
Combined Effects of Straw Return with Nitrogen Fertilizer on Leaf Ion Balance, Photosynthetic Capacity, and Rice Yield in Saline-Sodic Paddy Fields
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Experimental Design
2.3. Sampling Methods and Measurements
2.3.1. Determination of Na+ and K+ Content in Rice Leaves
2.3.2. Determination of Malondialdehyde (MDA) Content and Relative Electrolyte
Leakage (REL), Superoxide (SOD) and Peroxidase (POD) Activities in Rice Leaves
2.3.3. Determination of Leaf Relative Water Content (RWC) and Water Potential (Ψw)
2.3.4. Determination of Photosynthetic Parameters and SPAD Value of Rice Leaves
2.3.5. Determination of Rice Yield
2.3.6. Statistical Analysis
3. Results
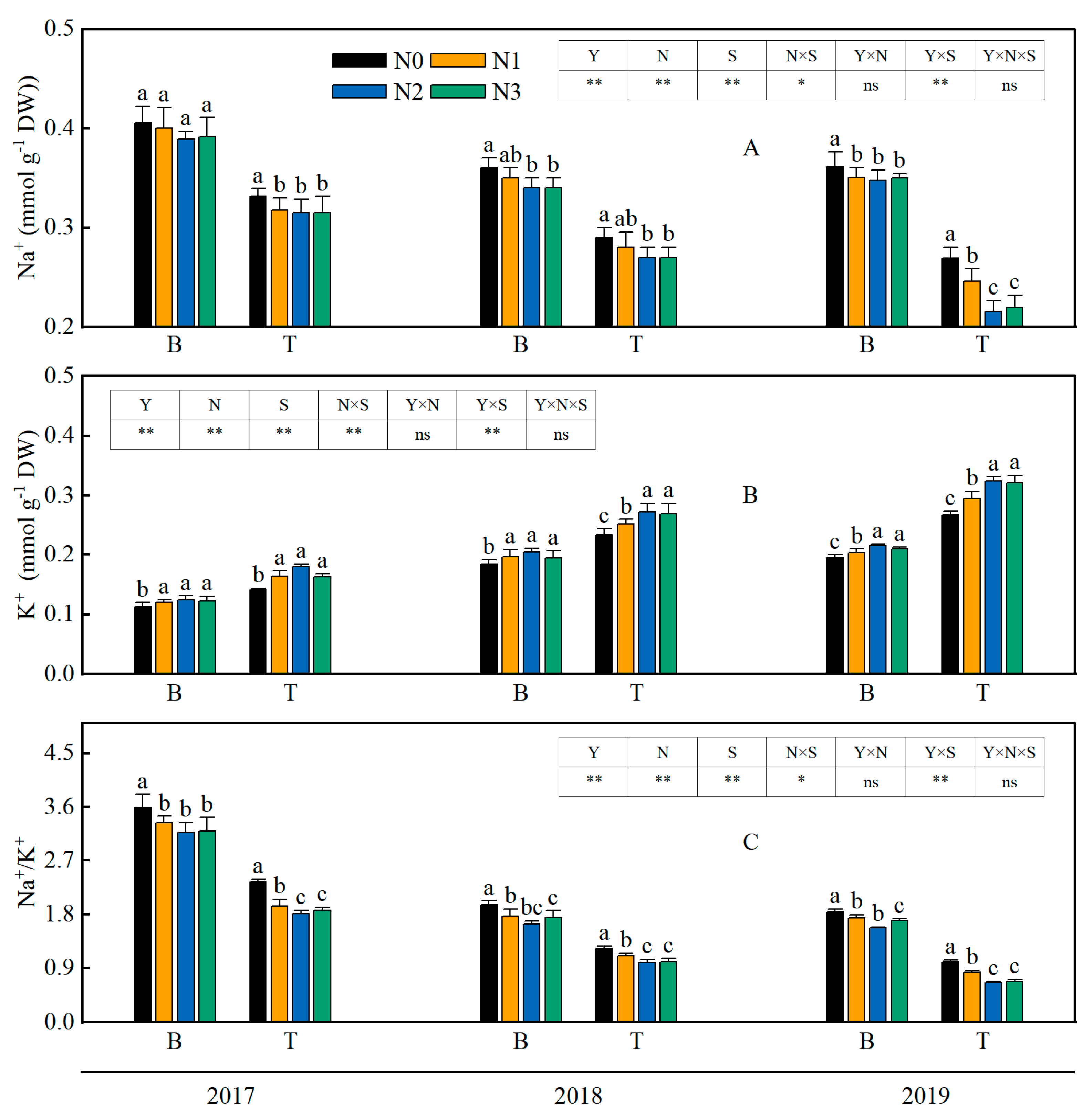
3.1. Na+ and K+ Contents and the Na+/K+ Ratio of Rice Leaves
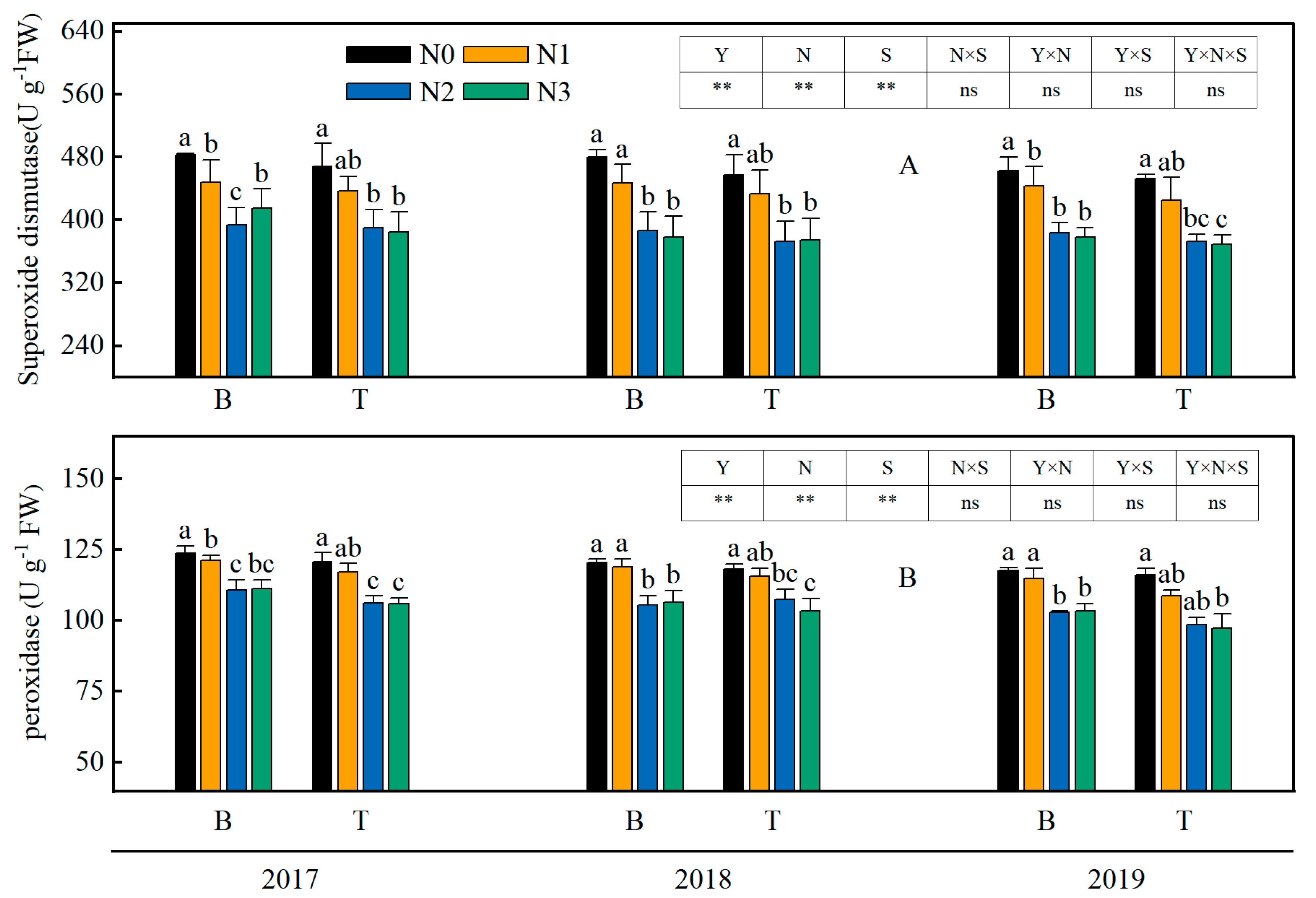
3.2. Malondialdehyde (MDA) and Relative Electricity Leakage (REL) of Rice Leaves
3.3. Superoxide Dismutase (SOD) and Peroxidase (POD) Activities of Rice Leaves
3.4. Relative Water Content (RWC) and Leaf Water Potential (Ψw) of Rice Leaves
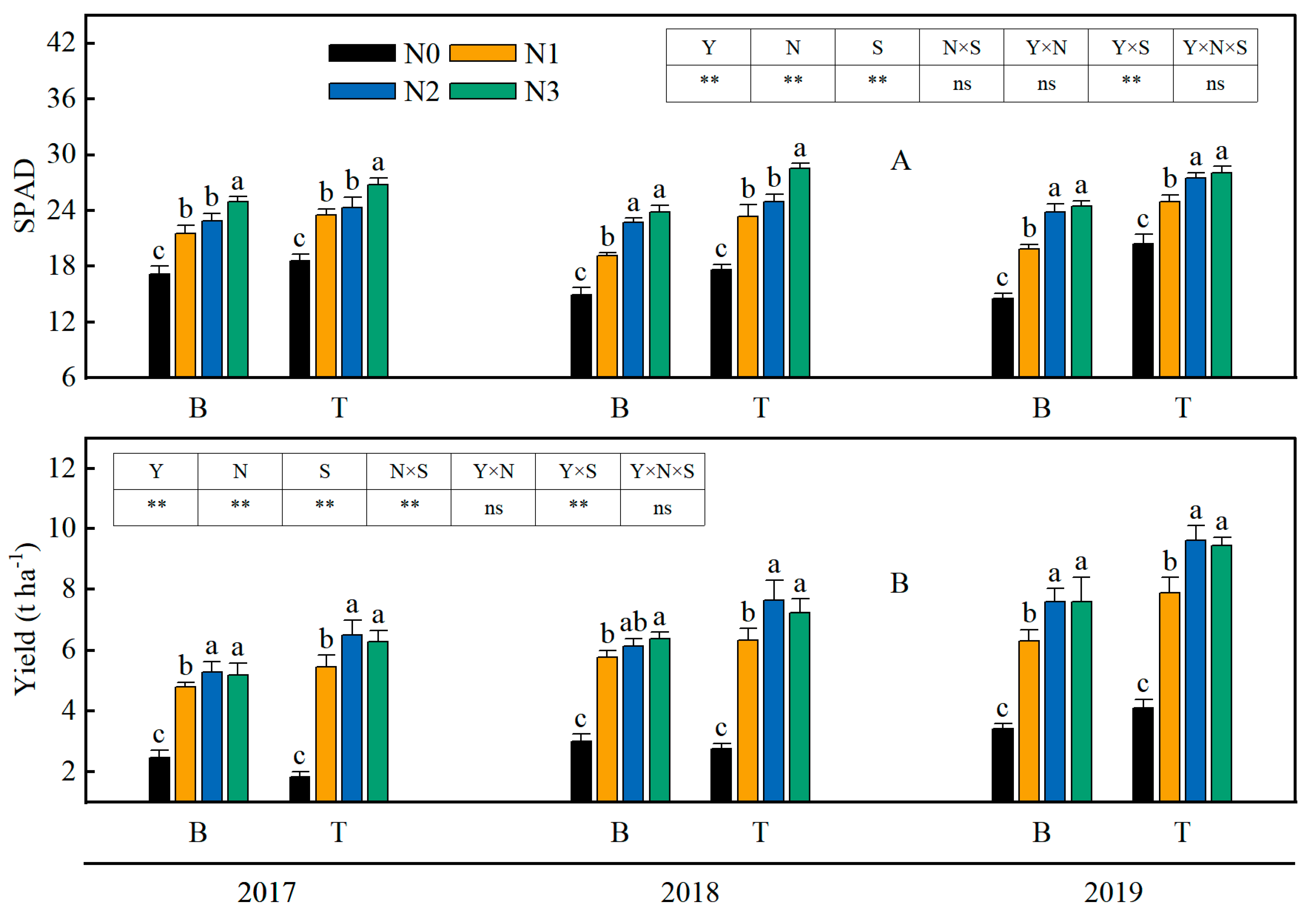
3.5. SPAD Value of Rice Leaves
3.6. Net Photosynthetic Rate (A), Stomatal Conductance (gs), and Transpiration Rate (E) of Rice
3.7. Grain Yield of Rice
4. Discussion
4.1. The Application of Both Straw and Nitrogen Fertilizer Effectively Maintained the Ion Balance of Rice Tissue in Saline-Sodic Land
4.2. The Combined Application of Straw and Nitrogen Fertilizer Enhanced the Oxidative Damage and Water Status of Rice Leaves in Saline-Sodic Land
4.3. The Application of Straw and Nitrogen Fertilizer in Combination Resulted in an Increase in the SPAD Value and Photosynthetic Capacity of Rice Leaves in Saline-Sodic Land
4.4. Combined Application of Straw and Nitrogen Fertilizer Improved the Rice Yield in Saline-Sodic Land
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mostofa, M.G.; Hossain, M.A.; Fujita, M. Trehalose pretreatment induces salt tolerance in rice (Oryza sativa L.) seedlings: Oxidative damage and co-induction of antioxidant defense and glyoxalase systems. Protoplasma 2015, 252, 461–475. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; An, F.; Ma, H.; Wang, Z.; Zhou, X.; Liu, Z. Variations on Soil Salinity and Sodicity and Its Driving Factors Analysis under Microtopography in Different Hydrological Conditions. Water 2016, 8, 227. [Google Scholar] [CrossRef]
- Venkatalaxmi, A.; Padmavathi, B.S.; Amaranath, T. A general solution of unsteady Stokes equations. Fluid Dyn. Res. 2004, 35, 229–236. [Google Scholar] [CrossRef]
- Wang, L.; Seki, K.; Miyazaki, T.; Ishihama, Y. The causes of soil alkalinization in the Songnen Plain of Northeast China. Paddy Water Environ. 2009, 7, 259–270. [Google Scholar] [CrossRef]
- Chi, C.M.; Zhao, C.W.; Sun, X.J.; Wang, Z.C. Reclamation of saline-sodic soil properties and improvement of rice (Oriza sativa L.) growth and yield using desulfurized gypsum in the west of Songnen Plain, northeast China. Geoderma 2012, 187–188, 24–30. [Google Scholar] [CrossRef]
- Roy, S.J.; Negrão, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef]
- Ruiz, K.B.; Biondi, S.; Martínez, E.A.; Orsini, F.; Antognoni, F.; Jacobsen, S.E. Quinoa—A Model Crop for Understanding Salt-tolerance Mechanisms in Halophytes. Plant Biosyst.—Int. J. Deal. All Asp. Plant Biol. 2015, 150, 357–371. [Google Scholar] [CrossRef]
- Liu, C.; Mao, B.; Yuan, D.; Chu, C.; Duan, M. Salt tolerance in rice: Physiological responses and molecular mechanisms. Crop J. 2022, 10, 13–25. [Google Scholar] [CrossRef]
- Jini, D.; Joseph, B. Physiological Mechanism of Salicylic Acid for Alleviation of Salt Stress in Rice. Rice Sci. 2017, 24, 97–108. [Google Scholar] [CrossRef]
- Feng, N.; Yu, M.; Li, Y.; Jin, D.; Zheng, D. Prohexadione-calcium alleviates saline-alkali stress in soybean seedlings by improving the photosynthesis and up-regulating antioxidant defense. Ecotoxicol. Environ. Saf. 2021, 220, 112369. [Google Scholar] [CrossRef]
- Wang, Y.; Yan, J.; Goult, B.T. Force-Dependent Binding Constants. Biochemistry 2019, 58, 4696–4709. [Google Scholar] [CrossRef] [PubMed]
- Ahanger, M.A.; Qin, C.; Begum, N.; Maodong, Q.; Dong, X.X.; El-Esawi, M.; El-Sheikh, M.A.; Alatar, A.A.; Zhang, L. Nitrogen availability prevents oxidative effects of salinity on wheat growth and photosynthesis by up-regulating the antioxidants and osmolytes metabolism, and secondary metabolite accumulation. BMC Plant Biol. 2019, 19, 479. [Google Scholar] [CrossRef]
- Iqbal, N.; Umar, S.; Khan, N.A. Nitrogen availability regulates proline and ethylene production and alleviates salinity stress in mustard (Brassica juncea). J. Plant Physiol. 2015, 178, 84–91. [Google Scholar] [CrossRef]
- Cong, R.; Liu, T.; Lu, P.; Ren, T.; Li, X.; Lu, J. Nitrogen fertilization compensation the weak photosynthesis of Oilseed rape (Brassca napus L.) under haze weather. Sci. Rep. 2020, 10, 4047. [Google Scholar] [CrossRef] [PubMed]
- Akram, M.; Ashraf, M.Y.; Jamil, M.; Iqbal, R.M.; Nafees, M.; Khan, M.A. Nitrogen application improves gas exchange characteristics and chlorophyll fluorescence in maize hybrids under salinity conditions. Russ. J. Plant Physiol. 2011, 58, 394–401. [Google Scholar] [CrossRef]
- Ran, C.; Gulaqa, A.; Zhu, J.; Wang, X.; Zhang, S.; Geng, Y.; Guo, L.; Jin, F.; Shao, X. Benefits of Biochar for Improving Ion Contents, Cell Membrane Permeability, Leaf Water Status and Yield of Rice Under Saline–Sodic Paddy Field Condition. J. Plant Growth Regul. 2019, 39, 370–377. [Google Scholar] [CrossRef]
- Elgharably, A.; Marschner, P.; Rengasamy, P. Wheat growth in a saline sandy loam soil as affected by N form and application rate. Plant Soil 2009, 328, 303–312. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Hou, Z.; Wu, L.; Liang, Y.; Wei, C. Effects of salinity and nitrogen on cotton growth in arid environment. Plant Soil 2009, 326, 61–73. [Google Scholar] [CrossRef]
- Ahmed, M.; Rauf, M.; Mukhtar, Z.; Saeed, N.A. Excessive use of nitrogenous fertilizers: An unawareness causing serious threats to environment and human health. Environ. Sci. Pollut Res. Int. 2017, 24, 26983–26987. [Google Scholar] [CrossRef]
- Sanchez, P.A. Soil Fertility and Hunger in Africa. Science 2002, 295, 2019–2020. [Google Scholar] [CrossRef]
- Xiao, Q.; Zhu, L.-X.; Zhang, H.-P.; Li, X.-Y.; Shen, Y.-F.; Li, S.-Q. Soil amendment with biochar increases maize yields in a semi-arid region by improving soil quality and root growth. Crop Pasture Sci. 2016, 67, 495–507. [Google Scholar] [CrossRef]
- Ju, X.-T.; Xing, G.-X.; Chen, X.-P.; Zhang, S.-L.; Zha, L.-J.; Liu, X.-J.; Cui, Z.-L.; Yin, B.; Christie, P.; Zhu, Z.-L.; et al. Reducing environmental risk by improving N management in intensive Chinese agricultural systems. Proc. Natl. Acad. Sci. USA 2009, 106, 3041–3046, Erratum in Proc. Natl. Acad. Sci. USA 2009, 106, 8077–8077. [Google Scholar] [CrossRef]
- Yin, H.; Zhao, W.; Li, T.; Cheng, X.; Liu, Q. Balancing straw returning and chemical fertilizers in China: Role of straw nutrient resources. Renew. Sustain. Energy Rev. 2018, 81, 2695–2702. [Google Scholar] [CrossRef]
- Li, Y.; Wang, J.; Pan, F.; Chapman, S.J.; Yao, H. Soil nitrogen availability alters rhizodeposition carbon flux into the soil microbial community. J. Soils Sediments 2016, 16, 1472–1480. [Google Scholar] [CrossRef]
- Sui, N.; Zhou, Z.; Yu, C.; Liu, R.; Yang, C.; Zhang, F.; Song, G.; Meng, Y. Yield and potassium use efficiency of cotton with wheat straw incorporation and potassium fertilization on soils with various conditions in the wheat–cotton rotation system. Field Crops Res. 2015, 172, 132–144. [Google Scholar] [CrossRef]
- Wang, Z.; Ma, L.J.; Khattak, W.A.; Hu, W.; Meng, Y.L.; Zhou, Z.G. Combined effects of nitrogen fertilizer and straw application on aggregate distribution and aggregate-associated organic carbon stability in an alkaline sandy loam soil. Eur. J. Soil Sci. 2018, 69, 1105–1116. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, Y.; Wang, J.; Pang, H.; Li, Y. Buried straw layer plus plastic mulching reduces soil salinity and increases sunflower yield in saline soils. Soil Tillage Res. 2016, 155, 363–370. [Google Scholar] [CrossRef]
- Saroa, G.S.; Lal, R. Soil restorative effects of mulching on aggregation and carbon sequestration in a Miamian soil in central Ohio. Land Degrad. Dev. 2003, 14, 481–493. [Google Scholar] [CrossRef]
- Zhao, W.; Zhou, Q.; Tian, Z.; Cui, Y.; Liang, Y.; Wang, H. Apply biochar to ameliorate soda saline-alkali land, improve soil function and increase corn nutrient availability in the Songnen Plain. Sci. Total Environ. 2020, 722, 137428. [Google Scholar] [CrossRef] [PubMed]
- Brennan, J.; Hackett, R.; McCabe, T.; Grant, J.; Fortune, R.A.; Forristal, P.D. The effect of tillage system and residue management on grain yield and nitrogen use efficiency in winter wheat in a cool Atlantic climate. Eur. J. Agron. 2014, 54, 61–69. [Google Scholar] [CrossRef]
- Li, H.; Wang, L.; Peng, Y.; Zhang, S.; Lv, S.; Li, J.; Abdo, A.I.; Zhou, C.; Wang, L. Film mulching, residue retention and N fertilization affect ammonia volatilization through soil labile N and C pools. Agric. Ecosyst. Environ. 2021, 308, 107272. [Google Scholar] [CrossRef]
- Singh, Y.; Singh, B.; Ladha, J.K.; Khind, C.S.; Khera, T.S.; Bueno, C.S. Effects of Residue Decomposition on Productivity and Soil Fertility in Rice–Wheat Rotation. Soil Sci. Soc. Am. J. 2004, 68, 954–964. [Google Scholar] [CrossRef]
- Manevski, K.; Børgesen, C.D.; Li, X.; Andersen, M.N.; Zhang, X.; Abrahamsen, P.; Hu, C.; Hansen, S. Optimising crop production and nitrate leaching in China: Measured and simulated effects of straw incorporation and nitrogen fertilisation. Eur. J. Agron. 2016, 80, 32–44. [Google Scholar] [CrossRef]
- Bahrani, M.; Raufat, M.; Ghadiri, H. Influence of wheat residue management on irrigated corn grain production in a reduced tillage system. Soil Tillage Res. 2007, 94, 305–309. [Google Scholar] [CrossRef]
- IUSS Working Group. World Reference Base for Soil Resources 2014 International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; FAO: Rome, Italy, 2014. [Google Scholar]
- Chen, C.; Travis, A.J.; Hossain, M.; Islam, R.; Price, A.H.; Norton, G.J. Genome-wide association mapping of sodium and potassium concentration in rice grains and shoots under alternate wetting and drying and continuously flooded irrigation. Theor. Appl. Genet. 2021, 134, 2315–2334. [Google Scholar] [CrossRef] [PubMed]
- Ahanger, M.A.; Agarwal, R.M. Salinity stress induced alterations in antioxidant metabolism and nitrogen assimilation in wheat (Triticum aestivum L.) as influenced by potassium supplementation. Plant Physiol. Biochem. 2017, 115, 449–460. [Google Scholar] [CrossRef]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 1998, 135, 1–9. [Google Scholar] [CrossRef]
- Machado, S.; Paulsen, G.M. Combined effects of drought and high temperature on water relations of wheat and sorghum. Plant Soil 2001, 233, 179–187. [Google Scholar] [CrossRef]
- Demidchik, V.; Davenport, R.J.; Tester, M. Nonselective cation channels in plants. Annu. Rev. Plant Biol. 2002, 53, 67–107. [Google Scholar] [CrossRef]
- Thomas, S.C.; Frye, S.; Gale, N.; Garmon, M.; Launchbury, R.; Machado, N.; Melamed, S.; Murray, J.; Petroff, A.; Winsborough, C. Biochar mitigates negative effects of salt additions on two herbaceous plant species. J. Environ. Manag. 2013, 129, 62–68. [Google Scholar] [CrossRef]
- Nunes, M.R.; Vaz, C.M.P.; Denardin, J.E.; van Es, H.M.; Libardi, P.L.; da Silva, A.P. Physicochemical and Structural Properties of an Oxisol under the Addition of Straw and Lime. Soil Sci. Soc. Am. J. 2017, 81, 1328–1339. [Google Scholar] [CrossRef]
- Khamzina, A.; Lamers, J.P.A.; Vlek, P.L.G. Tree establishment under deficit irrigation on degraded agricultural land in the lower Amu Darya River region, Aral Sea Basin. For. Ecol. Manag. 2008, 255, 168–178. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Blumwald, E. Sodium transport and salt tolerance in plants. Curr. Opin. Cell Biol. 2000, 12, 431–434. [Google Scholar] [CrossRef]
- Guo, R.; Shi, L.; Yang, Y. Germination, growth, osmotic adjustment and ionic balance of wheat in response to saline and alkaline stresses. Soil Sci. Plant Nutr. 2009, 55, 667–679. [Google Scholar] [CrossRef]
- Shao, G.C.; Lan, J.J.; Yu, S.E.; Liu, N.; Guo, R.Q.; She, D.L. Photosynthesis and growth of winter wheat in response to waterlogging at different growth stages. Photosynthetica 2013, 51, 429–437. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Torabian, S. Antioxidant enzyme and osmotic adjustment changes in bean seedlings as affected by biochar under salt stress. Ecotoxicol. Environ. Saf. 2017, 137, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Gross, F.; Durner, J.; Gaupels, F. Nitric oxide, antioxidants and prooxidants in plant defence responses. Front. Plant Sci. 2013, 4, 419. [Google Scholar] [CrossRef]
- Li, K.; Xing, C.; Yao, Z.; Huang, X. PbrMYB21, a novel MYB protein of Pyrus betulaefolia, functions in drought tolerance and modulates polyamine levels by regulating arginine decarboxylase gene. Plant Biotechnol. J. 2017, 15, 1186–1203. [Google Scholar] [CrossRef] [PubMed]
- Katerji, N.; van Hoorn, J.W.; Hamdy, A.; Mastrorilli, M. Comparison of corn yield response to plant water stress caused by salinity and by drought. Agric. Water Manag. 2004, 65, 95–101. [Google Scholar] [CrossRef]
- Vyrides, I.; Stuckey, D.C. Compatible solute addition to biological systems treating waste/wastewater to counteract osmotic and other environmental stresses: A review. Crit. Rev. Biotechnol. 2017, 37, 865–879. [Google Scholar] [CrossRef] [PubMed]
- Khavari-Nejad, R.A.; Mostofi, Y. Effects of NaCl on photosynthetic pigments, saccharides, and chloroplast ultrastructure in leaves of tomato cultivars. Photosynthetica 1997, 35, 151–154. [Google Scholar] [CrossRef]
- Liu, N.; Jin, Z.; Wang, S.; Gong, B.; Wen, D.; Wang, X.; Wei, M.; Shi, Q. Sodic alkaline stress mitigation with exogenous melatonin involves reactive oxygen metabolism and ion homeostasis in tomato. Sci. Hortic. 2015, 181, 18–25. [Google Scholar] [CrossRef]
- Uddling, J.; Gelang-Alfredsson, J.; Piikki, K.; Pleijel, H. Evaluating the relationship between leaf chlorophyll concentration and SPAD-502 chlorophyll meter readings. Photosynth. Res. 2007, 91, 37–46. [Google Scholar] [CrossRef]
- van Hoorn, J.; Katerji, N.; Hamdy, A.; Mastrorilli, M. Effect of salinity on yield and nitrogen uptake of four grain legumes and on biological nitrogen contribution from the soil. Agric. Water Manag. 2001, 51, 87–98. [Google Scholar] [CrossRef]
- Nieva, F.J.J.; Castellanos, E.M.; Figueroa, M.E.; Gil, F. Gas Exchange and Chlorophyll Fluorescence of C3 and C4 Saltmarsh Species. Photosynthetica 1999, 36, 397–406. [Google Scholar] [CrossRef]
- Wang, H.; Yang, G.; Wang, X.; Zhao, C.; Muhammad, N.; Chen, Y.; Hu, Y. Oilseed rape straw returning changes soil reducibility and affects the root and yield of rice in the rape-rice rotation field at Sichuan Basin area of China. Agron. J. 2020, 112, 4681–4692. [Google Scholar] [CrossRef]
- van de Weg, M.J.; Shaver, G.R.; Salmon, V.G. Contrasting effects of long term versus short-term nitrogen addition on photosynthesis and respiration in the Arctic. Plant Ecol. 2013, 214, 1273–1286. [Google Scholar] [CrossRef]
- Gaude, N.; Bréhélin, C.; Tischendorf, G.; Kessler, F.; Dörmann, P. Nitrogen deficiency in Arabidopsis affects galactolipid composition and gene expression and results in accumulation of fatty acid phytyl esters. Plant J. 2007, 49, 729–739. [Google Scholar] [CrossRef]
- Iqbal, M.; Ashraf, M. Changes in growth, photosynthetic capacity and ionic relations in spring wheat (Triticum aestivum L.) due to pre-sowing seed treatment with polyamines. Plant Growth Regul. 2005, 46, 19–30. [Google Scholar] [CrossRef]
- Yang, J.Y.; Zheng, W.; Tian, Y.; Wu, Y.; Zhou, D.W. Effects of various mixed salt-alkaline stresses on growth, photosynthesis, and photosynthetic pigment concentrations of Medicago ruthenica seedlings. Photosynthetica 2011, 49, 275–284. [Google Scholar] [CrossRef]
- Shah, A.N.; Yang, G.; Tanveer, M.; Iqbal, J. Leaf gas exchange, source–sink relationship, and growth response of cotton to the interactive effects of nitrogen rate and planting density. Acta Physiol. Plant. 2017, 39, 119. [Google Scholar] [CrossRef]
- Guan, X.-K.; Wei, L.; Turner, N.C.; Ma, S.-C.; Yang, M.-D.; Wang, T.-C. Improved straw management practices promote in situ straw decomposition and nutrient release, and increase crop production. J. Clean. Prod. 2020, 250, 119514. [Google Scholar] [CrossRef]
- Ran, C.; Gao, D.; Bai, T.; Geng, Y.; Shao, X.; Guo, L. Straw return alleviates the negative effects of saline sodic stress on rice by improving soil chemistry and reducing the accumulation of sodium ions in rice leaves. Agric. Ecosyst. Environ. 2022, 342, 108253. [Google Scholar] [CrossRef]
- Chen, B.; Liu, E.; Tian, Q.; Yan, C.; Zhang, Y. Soil nitrogen dynamics and crop residues. A review. Agron. Sustain. Dev. 2014, 34, 429–442. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Unit | Mean | Parameter | Unit | Mean |
|---|---|---|---|---|---|
| Bulk density | g cm−3 | 1.61 | Total N | g kg−1 | 0.27 |
| pH | - | 10.10 | Total K | g kg−1 | 22.36 |
| Salinity | g kg−1 | 4.78 | Organic matter | g kg−1 | 7.31 |
| ENa+ | cmolc kg−1 | 3.21 | Alkalihydrolysis N | mg kg−1 | 16.3 |
| CEC | cmolc kg−1 | 12.97 | Available P | mg kg−1 | 16.90 |
| ESP | % | 24.74 | Available K | mg kg−1 | 107.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dang, K.; Ran, C.; Tian, H.; Gao, D.; Mu, J.; Zhang, Z.; Geng, Y.; Zhang, Q.; Shao, X.; Guo, L. Combined Effects of Straw Return with Nitrogen Fertilizer on Leaf Ion Balance, Photosynthetic Capacity, and Rice Yield in Saline-Sodic Paddy Fields. Agronomy 2023, 13, 2274. https://doi.org/10.3390/agronomy13092274
Dang K, Ran C, Tian H, Gao D, Mu J, Zhang Z, Geng Y, Zhang Q, Shao X, Guo L. Combined Effects of Straw Return with Nitrogen Fertilizer on Leaf Ion Balance, Photosynthetic Capacity, and Rice Yield in Saline-Sodic Paddy Fields. Agronomy. 2023; 13(9):2274. https://doi.org/10.3390/agronomy13092274
Chicago/Turabian StyleDang, Kun, Cheng Ran, Hao Tian, Dapeng Gao, Jinmeng Mu, Zhenyu Zhang, Yanqiu Geng, Qiang Zhang, Xiwen Shao, and Liying Guo. 2023. "Combined Effects of Straw Return with Nitrogen Fertilizer on Leaf Ion Balance, Photosynthetic Capacity, and Rice Yield in Saline-Sodic Paddy Fields" Agronomy 13, no. 9: 2274. https://doi.org/10.3390/agronomy13092274