
Stem Hydraulic Conductance, Leaf Photosynthesis, and Carbon Metabolism Responses of Cotton to Short-Term Drought and Rewatering

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Measurements and Methods
2.2.1. Soil Water Content
2.2.2. Stem and Leaf Water Potential
2.2.3. Stem Hydraulic Conductance
2.2.4. Photosynthetic Parameters
2.2.5. Chlorophyll Fluorescence Parameters
2.2.6. Non-Structural Carbohydrates Contents
2.2.7. Vulnerability Curve of Xylem Embolism
2.3. Statistical Analysis
3. Results
3.1. The Dynamics of Soil Water Content
3.2. Stem and Leaf Water Potentials
3.3. Recovery of Stem Hydraulic Conductance
3.4. Photosynthetic Responses to Drought Episode and Rewatering
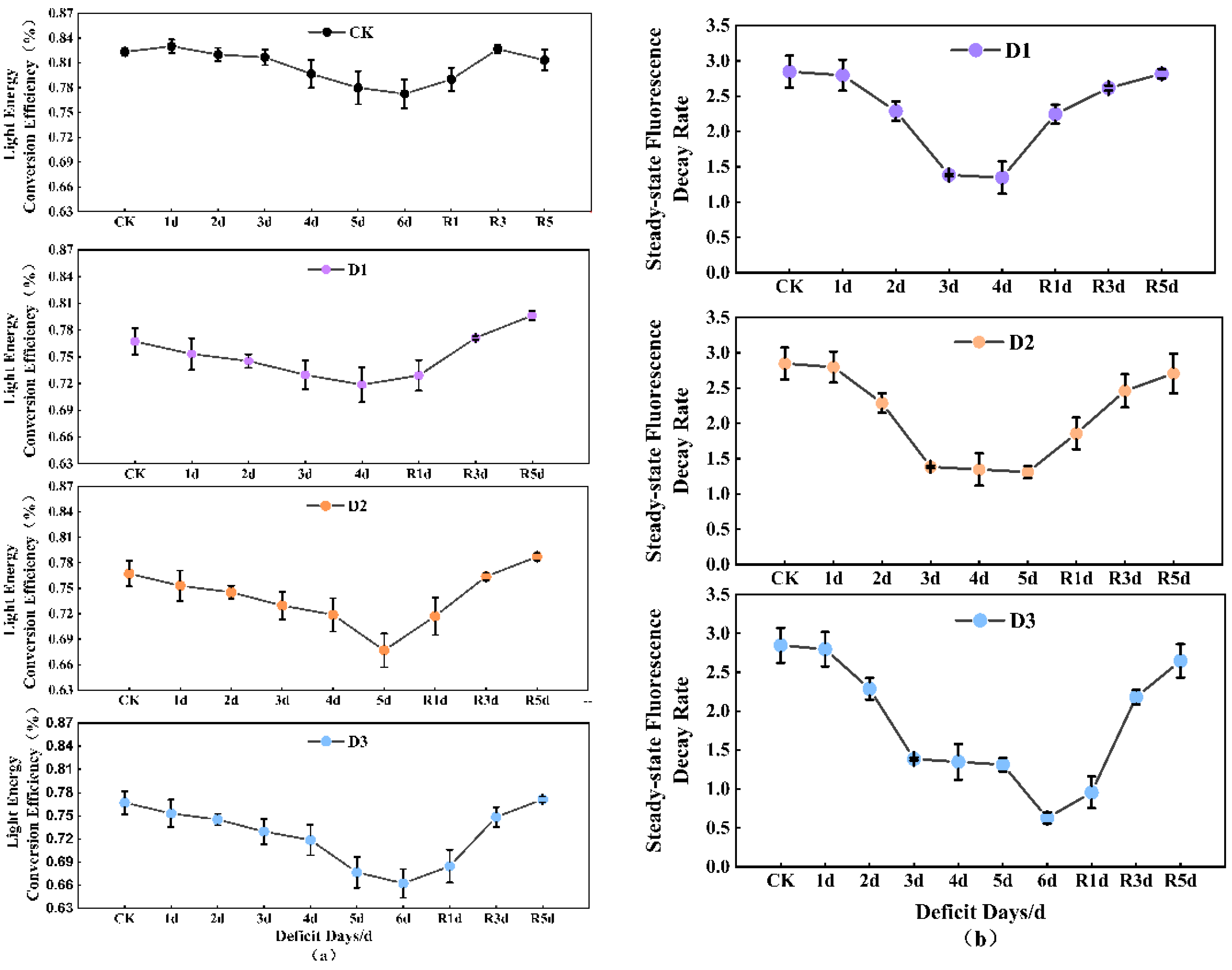
3.5. Chlorophyll Fluorescence Parameters
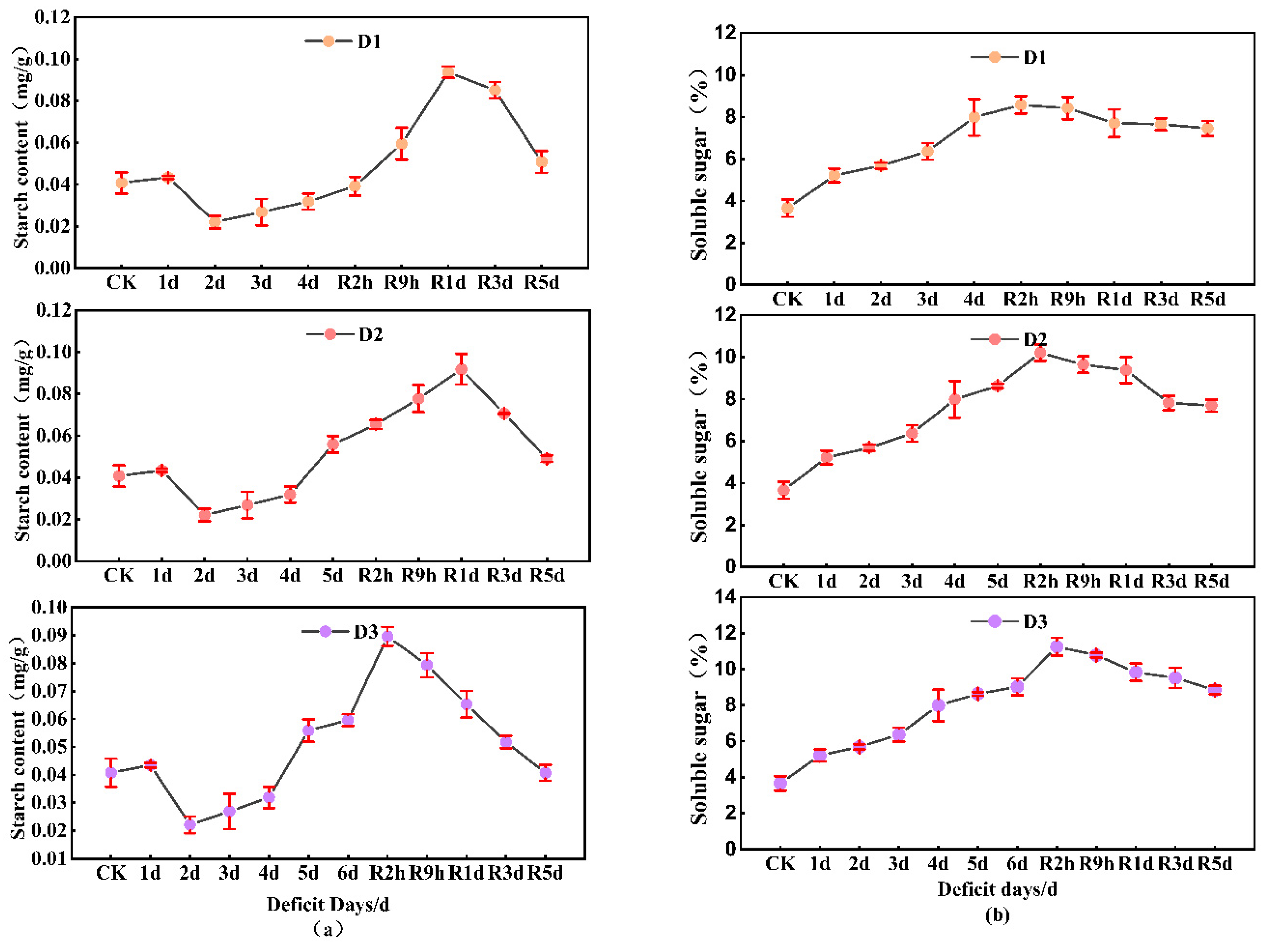
3.6. Non-Structural Carbohydrates
3.7. Stem Xylem Vulnerability Curves
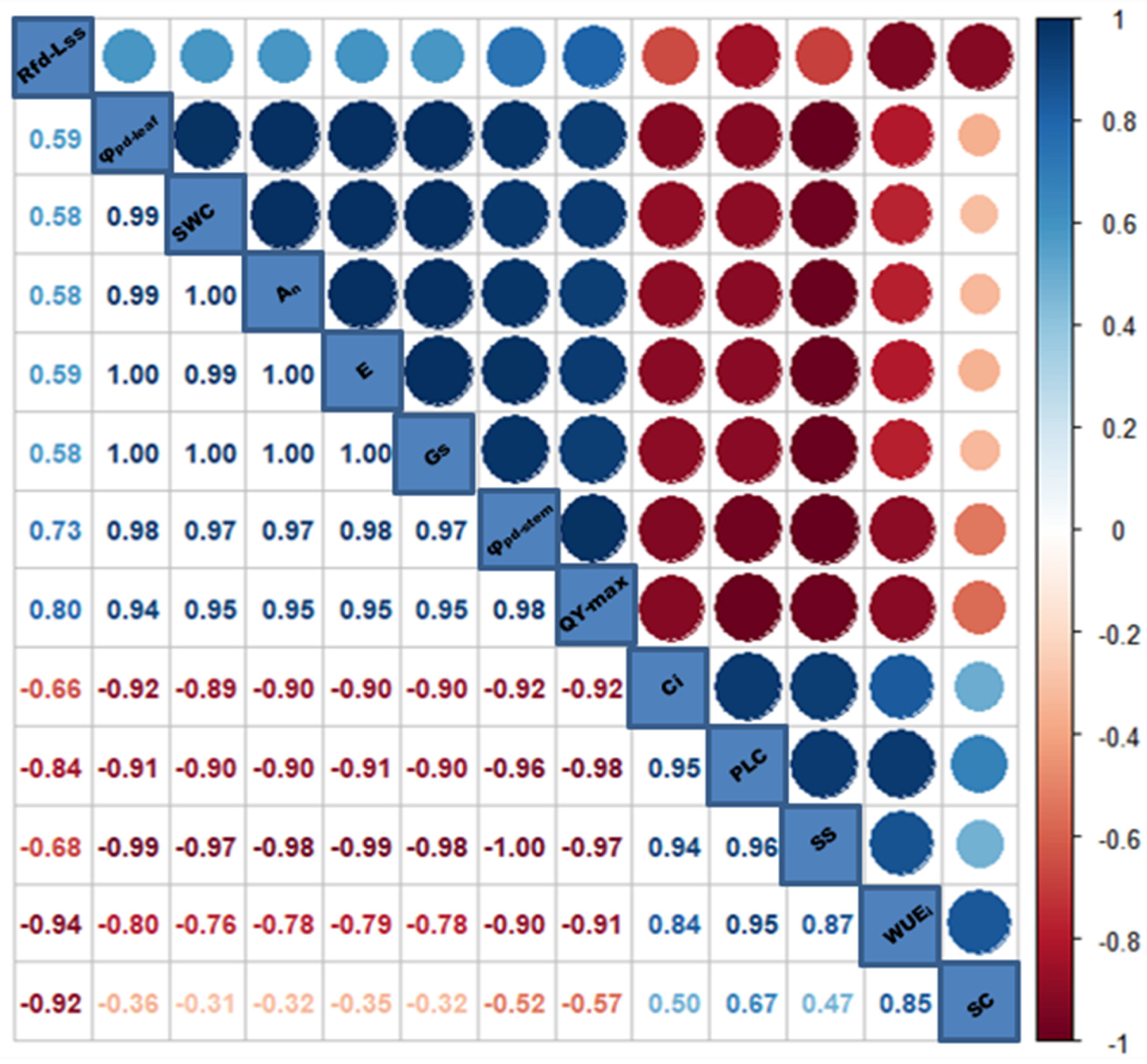
3.8. Analysis of Multiple Functional Traits under Drought–Rewatering Conditions
4. Discussion
4.1. Effect of Drought–Rewatering on Stem Hydraulic Conductance of Cotton
4.2. Effect of Drought–Rewatering on Photosynthesis and Chlorophyll Fluorescence Parameters of Cotton
4.3. Effect of Drought–Rewatering on Non-Structural Carbohydrates of Cotton
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-Wide Reduction in Primary Productivity Caused by the Heat and Drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef] [PubMed]
- AR5 Synthesis Report: Climate Change 2014; IPCC: Geneva, Switzerland, 2015.
- Hsiao, T.C. Plant Responses to Water Stress. Annu. Rev. Plant Physiol. 1973, 24, 519–570. [Google Scholar] [CrossRef]
- Gray, S.B.; Dermody, O.; Klein, S.P.; Locke, A.M.; McGrath, J.M.; Paul, R.E.; Rosenthal, D.M.; Ruiz-Vera, U.M.; Siebers, M.H.; Strellner, R.; et al. Intensifying Drought Eliminates the Expected Benefits of Elevated Carbon Dioxide for Soybean. Nat. Plants 2016, 2, 16132. [Google Scholar] [CrossRef] [PubMed]
- Chao, K.; Chao, J.C.Y. The Development of Cotton Textile Production in China, 1st ed.; Harvard University Asia Center: Cambridge, MA, USA, 1977; Volume 74, ISBN 978-0-674-20021-0. [Google Scholar]
- Klesse, S.; Babst, F.; Lienert, S.; Spahni, R.; Joos, F.; Bouriaud, O.; Carrer, M.; Di Filippo, A.; Poulter, B.; Trotsiuk, V.; et al. A Combined Tree Ring and Vegetation Model Assessment of European Forest Growth Sensitivity to Interannual Climate Variability. Glob. Biogeochem. Cycles 2018, 32, 1226–1240. [Google Scholar] [CrossRef]
- Khajeddin, S.; Matinkhah, S.; Jafari, Z. A Drought Resistance Index to Select Drought Resistant Plant Species Based on Leaf Water Potential Measurements. J. Arid Land 2019, 11, 623–635. [Google Scholar] [CrossRef]
- Reddy, Y.A.N.; Reddy, Y.N.P.; Ramya, V.; Suma, L.S.; Reddy, A.B.N.; Krishna, S.S. Chapter 8—Drought Adaptation: Approaches for Crop Improvement. In Millets and Pseudo Cereals; Singh, M., Sood, S., Eds.; Woodhead Publishing Series in Food Science, Technology and Nutrition; Woodhead Publishing: Sawston, UK, 2021; pp. 143–158. ISBN 978-0-12-820089-6. [Google Scholar]
- Sakoda, K.; Yamori, W.; Groszmann, M.; Evans, J.R. Stomatal, Mesophyll Conductance, and Biochemical Limitations to Photosynthesis during Induction. Plant Physiol. 2021, 185, 146–160. [Google Scholar] [CrossRef]
- Siddique, Z.; Jan, S.; Imadi, S.R.; Gul, A.; Ahmad, P. Drought Stress and Photosynthesis in Plants. In Water Stress and Crop Plants; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2016; pp. 1–11. ISBN 978-1-119-05445-0. [Google Scholar]
- Zhang, Y.J.; Gao, H.; Li, Y.H.; Wang, L.; Kong, D.S.; Guo, Y.Y.; Yan, F.; Wang, Y.W.; Lu, K.; Tian, J.W.; et al. Effect of Water Stress on Photosynthesis, Chlorophyll Fluorescence Parameters and Water Use Efficiency of Common Reed in the Hexi Corridor. Russ. J. Plant Physiol. 2019, 66, 556–563. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, Z.; Han, S.; Jiang, H.; Xu, S.; Zhao, H.; Ren, S. Using the Diurnal Variation Characteristics of Effective Quantum Yield of PSII Photochemistry for Drought Stress Detection in Maize. Ecol. Indic. 2022, 138, 108842. [Google Scholar] [CrossRef]
- Li, S.; Tan, T.; Fan, Y.; Raza, M.A.; Wang, Z.; Wang, B.; Zhang, J.; Tan, X.; Chen, P.; Shafiq, I.; et al. Responses of Leaf Stomatal and Mesophyll Conductance to Abiotic Stress Factors. J. Integr. Agric. 2022, 21, 2787–2804. [Google Scholar] [CrossRef]
- Snider, J.L.; Thangthong, N.; Pilon, C.; Virk, G.; Tishchenko, V. OJIP-Fluorescence Parameters as Rapid Indicators of Cotton (Gossypium Hirsutum L.) Seedling Vigor under Contrasting Growth Temperature Regimes. Plant Physiol. Biochem. 2018, 132, 249–257. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of Plant Survival and Mortality during Drought: Why Do Some Plants Survive While Others Succumb to Drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Nardini, A.; Battistuzzo, M.; Savi, T. Shoot Desiccation and Hydraulic Failure in Temperate Woody Angiosperms during an Extreme Summer Drought. New Phytol. 2013, 200, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Adams, H.D.; Zeppel, M.J.B.; Anderegg, W.R.L.; Hartmann, H.; Landhäusser, S.M.; Tissue, D.T.; Huxman, T.E.; Hudson, P.J.; Franz, T.E.; Allen, C.D.; et al. A Multi-Species Synthesis of Physiological Mechanisms in Drought-Induced Tree Mortality. Nat. Ecol. Evol. 2017, 1, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Tomasella, M.; Casolo, V.; Natale, S.; Petruzzellis, F.; Kofler, W.; Beikircher, B.; Mayr, S.; Nardini, A. Shade-Induced Reduction of Stem Nonstructural Carbohydrates Increases Xylem Vulnerability to Embolism and Impedes Hydraulic Recovery in Populus Nigra. New Phytol. 2021, 231, 108–121. [Google Scholar] [CrossRef] [PubMed]
- Barros, F.D.V.; Bittencourt, P.R.L.; Brum, M.; Restrepo-Coupe, N.; Pereira, L.; Teodoro, G.S.; Saleska, S.R.; Borma, L.S.; Christoffersen, B.O.; Penha, D.; et al. Hydraulic Traits Explain Differential Responses of Amazonian Forests to the 2015 El Niño-Induced Drought. New Phytol. 2019, 223, 1253–1266. [Google Scholar] [CrossRef] [PubMed]
- Ilyas, M.; Nisar, M.; Khan, N.; Hazrat, A.; Khan, A.H.; Hayat, K.; Fahad, S.; Khan, A.; Ullah, A. Drought Tolerance Strategies in Plants: A Mechanistic Approach. J. Plant Growth Regul. 2021, 40, 926–944. [Google Scholar] [CrossRef]
- Sakshaug, E.; Bricaud, A.; Dandonneau, Y.; Falkowski, P.G.; Kiefer, D.A.; Legendre, L.; Morel, A.; Parslow, J.; Takahashi, M. Parameters of Photosynthesis: Definitions, Theory and Interpretation of Results. J. Plankton Res. 1997, 19, 1637–1670. [Google Scholar] [CrossRef]
- Liao, Z.; Zhang, K.; Fan, J.; Li, Z.; Zhang, F.; Xiukang, W.; Wang, H.; Minghui, C.; Zou, Y. Ridge-Furrow Plastic Mulching and Dense Planting with Reduced Nitrogen Improve Soil Hydrothermal Conditions, Rainfed Soybean Yield and Economic Return in a Semi-Humid Drought-Prone Region of China. Soil Tillage Res. 2022, 217, 105291. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Sperry, J.S.; Tyree, M.T. Water-Stress-Induced Xylem Embolism in Three Species of Conifers. Plant Cell Environ. 1990, 13, 427–436. [Google Scholar] [CrossRef]
- Domec, J.-C.; Gartner, B.L. Cavitation and Water Storage Capacity in Bole Xylem Segments of Mature and Young Douglas-Fir Trees. Trees 2001, 15, 204–214. [Google Scholar] [CrossRef]
- Hayat, F.; Ahmed, M.A.; Zarebanadkouki, M.; Javaux, M.; Cai, G.; Carminati, A. Transpiration Reduction in Maize (Zea mays L) in Response to Soil Drying. Front. Plant Sci. 2020, 10, 1695. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Tang, Y.; Zhu, X.-G. Stomata Conductance as a Goalkeeper for Increased Photosynthetic Efficiency. Curr. Opin. Plant Biol. 2022, 70, 102310. [Google Scholar] [CrossRef]
- Hochberg, U.; Windt, C.W.; Ponomarenko, A.; Zhang, Y.-J.; Gersony, J.; Rockwell, F.E.; Holbrook, N.M. Stomatal Closure, Basal Leaf Embolism, and Shedding Protect the Hydraulic Integrity of Grape Stems. Plant Physiol. 2017, 174, 764–775. [Google Scholar] [CrossRef]
- Liu, H.; Ye, Q.; Gleason, S.M.; He, P.; Yin, D. Weak Tradeoff between Xylem Hydraulic Efficiency and Safety: Climatic Seasonality Matters. New Phytol. 2021, 229, 1440–1452. [Google Scholar] [CrossRef]
- Creek, D.; Lamarque, L.J.; Torres-Ruiz, J.M.; Parise, C.; Burlett, R.; Tissue, D.T.; Delzon, S. Xylem Embolism in Leaves Does Not Occur with Open Stomata: Evidence from Direct Observations Using the Optical Visualization Technique. J. Exp. Bot. 2020, 71, 1151–1159. [Google Scholar] [CrossRef]
- Brodribb, T.J.; Holbrook, N.M. Stomatal Closure during Leaf Dehydration, Correlation with Other Leaf Physiological Traits. Plant Physiol. 2003, 132, 2166–2173. [Google Scholar] [CrossRef]
- Hacke, U.G.; Sperry, J.S. Limits to Xylem Refilling under Negative Pressure in Laurus Nobilis and Acer Negundo. Plant Cell Environ. 2003, 26, 303–311. [Google Scholar] [CrossRef]
- Cardoso, A.A.; Batz, T.A.; McAdam, S.A.M. Xylem Embolism Resistance Determines Leaf Mortality during Drought in Persea Americana 1 [OPEN]. Plant Physiol. 2020, 182, 547–554. [Google Scholar] [CrossRef] [PubMed]
- de Campos Siega, T.; Bertoldo, E.; Vismara, L.; Nicareta, C.; Junior, A. Cavitation and Embolism in Plants: Literature Review. Aust. J. Basic Appl. Sci. 2018, 12, 1. [Google Scholar] [CrossRef]
- Ullah, A.; Sun, H.; Yang, X.; Zhang, X. Drought Coping Strategies in Cotton: Increased Crop per Drop. Plant Biotechnol. J. 2017, 15, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Skelton, R.P.; Dawson, T.E.; Thompson, S.E.; Shen, Y.; Weitz, A.P.; Ackerly, D. Low Vulnerability to Xylem Embolism in Leaves and Stems of North American Oaks. Plant Physiol. 2018, 177, 1066–1077. [Google Scholar] [CrossRef] [PubMed]
- de Souza Lorenzoni-Paschoa, L.; Amaral, G.C.; Pezzopane, J.E.M.; Toledo, J.V.; de Abreu, K.M.P.; Xavier, T.M.T.; Cuzzuol, G.R.F. Climate Change and the Ecophysiology of Bertholletia Excelsa Seedlings. Plant Growth Regul. 2022, 98, 155–165. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Escalona, J.M.; Sampol, B.; Medrano, H. Effects of Drought on Photosynthesis in Grapevines under Field Conditions: An Evaluation of Stomatal and Mesophyll Limitations. Funct. Plant Biol. 2002, 29, 461–471. [Google Scholar] [CrossRef]
- Aroca, A.; Gotor, C.; Romero, L.C. Hydrogen Sulfide Signaling in Plants: Emerging Roles of Protein Persulfidation. Front. Plant Sci. 2018, 9, 1369. [Google Scholar] [CrossRef] [PubMed]
- Divo, M.; Cote, C.; de Torres, J.P.; Casanova, C.; Marin, J.M.; Pinto-Plata, V.; Zulueta, J.; Cabrera, C.; Zagaceta, J.; Hunninghake, G.; et al. Comorbidities and Risk of Mortality in Patients with Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2012, 186, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Hetherington, A.M.; Woodward, F.I. The Role of Stomata in Sensing and Driving Environmental Change. Nature 2003, 424, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.M.; Gong, L.; Nie, Z.-F.; Li, F.-P.; Ahammed, G.J.; Fang, X.-W. ABA-Induced Stomatal Movements in Vascular Plants during Dehydration and Rehydration. Environ. Exp. Bot. 2021, 186, 104436. [Google Scholar] [CrossRef]
- Lovisolo, C.; Perrone, I.; Carra, A.; Ferrandino, A.; Flexas, J.; Medrano, H.; Schubert, A. Drought-Induced Changes in Development and Function of Grapevine (Vitis spp.) Organs and in Their Hydraulic and Non-Hydraulic Interactions at the Whole-Plant Level: A Physiological and Molecular Update. Funct. Plant Biol. 2010, 37, 98–116. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, C.; Chen, H.Y.H.; Ruan, H. Response of Plants to Water Stress: A Meta-Analysis. Front. Plant Sci. 2020, 11, 978. [Google Scholar] [CrossRef]
- Zia, R.; Nawaz, M.S.; Siddique, M.J.; Hakim, S.; Imran, A. Plant Survival under Drought Stress: Implications, Adaptive Responses, and Integrated Rhizosphere Management Strategy for Stress Mitigation. Microbiol. Res. 2021, 242, 126626. [Google Scholar] [CrossRef]
- Porcar-Castell, A.; Tyystjärvi, E.; Atherton, J.; van der Tol, C.; Flexas, J.; Pfündel, E.E.; Moreno, J.; Frankenberg, C.; Berry, J.A. Linking Chlorophyll a Fluorescence to Photosynthesis for Remote Sensing Applications: Mechanisms and Challenges. J. Exp. Bot. 2014, 65, 4065–4095. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.-M.; Liu, B.-B.; Wu, Y.; Zou, Z.-R. Interactive Effects of Drought Stresses and Elevated CO2 Concentration on Photochemistry Efficiency of Cucumber Seedlings. J. Integr. Plant Biol. 2008, 50, 1307–1317. [Google Scholar] [CrossRef] [PubMed]
- Cetner, M.D.; Kalaji, H.M.; Goltsev, V.; Aleksandrov, V.; Kowalczyk, K.; Borucki, W.; Jajoo, A. Effects of Nitrogen-Deficiency on Efficiency of Light-Harvesting Apparatus in Radish. Plant Physiol. Biochem. 2017, 119, 81–92. [Google Scholar] [CrossRef]
- Li, J.; Cang, Z.; Jiao, F.; Bai, X.; Zhang, D.; Zhai, R. Influence of Drought Stress on Photosynthetic Characteristics and Protective Enzymes of Potato at Seedling Stage. J. Saudi Soc. Agric. Sci. 2017, 16, 82–88. [Google Scholar] [CrossRef]
- Du, Y.; Zhao, Q.; Chen, L.; Yao, X.; Zhang, W.; Zhang, B.; Xie, F. Effect of Drought Stress on Sugar Metabolism in Leaves and Roots of Soybean Seedlings. Plant Physiol. Biochem. 2020, 146, 1–12. [Google Scholar] [CrossRef]
- Barber, T.M.; Hanson, P.; Kabisch, S.; Pfeiffer, A.F.H.; Weickert, M.O. The Low-Carbohydrate Diet: Short-Term Metabolic Efficacy Versus Longer-Term Limitations. Nutrients 2021, 13, 1187. [Google Scholar] [CrossRef]
- Irène, H.; Florent, P.; Ronan, S.; Maria, P.; Gaëlle, R.; Myriam, D.; Angélique, C.; Marjorie, P.; Marie, B.; Mark, S.; et al. Arabidopsis Plants Acclimate to Water Deficit at Low Cost through Changes of Carbon Usage: An Integrated Perspective Using Growth, Metabolite, Enzyme, and Gene Expression Analysis. Plant Phy. 2010, 154, 357–372. [Google Scholar] [CrossRef]
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; López, R.; Medlyn, B.E. Triggers of Tree Mortality under Drought. Nature 2018, 558, 531–539. [Google Scholar] [CrossRef]
- Sulpice, R.; Pyl, E.-T.; Ishihara, H.; Trenkamp, S.; Steinfath, M.; Witucka-Wall, H.; Gibon, Y.; Usadel, B.; Poree, F.; Piques, M.C.; et al. Starch as a Major Integrator in the Regulation of Plant Growth. Proc. Natl. Acad. Sci. USA 2009, 106, 10348–10353. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, Z.; Zhang, K.; Liao, Z.; Kou, H.; Pei, S.; Dou, Z.; Bai, Z.; Fan, J. Stem Hydraulic Conductance, Leaf Photosynthesis, and Carbon Metabolism Responses of Cotton to Short-Term Drought and Rewatering. Agronomy 2024, 14, 71. https://doi.org/10.3390/agronomy14010071
Lai Z, Zhang K, Liao Z, Kou H, Pei S, Dou Z, Bai Z, Fan J. Stem Hydraulic Conductance, Leaf Photosynthesis, and Carbon Metabolism Responses of Cotton to Short-Term Drought and Rewatering. Agronomy. 2024; 14(1):71. https://doi.org/10.3390/agronomy14010071
Chicago/Turabian StyleLai, Zhenlin, Kaibao Zhang, Zhenqi Liao, Hongtai Kou, Shengzhao Pei, Zhiyao Dou, Zhentao Bai, and Junliang Fan. 2024. "Stem Hydraulic Conductance, Leaf Photosynthesis, and Carbon Metabolism Responses of Cotton to Short-Term Drought and Rewatering" Agronomy 14, no. 1: 71. https://doi.org/10.3390/agronomy14010071