
Eastern Gamagrass Responds Inconsistently to Nitrogen Application in Long-Established Stands and within Diverse Ecotypes

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Long-Established Sites
2.2. Ecotypes Plot
2.3. Statistical Analysis
3. Results
3.1. Long-Established Sites
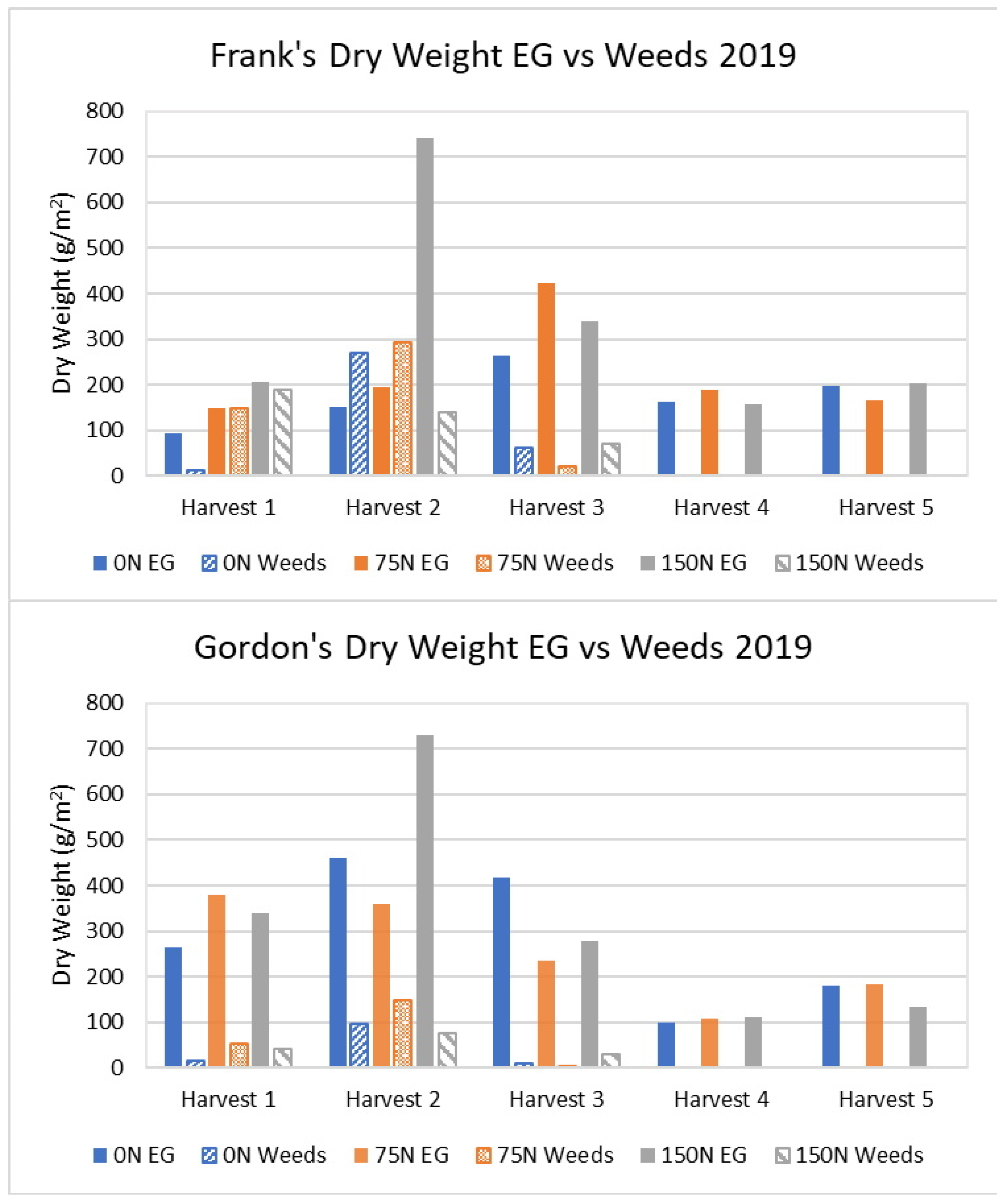
3.1.1. Long-Established Sites’ 2019 Results
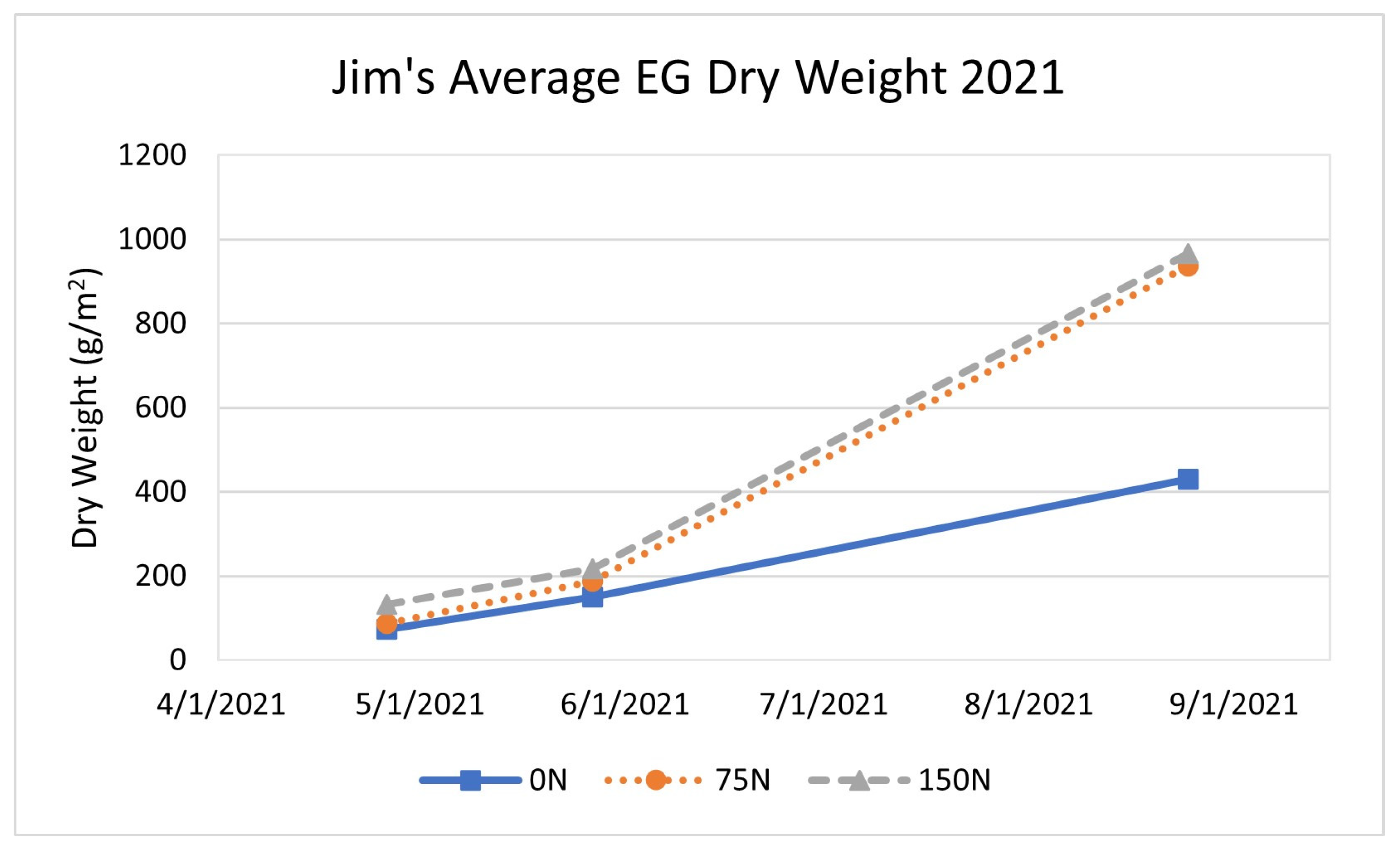
3.1.2. Long-Established Sites 2021 Results
3.2. Ecotypes Plot
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cooney, D.R.; Namoi, N.; Zumpf, C.; Lim, S.-H.; Villamil, M.; Mitchell, R.; Lee, D.K. Biomass Production and Nutrient Removal by Perennial Energy Grasses Produced on a Wet Marginal Land. BioEnergy Res. 2023, 16, 886–897. [Google Scholar] [CrossRef]
- Sainju, U.M.; Allen, B.L. Carbon footprint of perennial bioenergy crop production receiving various nitrogen fertilization rates. Sci. Total Environ. 2023, 861, 160663. [Google Scholar] [CrossRef] [PubMed]
- Waramit, N.; Moore, K.J.; Heggenstaller, A.H. Composition of Native Warm-Season Grasses for Bioenergy Production in Response to Nitrogen Fertilization Rate and Harvest Date. Agron. J. 2011, 103, 655–662. [Google Scholar] [CrossRef]
- Brejda, J.J.; Brown, J.R.; Lorenz, T.E.; Henry, J.; Reid, J.L.; Lowry, S.R. Eastern Gamagrass Responses to Different Harvest Intervals and Nitrogen Rates in Northern Missouri. J. Prod. Agric. 1996, 9, 130–135. [Google Scholar] [CrossRef]
- Douglas, J.L. Effects of Clipping Frequency and N Rates on Yield and Quality of Eastern Gamagrass; Stephen, F., Ed.; Austin State University: Nacogdoches, TX, USA, 1993. [Google Scholar]
- Brakie, M. Yield and Quality of Eastern Gamagrass Selections as Affected by Clipping Interval and Nitrogen Rates; Stephen, F., Ed.; Austin State University ProQuest Dissertations Publishing: Nacogdoches, TX, USA, 1998. [Google Scholar]
- Heggenstaller, A.H.; Moore, K.J.; Liebman, M.; Anex, R.P. Nitrogen Influences Biomass and Nutrient Partitioning by Perennial, Warm-Season Grasses. Agron. J. 2009, 101, 1363–1371. [Google Scholar] [CrossRef]
- Moyer, J.L.; Sweeney, D.W. Responses of eastern gamagrass [Tripsacum dactyloides (L.) L.] forage quality to nitrogen application and harvest system. J. Plant Nutr. 2014, 39, 17–26. [Google Scholar] [CrossRef]
- Coblentz, W.K. Post-Harvest Physiology. In Forages: The Science of Grassland Agriculture; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2020; pp. 721–747. [Google Scholar]
- Brejda, J.J.; Kremer, R.J.; Brown, J.R. Indications of associative nitrogen fixation in eastern gamagrass. Rangel. Ecol. Manag. J. Range Manag. Arch. 1994, 47, 192–195. [Google Scholar] [CrossRef]
- Gillen, R. Production and grazing management for eastern gamagrass. In Proceedings of the 56th Southern Pasture and Forage Crop Improvement Conference, Springdale, AR, USA, 21–22 April 2001. [Google Scholar]
- Swinson, P.J. Establishment of Eastern Gamagrass into a Cool Season Grazing System in Texas Blackland Prairie Soils. In Ag Science and Natural Resources; Texas A&M University-Commerce: Commerce, TX, USA, 2021. [Google Scholar]
- Ahring, R.M.; Frank, H. Establishment of eastern gamagrass from seed and vegetative propagation. J. Range Manag. 1968, 21, 27. [Google Scholar] [CrossRef]
- Springer, T.L.; Dewald, C.L. Eastern gamagrass and other Tripsacum species. In Warm-Season (C4) Grasses; Moser, L.E., Burson, B.L., Sollenberger, L.E., Eds.; American Society of Agronomy, Inc.: Madison, WI, USA; Crop Science Society of America, Inc.: Madison, WI, USA; Soil Science Society of America, Inc.: Madison, WI, USA, 2004. [Google Scholar]
- Gillen, R.L.; Berg, W.A.; Dewald, C.L.; Sims, P.L. Sequence grazing systems on the southern plains. J. Range Manag. 1999, 52, 583. [Google Scholar] [CrossRef]
- Keyser, P.D.; Lituma, C.M.; Bates, G.E.; Holcomb, E.D.; Waller, J.C.; Griffith, A.P. Evaluation of eastern gamagrass and a sorghum × sudangrass for summer pasture. Agron. J. 2020, 112, 1702–1712. [Google Scholar] [CrossRef]
- Burton, G.W.; Southwell, B.L.; Johnson, J.C., Jr. The Palatability of Coastal Bermudagrass (Cynodon dactylon (L.) Pers.) as Influenced by Nitrogen Level and Age. Agron. J. 1956, 48, 360–362. [Google Scholar] [CrossRef]
- Haney, R.L.; Haney, E.B.; Smith, D.R.; Harmel, R.D.; White, M.J. The soil health tool—Theory and initial broad-scale application. Appl. Soil Ecol. 2018, 125, 162–168. [Google Scholar] [CrossRef]
- Kiniry, J.R.; Williams, A.S.; Jacot, J.; Shadow, A.; Brakie, M.; Burson, B.; Jessup, R.; Cordsiemon, R.; Kim, S.; Avila, A.; et al. Diverse eastern gamagrass ecotypes: General characteristics, ploidy levels, and biogeography. Crop. Sci. 2023, 63, 3545–3556. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Latitude, Longitude | Soil Name | Soil Description | Annual Precipitation | ||
|---|---|---|---|---|---|---|
| 2019 | 2020 | 2021 | ||||
| Frank | 29.905733, −96.252083 | Trinity clay | Very-fine, smectitic, thermic Typic Hapluderts | 807 | 865 | 1144 |
| Gordon | 29.898967, −96.236583 | |||||
| Jim | 31.239535, −97.371378 | Heiden clay | Fine, smectitic, thermic Udic Haplusterts | 612 | 796 | 707 |
| Ronnie | 31.027004, −97.245005 | Heiden–Ferris complex | ||||
| Temple | 31.045327, −97.345555 | Houston Black clay | ||||
| Ecotype | Origin Coordinates | Closest City | County | State | Ploidy |
|---|---|---|---|---|---|
| Bellville | 29.910744, −96.250814 | Bellville | Austin | TX | 2x |
| Bryan | 30.743083, −96.497133 | Bryan | Robertson | TX | 3x |
| Jackson | 28.90000, −96.53000 | Edna | Jackson | TX | 4x |
| Roaring Springs | 33.85768, −100.85152 | Roaring Springs | Motley | TX | 4x |
| Stampede | 31.28295, −97.44733 | Stampede | Bell | TX | 2x |
| WNWR | 34.74290, −98.62850 | Meers | Comanche | OK | 2x |
| 0 N | 75 N | 150 N | |||
|---|---|---|---|---|---|
| 2019 | g/m2 | g/m2 | % of 0 N | g/m2 | % of 0 N |
| Frank | 151 | 194 | 128 | 742 | 490 |
| Gordon | 460 | 360 | 78 | 730 | 159 |
| Jim | 413 | 277 | 67 | 604 | 146 |
| Ronnie | 572 | 480 | 84 | 819 | 143 |
| Average | 89 | 149 * | |||
| 2021 | |||||
| Frank | 200 | 196 | 98 | 279 | 142 |
| Gordon | 188 | 188 | 100 | 250 | 133 |
| Jim | 431 | 937 | 224 | 966 | 218 |
| Average | 141 | 164 | |||
| 2019 | 2019 | 2021 | 2022 | First Year | |
|---|---|---|---|---|---|
| Initial Soil | End of Season | Pretreatment | Spring | NO3-N Increase | |
| Frank | |||||
| 0 N | 0.42 | 4.07 | 2.03 | 5.29 | 3.65 |
| 75 N | 2.54 | 7.18 | 4.64 | - | 4.63 |
| 150 N | 1.54 | 4.24 | 2.77 | 1.46 | 2.70 |
| Average | 1.50 | 5.16 | 3.14 | 3.38 | 3.66 |
| Gordon | |||||
| 0 N | 4.38 | 12.28 | 0.82 | 1.11 | 7.90 |
| 75 N | 7.92 | 13.47 | 1.26 | - | 5.56 |
| 150 N | 2.91 | 16.23 | 1.57 | 1.12 | 13.31 |
| Average | 5.07 | 13.99 | 1.22 | 1.11 | 8.92 |
| Jim | |||||
| 0 N | 2.30 | 6.94 | 1.71 | - | 4.63 |
| 75 N | 1.14 | 7.08 | 2.22 | - | 5.94 |
| 150 N | 0.75 | 6.47 | 2.15 | - | 5.72 |
| Average | 1.40 | 6.83 | 2.03 | - | 5.43 |
| Ronnie | |||||
| 0 N | 0.79 | 3.17 | - | - | 2.38 |
| 75 N | 0.47 | 2.07 | - | - | 1.60 |
| 150 N | 0.72 | 3.78 | - | - | 3.06 |
| Average | 0.66 | 3.01 | - | - | 2.35 |
| All Sites | |||||
| 0 N | 1.98 | 6.62 | 1.52 | 3.20 | 4.64 |
| 75 N | 3.02 | 7.45 | 2.70 | - | 4.43 |
| 150 N | 1.48 | 7.68 | 2.16 | 1.29 | 6.20 |
| Average | 2.16 | 7.25 | 2.13 | 2.24 | 5.09 |
| 2020 | 2021 | 2022 | Average | |||||
|---|---|---|---|---|---|---|---|---|
| Dry Wt | 150 N/0 N | Dry Wt | 150 N/0 N | Dry Wt | 150 N/0 N | Dry Wt | 150 N/0 N | |
| Bellville 0 N | 1116 | 1.01 | 997 | 0.91 | 1900 | 0.58 | 1285 | 0.83 |
| Bellville 150 N | 1124 | 912 | 1104 | 1042 | ||||
| Bryan 0 N | 1659 | 1.31 | 2620 | 0.48 | 1834 | 0.94 | 1818 | 0.91 |
| Bryan 150 N | 2174 | 1257 | 1668 | 2251 | ||||
| Jackson 0 N | 1605 | 0.39 | 1878 | 0.70 | 1954 | 1.17 | 1812 | 0.75 |
| Jackson 150 N | 629 | 1307 | 520 | 819 | ||||
| Roaring Springs 0 N | 492 | 3.12 | 1271 | 1.41 | 1181 | 2.27 | 981 | 2.27 |
| Roaring Springs 150 N | 1536 | 1797 | 1667 | |||||
| Stampede 0 N | 421 | 1.60 | 412 | 1.14 | 637 | 0.94 | 490 | 1.23 |
| Stampede 150 N | 676 | 470 | 598 | 581 | ||||
| WNWR 0 N | 736 | 0.94 | 1140 | 0.36 | 467 | 1.82 | 781 | 1.04 |
| WNWR 150 N | 689 | 407 | 851 | 649 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiniry, J.R.; Williams, A.S.; Jacot, J.; Kim, S.; Schantz, M.C. Eastern Gamagrass Responds Inconsistently to Nitrogen Application in Long-Established Stands and within Diverse Ecotypes. Agronomy 2024, 14, 907. https://doi.org/10.3390/agronomy14050907
Kiniry JR, Williams AS, Jacot J, Kim S, Schantz MC. Eastern Gamagrass Responds Inconsistently to Nitrogen Application in Long-Established Stands and within Diverse Ecotypes. Agronomy. 2024; 14(5):907. https://doi.org/10.3390/agronomy14050907
Chicago/Turabian StyleKiniry, James R., Amber S. Williams, Jacqueline Jacot, Sumin Kim, and Merilynn C. Schantz. 2024. "Eastern Gamagrass Responds Inconsistently to Nitrogen Application in Long-Established Stands and within Diverse Ecotypes" Agronomy 14, no. 5: 907. https://doi.org/10.3390/agronomy14050907