Seedling Growth and Transcriptional Responses to Salt Shock and Stress in Medicago sativa L., Medicago arborea L., and Their Hybrid (Alborea)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Seed Pretreatment
2.3. Growth Conditions and Salt Stress Treatments
2.4. Growth Characteristics Measurements
2.5. Determination of Na+ and K+ Contents
2.6. RNA Isolation and Complementary DNA Synthesis
2.7. Real-Time PCR Experiments
2.8. Statistical Analysis
3. Results
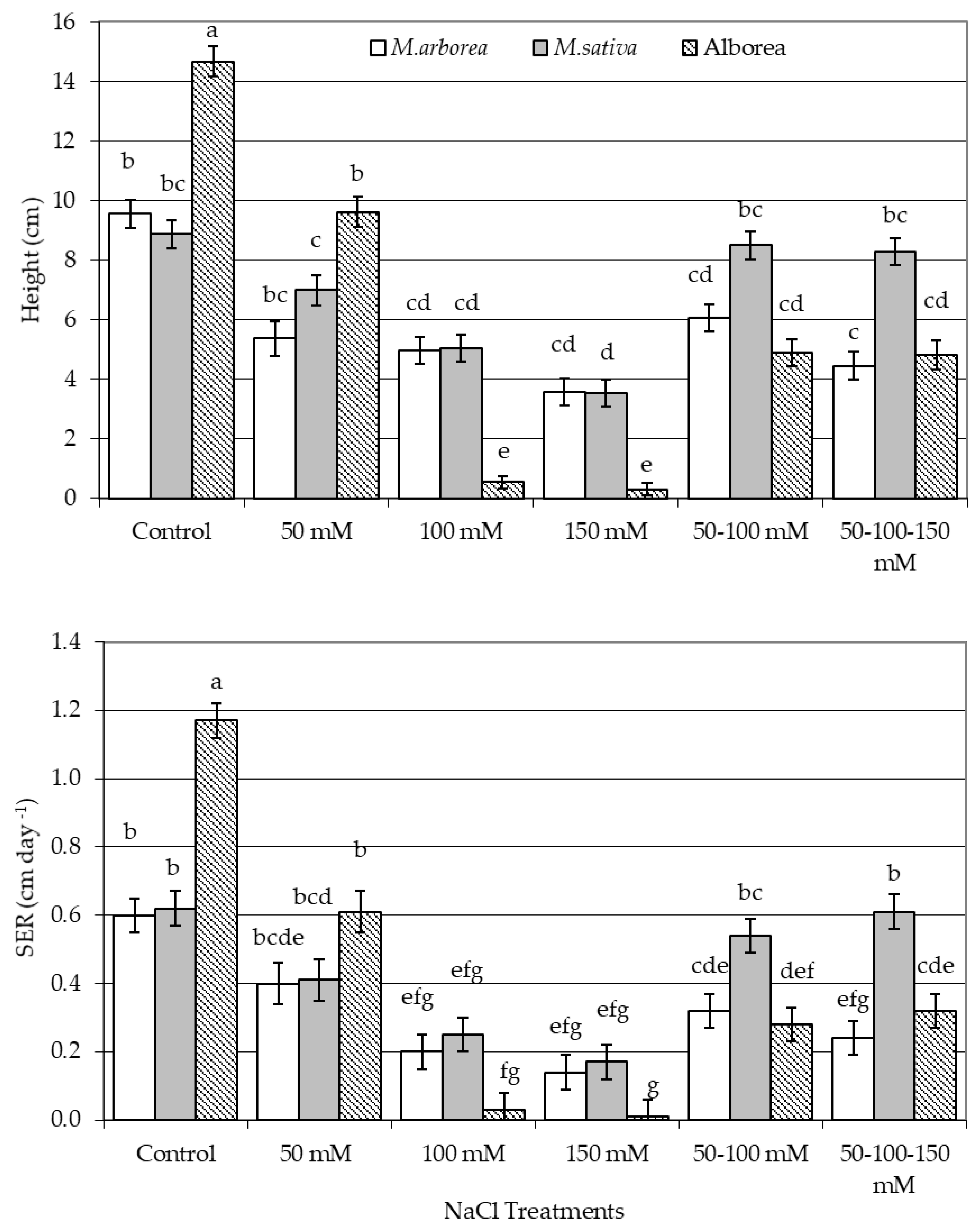
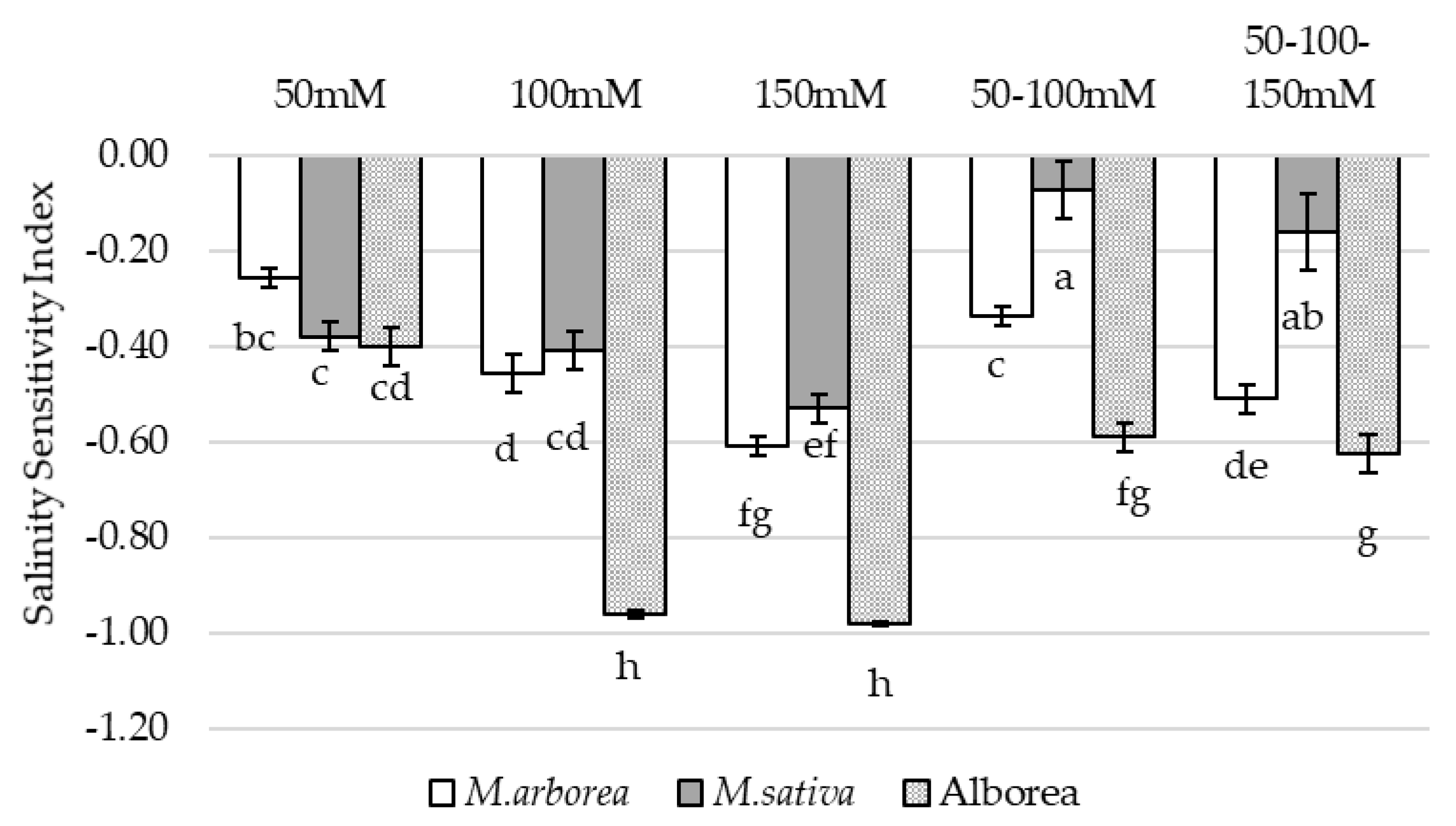
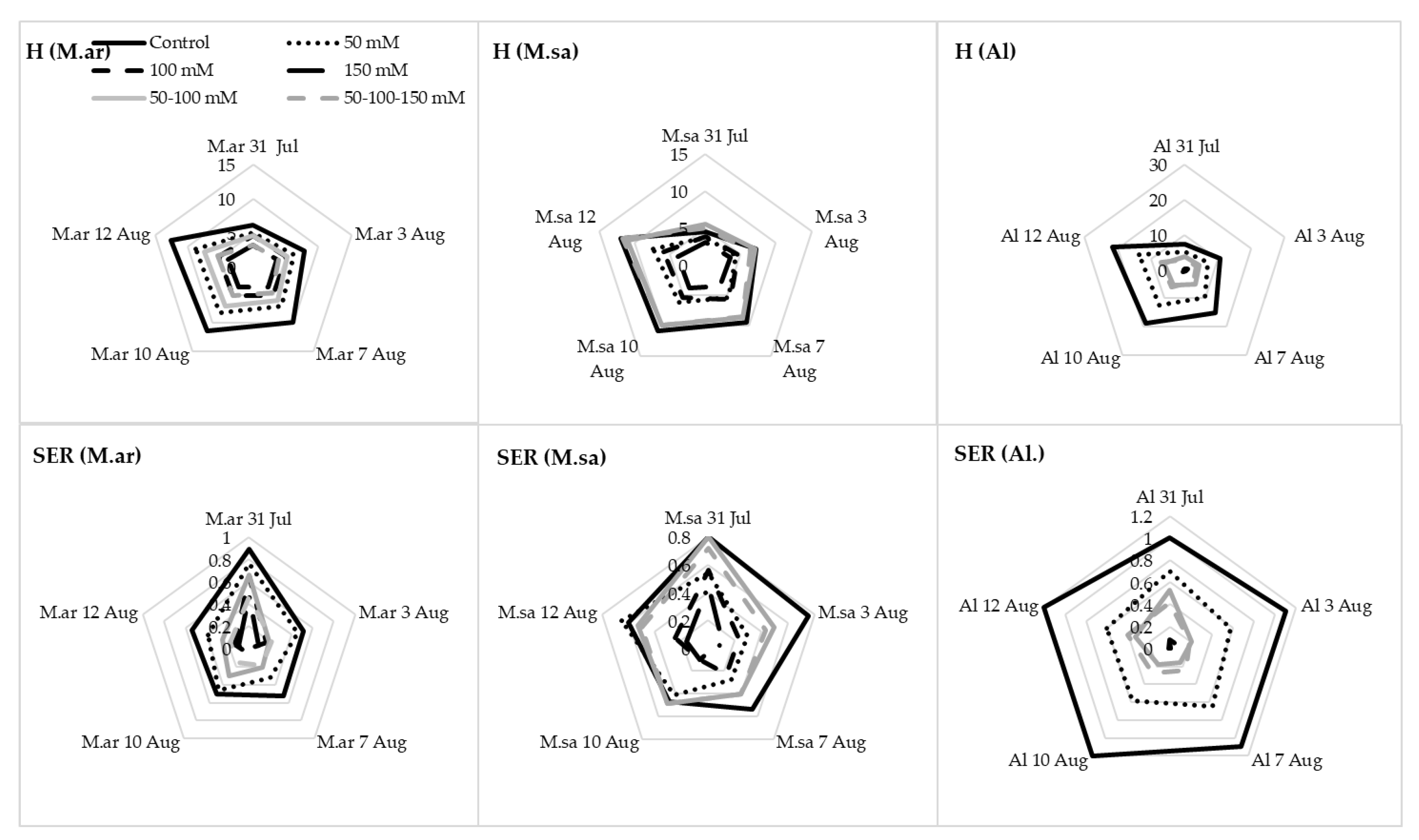
3.1. Growth Parameters and Salinity Sensitivity Index
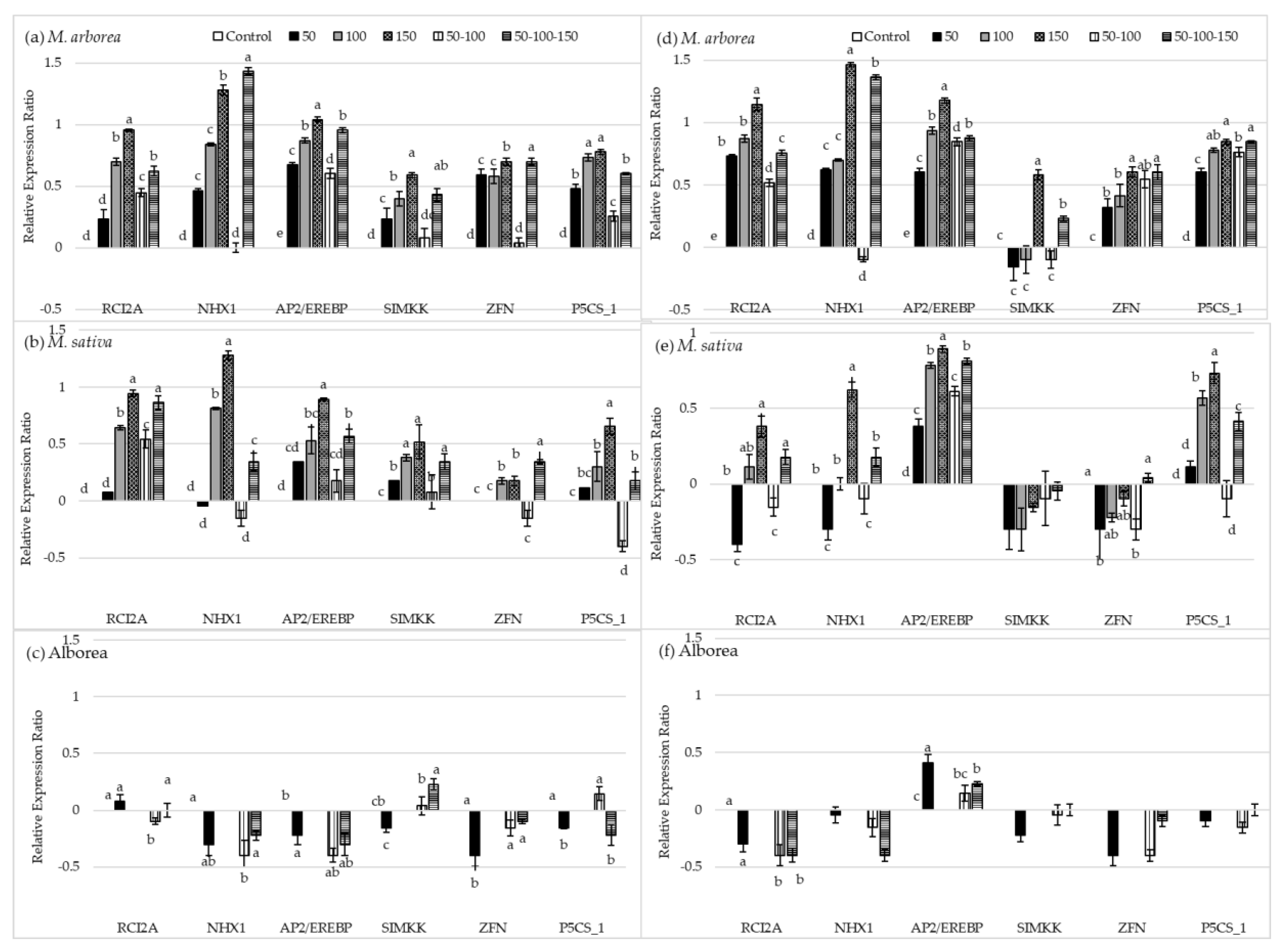
3.2. Gene Expression Levels
4. Discussion
4.1. Growth of Seedlings under Salinity
4.2. Gene Expression under Salt Treatments
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pitman, M.G.; Läuchli, A. Global impact of salinity and agricultural ecosystems. In Salinity: Environment—Plants—Molecules; Läuchli, A., Luttge, U., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 3–20. [Google Scholar]
- Rengasamy, P. Soil chemistry factors confounding crop salinity tolerance—A review. Agronomy 2016, 6, 53. [Google Scholar] [CrossRef]
- Young, J.; Udeigwe, T.K.; Weindorf, D.C.; Kandakji, T.; Gautam, P.; Mahmoud, M.A. Evaluating management-induced soil salinization in golf courses in semi-arid landscapes. Solid Earth 2015, 6, 393–402. [Google Scholar] [CrossRef] [Green Version]
- Pariente, S. Soluble salts dynamics in the soil under different climatic conditions. Catena 2001, 43, 307–321. [Google Scholar] [CrossRef]
- Ispail, A.; Takeda, S.; Nick, P. Life and death under salt stress: Same players, different timing? J. Exp. Bot. 2014, 65, 2963–2979. [Google Scholar]
- Munns, R.; James, R.A. Screening methods for salinity tolerance: A case study with tetraploid wheat. Plant. Soil 2003, 253, 201–218. [Google Scholar] [CrossRef]
- Shavrukov, Y.; Langridge, P.; Tester, M. Salinity tolerance and sodium exclusion in genus Triticum. Breed. Sci. 2009, 59, 671–678. [Google Scholar] [CrossRef]
- Apse, M.P.; Aharon, G.S.; Snedden, W.A.; Blumwald, E. Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science 1999, 285, 1256–1258. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shavrukov, Y. Salt stress or salt shock: Which genes are we studying? J. Exp. Bot. 2013, 64, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Genes and salt tolerance: Bringing them together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, K.; Sairam, R.K.; Bhattacharya, R.C. Differential expression of salt overly sensitive pathway genes determines salinity stress tolerance in Brassica genotypes. Plant Physiol. Biochem. 2012, 51, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Roshandel, P.; Flowers, T. The ionic effects of NaCl on physiology and gene expression in rice genotypes differing in salt tolerance. Plant Soil 2009, 315, 135–147. [Google Scholar] [CrossRef]
- Sanchez, D.H.; Lippold, F.; Redestig, H.; Hannah, M.A.; Erban, A.; Krämer, U.; Kopka, J.; Udvardi, M.K. Integrative functional genomics of salt acclimatization in the model legume Lotus japonicus. Plant J. 2008, 53, 973–987. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Shi, F.; Fukuda, K.; Yang, Y. Effects of salt and alkali stresses on germination, growth, photosynthesis and ion accumulation in alfalfa (Medicago sativa L.). Soil Sci. Plant. Nutr. 2010, 56, 725–733. [Google Scholar] [CrossRef]
- Yang, Q.C.; Wu, M.S.; Wang, P.Q.; Kang, J.M.; Zhou, X.L. Cloning and expression analysis of a vacuolar Na+/H+ antiporter gene from alfalfa. DNA Seq. 2005, 16, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.F.S.; Cornacchione, M.V.; Liu, X.; Suarez, D.L. Nutrient composition, forage parameters, and antioxidant capacity of alfalfa (Medicago sativa, L.) in response to saline irrigation water. Agriculture 2015, 5, 577–597. [Google Scholar] [CrossRef]
- Graham, P.H.; Vance, C.P. Legumes: Importance and constraints to greater use. Plant Physiol. 2003, 131, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Campanelli, A.; Ruta, C.; Morone-Fortunato, I.; De Mastro, G. Alfalfa (Medicago sativa L.) clones tolerant to salt stress: In vitro selection. Cent. Eur. J. Biol. 2013, 8, 765–776. [Google Scholar] [CrossRef]
- Postnikova, O.A.; Shao, J.; Nemchinov, L.G. Analysis of the alfalfa root transcriptome in response to salinity stress. Plant Cell Physiol. 2013, 54, 1041–1055. [Google Scholar] [CrossRef] [PubMed]
- Quan, W.L.; Liu, X.; Wang, H.Q.; Chan, Z.L. Physiological and transcriptional responses of contrasting alfalfa (Medicago sativa L.) varieties to salt stress. Plant Cell Tissue Organ Cult. 2016, 126, 105–115. [Google Scholar] [CrossRef]
- Sandhu, D.; Cornacchione, M.V.; Ferreira, J.F.S.; Suarez, D.L. Variable salinity responses of 12 alfalfa genotypes and comparative expression analyses of salt-response genes. Sci. Rep. 2017, 7, 42958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boughalleb, F.; Denden, M.; Ben Tiba, B. Photosystem II photochemistry and physiological parameters of three fodder shrubs, Nitraria retusa, Atriplex halimus and Medicago arborea under salt stress. Acta Physiol. Plant. 2009, 31, 463–476. [Google Scholar] [CrossRef]
- Boughalleb, F.; Denden, M.; Tiba, B.B. Anatomical changes induced by increasing NaCl salinity in three fodder shrubs, Nitraria retusa, Atriplex halimus and Medicago arborea. Acta Physiol. Plant. 2009, 31, 947–960. [Google Scholar] [CrossRef]
- Lefi, E.; Conesa, M.À.; Cifre, J.; Gulías, J.; Medrano, H. Dry matter allocation in Medicago arborea and Medicago citrina in response to drought and defoliation. Crop Pasture Sci. 2012, 63, 179–189. [Google Scholar] [CrossRef]
- Bingham, E.T.; Armour, D.; Irwin, J.A.G. The hybridization barrier between herbaceous Medicago sativa and woody M. arborea is weakened by selection of seed parents. Plants 2013, 2, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Irwin, J.A.G.; Sewell, J.C.; Woodfield, D.R.; Bingham, E.T. Restructuring lucerne (Medicago sativa) through introgression of the Medicago arborea genome. Agric. Sci. 2016, 28, 40–46. [Google Scholar]
- Clifton-Brown, J.C.; Lewandowski, I. Water use efficiency and biomass partitioning of three different Miscanthus genotypes with limited and unlimited water supply. Ann. Bot. 2000, 86, 191–200. [Google Scholar] [CrossRef]
- Lei, Y.; Xu, Y.; Hettenhausen, C.; Lu, C.; Shen, G.; Zhang, C.H.; Li, J.G.; Song, J.; Lin, H.; Wu, J. Comparative analysis of alfalfa (Medicago sativa L.) leaf transcriptomes reveals genotype-specific salt tolerance mechanisms. BMC Plant Biol. 2018, 18, 35. [Google Scholar] [CrossRef] [PubMed]
- Leidi, E.O.; Barragán, V.; Rubio, L.; El-Hamdaoui, A.; Ruiz, M.T.; Cubero, B.; Fernández, J.A.; Bressan, R.A.; Hasegawa, P.M.; Quintero, F.J.; et al. The AtNHX1 exchanger mediates potassium compartmentation in vacuoles of transgenic tomato. Plant J. 2010, 61, 495–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, R.C.; Zhang, F.; Li, Z.Y.; Li, M.N.; Cong, L.L.; Kang, J.M.; Zhang, T.J.; Zhao, Z.X.; Sun, Y.; Yang, Q.C. Isolation and functional characterization of salt-stress induced RCI2-like genes from Medicago sativa and Medicago truncatula. J. Plant Res. 2015, 128, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Mitsuya, S.; Taniguchi, M.; Miyake, H.; Takabe, T. Overexpression of RCI2A decreases Na+ uptake and mitigates salinity-induced damages in Arabidopsis thaliana plants. Physiol. Plant. 2006, 128, 95–102. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B. Proline: A key player in plant abiotic stress tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Wang, X.H.; Gao, B.W.; Liu, X.; Dong, X.J.; Zhang, Z.X.; Fan, H.Y.; Zhang, L.; Wang, J.; Shi, S.P.; Tu, P.F. Salinity stress induces the production of 2-(2-phenylethyl)chromones and regulates novel classes of responsive genes involved in signal transduction in Aquilaria sinensis calli. BMC Plant Biol. 2016, 16, 119. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guo, Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Suh, A. Evaluation of Bioactivity of Phytotoxins from Pathogenic Fungi of Orobanche sp. Ph.D. Thesis, Agricultural University of Athens, Athina, Greece, 2011. [Google Scholar]
- Hamrouni, L.; Ben Abdallah, F.; Abdelly, C.; Ghorbel, A. La culture in vitro: Un moyen rapide et efficace pour sélectionner des génotypes de vigne tolérant la salinité. C. R. Biol. 2008, 331, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using realtime quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Beyer, H.T.; John, W. Exploratory Data Analysis. Addison-Wesley Publishing Company Reading, Mass.—Menlo Park, Cal., London, Amsterdam, Don Mills, Ontario, Sydney 1977, XVI, 688 S. Biom. J. 1981, 23, 413–414. [Google Scholar] [CrossRef]
- Almansouri, M.; Kinet, J.-M.; Lutts, S. Effect of salt and osmotic stresses on germination in durum wheat (Triticum durum Desf.). Plant Soil 2001, 231, 243–254. [Google Scholar] [CrossRef]
- Zou, J.; Liu, C.F.; Liu, A.L.; Zou, D.; Chen, X.B. Overexpression of OsHsp17.0 and OsHsp23.7 enhances drought and salt tolerance in rice. J. Plant Physiol. 2012, 169, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Scasta, J.D.; Trostle, C.L.; Foster, M.A. Evaluating alfalfa (Medicago sativa L.) cultivars for salt tolerance using laboratory, greenhouse and field methods. J. Agric. Sci. 2012, 4, 90. [Google Scholar] [CrossRef]
- Cornacchione, M.V.; Suarez, D.L. Evaluation of alfalfa (Medicago sativa L.) populations’ response to salinity stress. Crop Sci. 2017, 57, 137–150. [Google Scholar] [CrossRef]
- Farissi, M.; Bouizgaren, A.; Faghire, M.; Bargaz, A.; Ghoulam, C. Agro-physiological responses of Moroccan alfalfa (Medicago sativa L.) populations to salt stress during germination and early seedling stages. Seed Sci. Technol. 2011, 39, 389–401. [Google Scholar] [CrossRef]
- Boughalleb, F.; Hajlaoui, H.; Mhamdi, M.; Dendon, M. Possible involvement of organic compounds and the antioxidant defense system in salt tolerance of Medicago arborea (L.). Agric. J. 2011, 6, 353–365. [Google Scholar] [CrossRef]
- Sibole, J.V.; Cabot, C.; Poschenrieder, C.; Barcelo, J. Ion allocation in two different salt-tolerant Mediterranean Medicago species. J. Plant Physiol. 2003, 160, 1361–1365. [Google Scholar] [CrossRef] [PubMed]
- Galmes, J.; Cifre, J.; Medrano, H.; Flexas, J. Modulation of relative growth rate and its components by water stress in Mediterranean species with different growth forms. Oecologia 2005, 145, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Teakle, N.L.; Amtmann, A.; Real, D.; Colmer, T.D. Lotus tenuis tolerates combined salinity and waterlogging: Maintaining O2 transport to roots and expression of an NHX1-like gene contribute to regulation of Na+ transport. Physiol. Plant. 2010, 139, 358–374. [Google Scholar] [CrossRef] [PubMed]
- Rocha, P.S.C.F. Plant abiotic stress-related RCI2/PMP3s: Multigenes for multiple roles. Planta 2016, 244, 287. [Google Scholar] [CrossRef] [PubMed]
- Sairam, R.K.; Tyagi, A. Physiology and molecular biology of salinity stress tolerance in plants. Curr. Sci. 2004, 86, 407–421. [Google Scholar]
- Gharsallah, C.; Fakhfakh, H.; Grubb, D.; Gorsane, F. Effect of salt stress on ion concentration, proline content, antioxidant enzyme activities and gene expression in tomato cultivars. AOB Plants 2016, 8, plw055. [Google Scholar] [CrossRef] [PubMed]
- Silva-Ortega, C.O.; Ochoa-Alfaro, A.E.; Reyes-Agüero, J.A.; Aguado-Santacruz, G.A.; Jiménez-Bremont, J.F. Salt stress increases the expression of P5CS gene and induces proline accumulation in cactus pear. Plant Physiol. Biochem. 2008, 46, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AP2/ERF family transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta 2012, 1819, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol. 2006, 140, 411–432. [Google Scholar] [CrossRef] [PubMed]
- Mizukami, Y.; Fischer, R.L. Plant organ size control: AINTEGUMENTA regulates growth and cell numbers during organogenesis. Proc. Natl. Acad. Sci. USA 2000, 97, 942–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, L.S.; Wang, Y.B.; Yao, S.Q.; Liu, A.Z. Arabidopsis AINTEGUMENTA mediates salt tolerance by trans-repressing SCABP8. J. Cell Sci. 2015, 128, 2919–2927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, A.K.; Jaggi, M.; Raghuram, B.; Tuteja, N. Mitogen-activated protein kinase signaling in plants under abiotic stress. Plant Signal. Behav. 2011, 6, 196–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiegerl, S.; Cardinale, F.; Siligan, C.; Gross, A.; Baudouin, E.; Liwosz, A.; Eklof, S.; Till, S.; Bogre, L.; Hirt, H.; et al. SIMKK, a mitogen-activated protein kinase (MAPK) kinase, is a specific activator of the salt stress-induced MAPK, SIMK. Plant Cell 2000, 12, 2247–2258. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.L.; Cai, H.; Ji, W.; Luo, X.; Wang, Z.Y.; Wu, J.; Wang, X.D.; Cui, L.; Wang, Y.; Zhu, Y.M.; et al. Overexpression of GsZFP1 enhances salt and drought tolerance in transgenic alfalfa (Medicago sativa L.). Plant Physiol. Biochem. 2013, 71, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y.; Kang, J.; Sun, Y.; Yang, Q.; Wang, P.; Wu, M.; Li, Y.; Long, R.; Qin, Z. Molecular cloning and characterization of a novel gene encoding zinc finger protein from Medicago sativa L. Mol. Biol. Rep. 2009, 36, 2315. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, S.; Borchert, C.; Deyholos, M.; Wang, H.; Brazille, S.; Kawai, K.; Galbraith, D.; Bohnert, H.J. Gene expression profiles during the initial phase of salt stress in rice. Plant Cell 2001, 13, 889–905. [Google Scholar] [CrossRef] [PubMed]
- Lisa, A.L.; Elias, S.; Rahman, M.; Shahid, S.; Iwasaki, T.; Hasan, A.K.M.M.; Kosuge, K.; Fukami, Y.; Seraj, Z. Physiology and gene expression of the rice landrace Horkuch under salt stress. Funct. Plant Biol. 2011, 38, 282–292. [Google Scholar] [CrossRef]
- Bojórquez-Quintal, E.; Velarde-Buendía, A.; Ku-González, Á.; Carillo-Pech, M.; Ortega-Camacho, D.; Echevarría-Machado, I.; Pottosin, I.; Martínez-Estévez, M. Mechanisms of salt tolerance in habanero pepper plants (Capsicum chinense Jacq.): Proline accumulation, ions dynamics and sodium root-shoot partition and compartmentation. Front. Plant Sci. 2014, 5, 605. [Google Scholar] [PubMed]
- Shabala, S. Learning from halophytes: Physiological basis and strategies to improve abiotic stress tolerance in crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence Name | Genbank Number | Primer Sequence | Amplicon Size |
|---|---|---|---|
| Ms-actin2-F | JQ028730.1 | TTCTCACCACACTTCTCGCC | 173 bp |
| Ms-actin2-R | CCAGCCTTCACCATTCCAGT | ||
| Ms-AP2/EREB-F | Not deposited | AATGGGTGGGGAAACGGAAC | 95 bp |
| Ms-AP2/EREB-R | TTTGGTGGTGGAGTGTGGTT | ||
| Ms-NHX1-F | AY513732.1 | GCCATGAAATTCACCGACCG | 118 bp |
| Ms-NHX1-R | CTGCCACCAAAAACAGGACG | ||
| Ms-P5CS-F | X98421.1 | TTTGCGGTCGGAAGGTGTTA | 119 bp |
| Ms-P5CS-R | CGATTTCCAAGGTGCAAGCC | ||
| Ms-ZFN-F | JX131368.1 | CCCAAGCTGCAAGTTTGACC | 154 bp |
| Ms-ZFN-R | TGAGCCCGACTCAACAAGTC | ||
| Ms-SIMKK-F | AJ293274.1 | ACCAGAAGCTCCAACGACTG | 94 bp |
| Ms-SIMKK-R | CCTCGAAGCAGTCCATCTCC | ||
| Ms-RCI2-F | JQ665271.1 | GTTGTCAGGGGCGTCATTCT | 169 bp |
| Ms-RCI2-R | TCCAAGCAGGACAAAACGGA |
| Source of Variation | Height | SER | Salinity Sensitivity Index |
|---|---|---|---|
| Salt (A) | p < 0.05 | p < 0.05 | p < 0.05 |
| Species (B) | p < 0.05 | p < 0.05 | p < 0.05 |
| Dates (C) | p < 0.05 | p < 0.05 | p < 0.05 |
| A × B (Interaction) | p < 0.05 | p < 0.05 | p < 0.05 |
| A × C (Interaction) | p < 0.05 | ns | ns |
| B × C (Interaction) | ns * | p < 0.05 | ns |
| A × B × C (Interaction) | ns | ns | ns |
| K+ | Na+ | ||
|---|---|---|---|
| (mg·g−1 dw) | (mg·g−1 dw) | ||
| M. arborea | Control | 38.5 | 1.1 |
| 50 mM | 24.4 | 10.6 | |
| 50–100 mM | 31.8 | 20.4 | |
| 50–100–150 mM | 13.8 | 21.7 | |
| M. sativa | Control | 22.8 | 3.5 |
| 50–100 mM | 21.5 | 6.9 | |
| 50–100–150 mM | 25.1 | 5.1 | |
| Alborea | Control | 51.2 | 2.3 |
| 50 mM | 38.1 | 19.6 | |
| 50–100 mM | 13.4 | 23.4 |
| Source of Variation | RCI2A | NHX1 | AP2/EREB | SIMKK | ZFN | P5CS_1 |
|---|---|---|---|---|---|---|
| Salt (A) | p < 0.05 | p < 0.05 | p < 0.05 | p ≤ 0.05 | p ≤ 0.05 | p ≤ 0.05 |
| Species (B) | p < 0.05 | p < 0.05 | p < 0.05 | p ≤ 0.05 | p ≤ 0.05 | p ≤ 0.05 |
| Organs (C) | p < 0.05 | p < 0.05 | p < 0.05 | p ≤ 0.05 | p ≤ 0.05 | p ≤ 0.05 |
| A × B (Interaction) | p < 0.05 | p < 0.05 | p < 0.05 | p ≤ 0.05 | ns | p ≤ 0.05 |
| A × C (Interaction) | p < 0.05 | ns * | p < 0.05 | p < 0.05 | p ≤ 0.05 | ns |
| B × C (Interaction) | p < 0.05 | p < 0.05 | p < 0.05 | ns | ns | p ≤ 0.05 |
| A × B × C (Interaction) | p < 0.05 | p < 0.05 | p < 0.05 | p < 0.05 | p ≤ 0.05 | p ≤ 0.05 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tani, E.; Sarri, E.; Goufa, M.; Asimakopoulou, G.; Psychogiou, M.; Bingham, E.; Skaracis, G.N.; Abraham, E.M. Seedling Growth and Transcriptional Responses to Salt Shock and Stress in Medicago sativa L., Medicago arborea L., and Their Hybrid (Alborea). Agronomy 2018, 8, 231. https://doi.org/10.3390/agronomy8100231
Tani E, Sarri E, Goufa M, Asimakopoulou G, Psychogiou M, Bingham E, Skaracis GN, Abraham EM. Seedling Growth and Transcriptional Responses to Salt Shock and Stress in Medicago sativa L., Medicago arborea L., and Their Hybrid (Alborea). Agronomy. 2018; 8(10):231. https://doi.org/10.3390/agronomy8100231
Chicago/Turabian StyleTani, Eleni, Efi Sarri, Maria Goufa, Georgia Asimakopoulou, Maria Psychogiou, Edwin Bingham, George N. Skaracis, and Eleni M. Abraham. 2018. "Seedling Growth and Transcriptional Responses to Salt Shock and Stress in Medicago sativa L., Medicago arborea L., and Their Hybrid (Alborea)" Agronomy 8, no. 10: 231. https://doi.org/10.3390/agronomy8100231