
A Method for Selection of Coffee Varieties Resistant to Fusarium stilboides


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Planting of Coffee Seeds
2.2. Preparation, Purification, and Inoculation of Fungal Isolates
2.3. Harvesting and Quantification of Inoculum
2.4. Formulation of Potting Mixtures, Transplanting and Pathogen Inoculation
2.5. Confirmation of the Pathogen
2.6. Data Collection and Analysis
3. Results
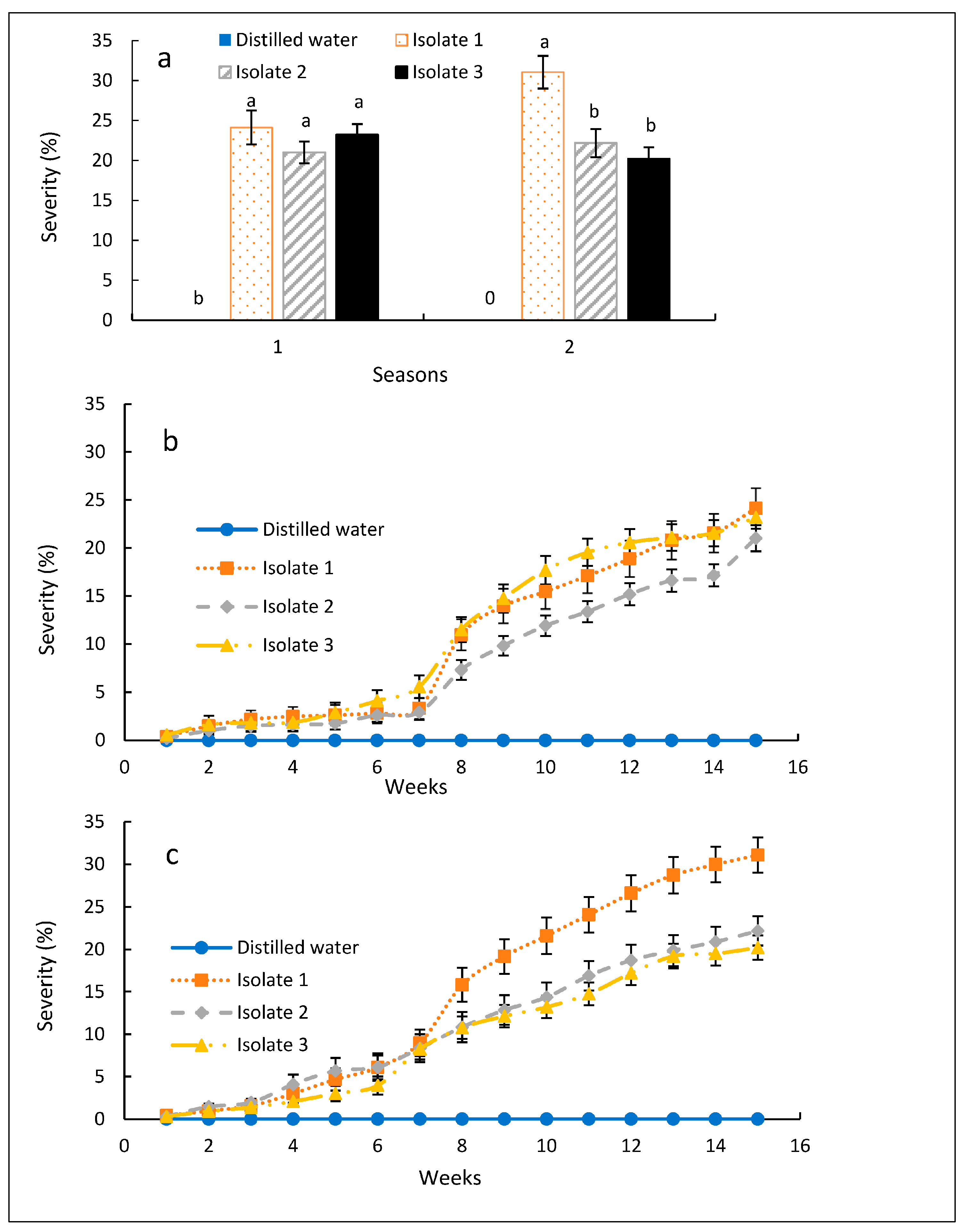
3.1. Pathogenicity of Isolates of Fusarium stilboides Isolated from Infected Coffee Trees
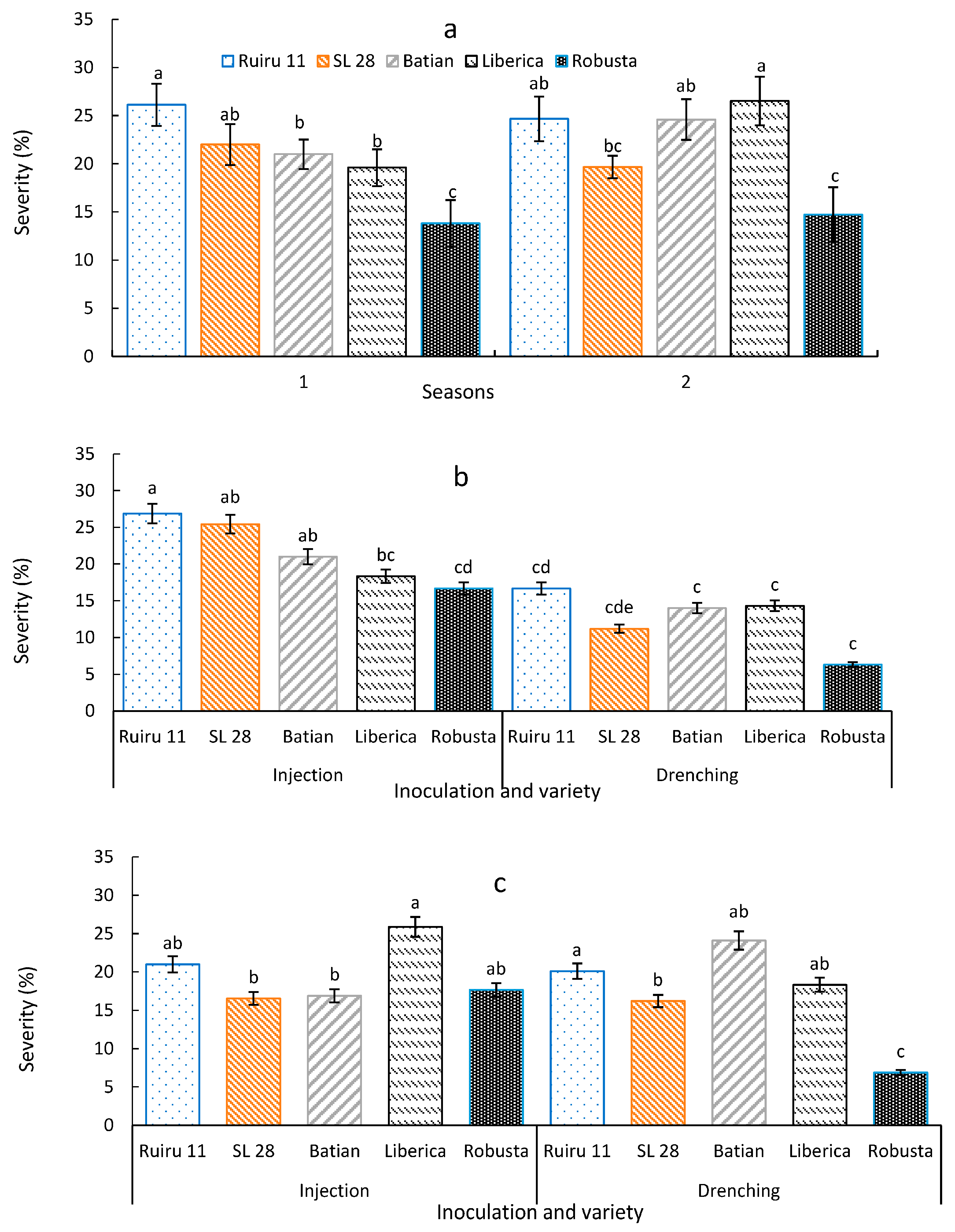
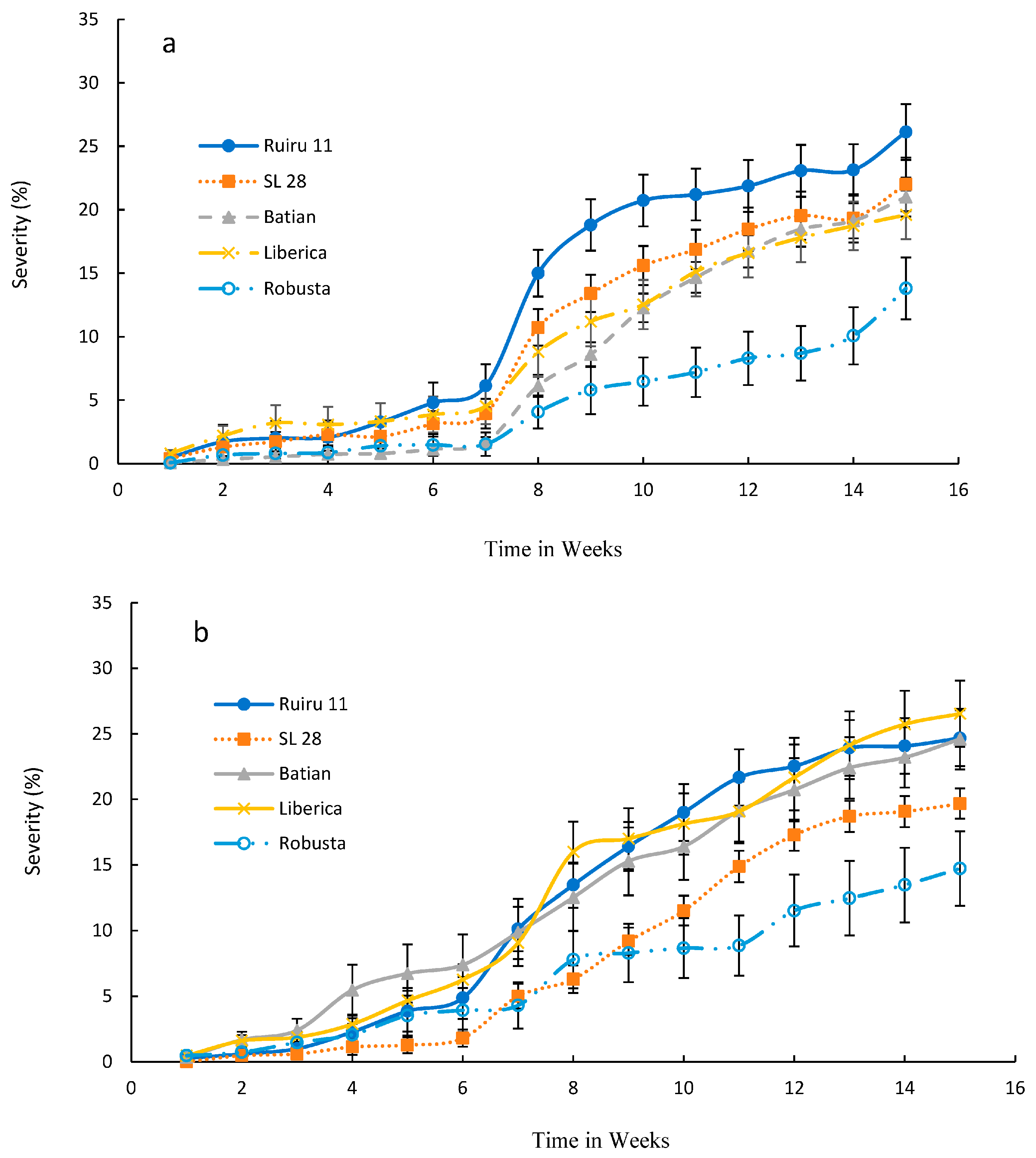
3.2. Effect of Various Isolates of F. stilboides on Disease Severity and Progression
3.3. Severity of Fusarium Bark Disease of Coffee
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Coffee Organization. Coffee Profile: Kenya. 2019. Available online: https://www.ico.org/documents/cy2018-19/icc-124-7e-profile-kenya.pdf (accessed on 26 July 2023).
- Silva, M.C.; Várzea, V.; Guerra-Guimarães, L.; Azinheira, H.G.; Fernandez, D.; Petitot, A.S.; Bertrand, B.; Lashermes, P.; Nicole, M. Coffee resistance to the main diseases: Leaf rust and coffee berry disease. Braz. J. Plant Physiol. 2006, 18, 119–147. [Google Scholar] [CrossRef]
- Serani, S.; Taligoola, H.K.; Hakiza, G.J. An investigation into Fusarium spp. associated with coffee and banana plants as potential pathogens of robusta coffee. Afr. J. Ecol. 2007, 45 (Suppl. S1), 91–95. [Google Scholar] [CrossRef]
- Tshilenge, L.; Kalonji, A.; Tshilenge, P.J. Determination of cultural and biometrical characters of Fusarium species isolated from plant material harvested from coffee (Coffea canephora Pierre.) infected with CWD in Democratic Republic of Congo. Afr. J. Agric. Res. 2010, 5, 3145–3150. [Google Scholar]
- Gichuru, E.K. Fusarium bark disease (FBD). In Compendium of Coffee Diseases and Pests; American Phytopathology Society: Saint Paul, MN, USA, 2016; ISBN 978-0-89054-472-3. [Google Scholar]
- Siddiqi, M.A.; Corbett, D.C.M. Coffee bark diseases in Nyasaland: Pathogenecity, description and identity of the causal organism. Trans. Br. Mycol. Soc. 1963, 46, 91–101. [Google Scholar] [CrossRef]
- Siddiqi, M.A.; Corbett, D.C.M. Coffee bark diseases in Malawi: Properties of the causal organism and conditions favouring the disease. Trans. Br. Mycol. Soc. 1968, 51, 129–135. [Google Scholar] [CrossRef]
- CRI. Coffee Production Recommendations; Minai, J., Ed.; KALRO: Nairobi, Kenya, 2016. [Google Scholar]
- Moura, R.D.; de Castro, L.A.M.; Culik, M.P.; Fernandes, A.A.R.; Fernandes, P.M.B.; Ventura, J.A. Culture medium for improved production of conidia for identification and systematic studies of Fusarium pathogens. J. Microbiol. Methods 2020, 173, 105915. [Google Scholar] [CrossRef] [PubMed]
- Lazarotto, M.; Milanesi, P.M.; Muniz, M.F.B.; Reiniger, L.R.S.; Beltrame, R.; Harakava, R.; Blume, E. Morphological and molecular characterization of Fusarium spp. pathogenic to pecan tree in Brazil. Genet. Mol. Res. 2014, 13, 9390–9402. [Google Scholar] [CrossRef] [PubMed]
- Purwati, R.D.; Hidayah, N. Inoculation methods and conidial densities of Fusarium oxysporum f.sp. cubense in Abaca. Hayati J. Biosciences 2008, 15, 1–7. [Google Scholar] [CrossRef]
- Khan, M.A.; Khan, S.A.; Waheed, U.; Raheel, M.; Khan, Z.; Alrefaei, A.F.; Alkhamis, H.H. Morphological and genetic characterization of Fusarium oxysporum and its management using weed extracts in cotton. J. King Saud Univ.-Sci. 2021, 33, 101299. [Google Scholar] [CrossRef]
- Li, J.; Xu, X.; Ma, Y.; Sun, Q.; Xie, C.; Ma, J. An improved inoculation method to detect wheat and barley genotypes for resistance to Fusarium crown rot. Plant Dis. 2022, 106, 1122–1127. [Google Scholar] [CrossRef] [PubMed]
- Cooke, B.M.; Gareth, J.D.; Kaye, B. The Epidemiology of Plant Diseases, 2nd ed.; Cooke, B.M., Gareth Jones, D., Kaye, B., Eds.; Springer: Dordrecht, The Netherlands, 2007; 576p. [Google Scholar]
- Simko, I.; Piepho, H.-P. The area under the disease progress stairs: Calculation, advantage, and application. Phytopathology 2012, 102, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Omondi, C.O.; Hindorf, H.; Welz, H.G.; Saucke, D.; Ayiecho, P.O.; Mwangombe, A.W. Genetic diversity among isolates of Colletotrichum kahawae causing Coffee Berry Disease. In Proceedings of the 17th International Conference on Coffee Science (ASIC), Nairobi, Kenya, 20–15 July 1997; pp. 800–803. [Google Scholar]
- Mphahlele, G.H.; Kena, M.A.; Manyevere, A. Evaluation of aggressiveness of Alternaria solani isolates to commercial tomato cultivars. Arch. Phytopathol. Plant Prot. 2020, 53, 570–580. [Google Scholar] [CrossRef]
- Storey, H.H. A bark disease of Coffee in East Africa. Ann. Appl. Biol. 1932, 19, 173–184. [Google Scholar] [CrossRef]
- Gimase, J.M.; Thagana, W.M.; Omondi, C.O.; Cheserek, J.J.; Gichuru, E.K. Genetic relationship and the occurrence of multiple gene resistance to coffee berry disease (Colletotrichum kahawae, Waller Bridge) within selected Coffea arabica varieties in Kenya. Afr. J. Plant Sci. 2021, 15, 39–48. [Google Scholar] [CrossRef]
- Zhang, X.; Peck, L.D.; Flood, J.; Ryan, M.J.; Barraclough, T.G. Temperature contributes to host specialization of coffee wilt disease (Fusarium xylarioides) on arabica and robusta coffee crops. Sci. Rep. 2023, 13, 9327. [Google Scholar] [CrossRef] [PubMed]
- Fick, S.; Hijmans, R. New 1 km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2021, 37, 4302–4315. [Google Scholar] [CrossRef]
- Voss, H.H.; Bowden, R.L.; Leslie, J.F.; Miedaner, T. Variation and transgression of aggressiveness among two Gibberella zeae crosses developed from highly aggressive parental isolates. Phytopathology 2010, 100, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Vanderplank, J.E. Disease Resistance in Plants; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Girma, A. Diversity in pathogenicity and genetics of Gibberella xylarioides (Fusarium xylarioides) populations and resistance of Coffea spp. in Ethiopia. J. Plant Dis. Prot. 2004, 112, 134–145. [Google Scholar]
- Gong, X.; He, X.; Zhang, Y.; Li, L.; Sun, Z.; Bai, G.; Singh, P.K.; Li, T. Development of an evaluation system for Fusarium resistance in wheat grains and its application in assessment of the corresponding effects of Fhb1. Plant Dis. 2020, 104, 2210–2216. [Google Scholar] [CrossRef]
- Jeger, M.J.; Viljanen-Rollinson, S.L.H. The use of the area under the disease-progress curve (AUDPC) to assess quantitative disease resistance in crop cultivars. Theor. Appl. Genet. 2001, 102, 32–40. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Score | 0 (0%) | 1 (<25% Infection at Soil Level) | 2 (>25%–<50% Infections from Soil Level Upwards) | 3 (>50%–<75% Infection from Soil Level Upwards) | 4 (>75% Infection) |
|---|---|---|---|---|---|
| Description | Healthy plants, turgid and shiny stems and leaves. No visible lesion at the soil level. | Small visible brown water-soaked lesions at the soil level. Flaccid leaves and stem. Note: There may be wilting but no visible lesion at the soil level. | Dark brown/black lesions at the soil level extending upwards and wilting. Note: There may be extensive wilting and stem drooping even without visible lesions at the soil level upwards. | ExtendedExtened dark brown/black lesions from soil level upwards. Wilting, drying, and yellowing of leaves and stem drooping. | Dead, dried, and rotten plants. Extended dark brown/black lesions colonizing more than 75% of the stem area or the entire plant. |
| Visual aid |  |  |  |  |  |
| Score | 0 (0%) Stem Colonization | 1 (<10% Stem Colonization) | 2 (>10%–<30% Stem Colonization) | 3 (>30%–<50% Stem Colonization) | 4 (>50% Stem Colonization) |
|---|---|---|---|---|---|
| Description | Healthy plants/scab (healing injection point)-(no visible lesion even under magnifying lens). Turgid and shiny stems and leaves. | Turgid and shiny stems and leaves with tiny brown/black lesions at the injection point. Spores may be visible using a magnifying lens. Note: There may be no visible lesions at the point of injection, but the stems and Leaves are flaccid but not drooping. Check for lesions along the stem or at soil level. | Extended dark brown/black lesions at the injection point, either extending upwards or downwards. There may be extended lesions from the soil level upwards and wilted leaves. Spores may be visible using a magnifying lens. Drooping and/or drying stem above the lesion and wilted leaves. | Extended dark brown/black lesions at the injection point, either extending upwards or downwards. There may be extended lesions from the soil level upwards and wilted leaves. Spores may be visible using a magnifying lens. Drooping and/or drying stem above the lesion and wilted leaves. | Extended dark brown/black lesions at the injection point, either extending upwards or downwards. There may be extended lesions from soil level upwards and wilted leaves. Spores may be visible using a magnifying lens. Drooping and/or drying stem above the lesion and wilted leaves. |
| Visual aid |  |  |  |  |  |
| Season One | |||||
|---|---|---|---|---|---|
| Source | df | SS | MS | F Value | Pr > F |
| Inoculation method | 1 | 5029.45 | 5029.45 | 29.83 | <0.0001 * |
| Inoculum concentration | 3 | 15,550.01 | 5183.34 | 30.74 | <0.0001 * |
| Pathogen isolate | 2 | 458.79 | 229.39 | 1.15 | 0.3185 |
| Variety | 4 | 3320.34 | 830.09 | 4.16 | 0.0028 * |
| Variety × inoculation method | 4 | 713.07 | 178.27 | 1.06 | 0.3782 |
| Variety × inoculum concentration | 8 | 395.14 | 49.39 | 0.25 | 0.9811 |
| Variety × pathogen isolate | 8 | 1812.03 | 226.5 | 1.13 | 0.3403 |
| Inoculation method × inoculum concentration | 3 | 1500.23 | 500.08 | 2.97 | 0.0326 |
| Pathogen isolate × inoculum concentration | 4 | 1981.39 | 495.35 | 2.48 | 0.0444 * |
| Variety × inoculation method × inoculum concentration | 12 | 2150.79 | 179.23 | 1.06 | 0.392 |
| Variety × pathogen isolate × inoculum concentration | 16 | 3740.68 | 233.79 | 1.17 | 0.2915 |
| Season Two | |||||
| Inoculation method | 1 | 365.07 | 365.07 | 2.74 | 0.0664 |
| Inoculum concentration | 3 | 17,210.01 | 5736.67 | 24.01 | <0.0001 * |
| Pathogen isolate | 2 | 6013.16 | 3006.58 | 13.46 | <0.0001 * |
| Variety | 4 | 3849.08 | 962.27 | 4.31 | 0.0022 * |
| Variety × inoculation method | 4 | 2350.63 | 587.66 | 2.46 | 0.0461 * |
| Variety × inoculum concentration | 8 | 2504.41 | 313.05 | 1.4 | 0.1962 |
| Variety × pathogen isolate | 8 | 2277.21 | 284.65 | 1.27 | 0.2573 |
| Inoculation method × inoculum concentration | 3 | 265.37 | 88.46 | 0.37 | 0.7746 |
| Pathogen isolate × inoculum concentration | 4 | 800.36 | 200.09 | 0.9 | 0.4671 |
| Variety × inoculation method × inoculum concentration | 12 | 2040.14 | 170.01 | 0.71 | 0.7399 |
| Variety × pathogen isolate × inoculum concentration | 16 | 4358.16 | 272.39 | 1.22 | 0.2532 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alworah, G.O.; Ogendo, J.O.; Mafurah, J.J.; Gichuru, E.K.; Miano, D.W.; Okumu, O.O. A Method for Selection of Coffee Varieties Resistant to Fusarium stilboides. Agronomy 2023, 13, 2321. https://doi.org/10.3390/agronomy13092321
Alworah GO, Ogendo JO, Mafurah JJ, Gichuru EK, Miano DW, Okumu OO. A Method for Selection of Coffee Varieties Resistant to Fusarium stilboides. Agronomy. 2023; 13(9):2321. https://doi.org/10.3390/agronomy13092321
Chicago/Turabian StyleAlworah, Getrude Okutoyi, Joshua Ondura Ogendo, Joseph Juma Mafurah, Elijah Kathurima Gichuru, Douglas Watuku Miano, and Otieno Oliver Okumu. 2023. "A Method for Selection of Coffee Varieties Resistant to Fusarium stilboides" Agronomy 13, no. 9: 2321. https://doi.org/10.3390/agronomy13092321