; Liu, Y.Z.; Wu, Y.H. Study on the stay-green capacity and leaf senescence of winter wheat. Acta Bot. Boreal. Occident. Sin. 2010, 30, 0336–0343. [Google Scholar]
- Wang, F.; Liu, J.; Zhou, L.; Pan, G.; Li, Z.; Zaidi, S.H.R.; Cheng, F. Senescence-specific change in ROS scavenging enzyme activities and regulation of various SOD isozymes to ROS levels in Psf mutant rice leaves. Plant Physiol. Biochem. 2016, 109, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Asada, K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Vyroubalová, Š.; Václavíková, K.; Turečková, V.; Novák, O.; Šmehilová, M.; Hluska, T.; Ohnoutková, L.; Frébort, I.; Galuszka, P. Characterization of new maize genes putatively involved in cytokinin metabolism and their expression during osmotic stress in relation to cytokinin levels. Plant Physiol. 2009, 151, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Badenoeh-Jones, J.; Parker, C.W.; Letham, D.S. Effect of cytokinin supplied via the xylem at multiples of endogenous concentration on transpiration and senescence in derooted seedlings of oat and wheat. Plant Cell Environ. 1996, 19, 504–516. [Google Scholar] [CrossRef]
- Gan, S.; Amasino, R.M. Inhibition of Leaf Senescence by Autoregulated Production of Cytokinin. Science 1995, 270, 1986–1988. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Zhu, J.K. Regulation of abscisic acid biosynthesis. Plant Physiol. 2003, 133, 29–36. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Generation | Material Code | Hybrid Lines |
|---|---|---|
| Self-Cross | SG1 | CZ01 |
| SG2 | Q319 | |
| NSG1 | BM | |
| NSG2 | B73 | |
| Hybrid Cross | SG1 × SG2 | CZ01 × Q319 |
| SG2 × SG1 | Q319 × CZ01 | |
| SG1 × NSG1 | CZ01 × BM | |
| SG2 × NSG1 | Q319 × BM | |
| NSG1 × SG1 | BM × ZC01 | |
| NSG1 × SG2 | BM × Q319 | |
| NSG1 × NSG2 | BM × B73 | |
| NSG2 × NSG1 | B73 × BM |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.; Li, Z.; Liu, T.; Zhang, H.; Zhao, H.; Wang, Q.; Shi, Y.; Dai, H.; Liu, K. Impact of Genetic Background on the Leaf-Protective Enzyme Activity and Hormone Levels of Maize. Agronomy 2018, 8, 234. https://doi.org/10.3390/agronomy8100234
Liu C, Li Z, Liu T, Zhang H, Zhao H, Wang Q, Shi Y, Dai H, Liu K. Impact of Genetic Background on the Leaf-Protective Enzyme Activity and Hormone Levels of Maize. Agronomy. 2018; 8(10):234. https://doi.org/10.3390/agronomy8100234
Chicago/Turabian StyleLiu, Chunxiao, Zongxin Li, Tieshan Liu, Hui Zhang, Haijun Zhao, Qingcheng Wang, Yonghui Shi, Hongcui Dai, and Kaichang Liu. 2018. "Impact of Genetic Background on the Leaf-Protective Enzyme Activity and Hormone Levels of Maize" Agronomy 8, no. 10: 234. https://doi.org/10.3390/agronomy8100234