
Identification, Characterization, and Expression Profiling of Maize GATA Gene Family in Response to Abiotic and Biotic Stresses


Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification and Chromosome Mapping of GATA Family Genes in Maize
2.2. Phylogenetic Analysis of GATA Family Genes in Maize
2.3. Collinearity Analysis of GATA Family Genes in Maize
2.4. Reanalysis of Maize Transcriptome Sequencing Big Data through RNA-Seq
2.5. Tissue-Specific Expression Analysis of GATA Family Genes in Maize
2.6. Expression Pattern Analysis of GATA Family Genes in Maize under Abiotic Stress and Biotic Stress
2.7. Protein Interaction Network Prediction
3. Results
3.1. Basic Information of GATA Gene Family Members in Maize
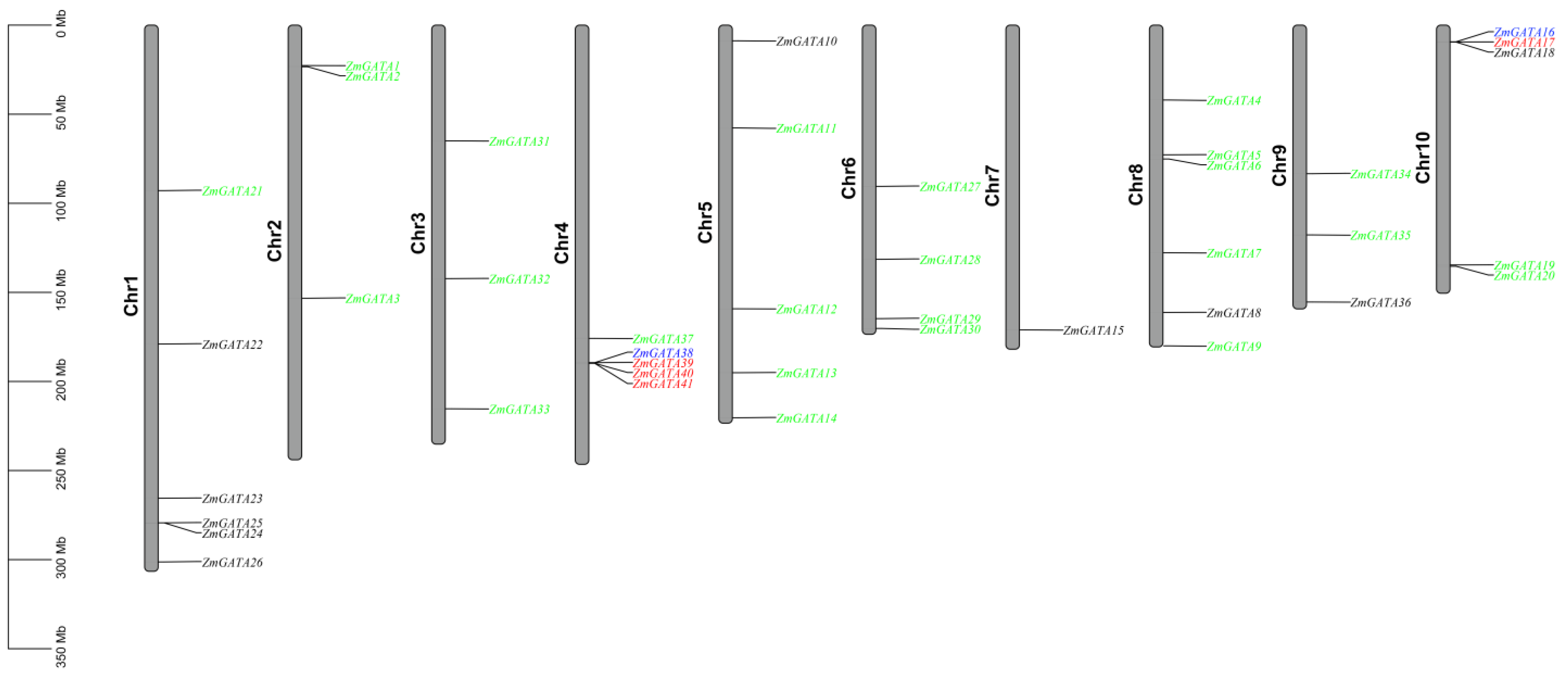
3.2. Chromosome Mapping of GATA Family Genes in Maize
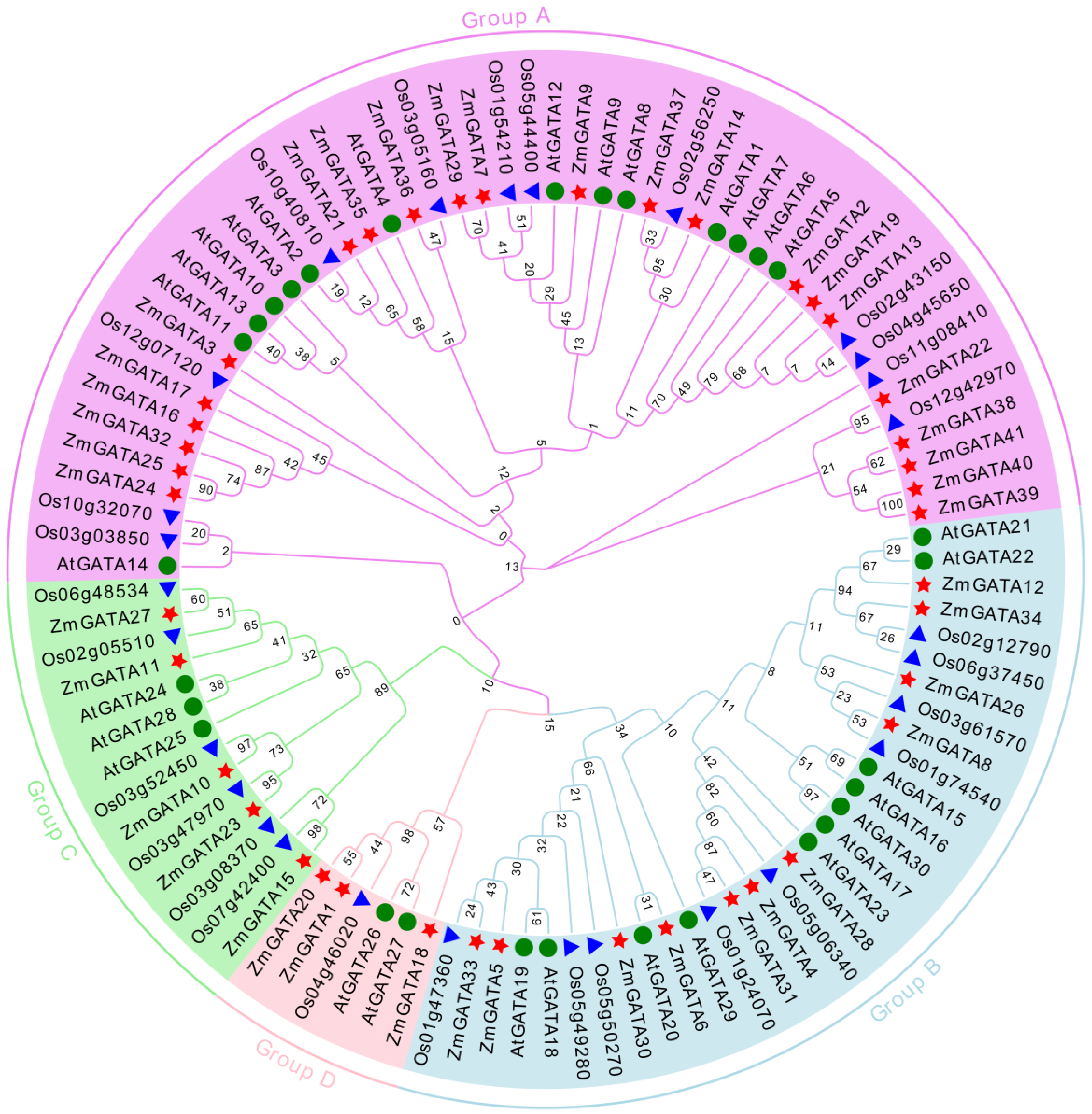
3.3. Cluster Analysis of GATA Family Genes in Maize, A. thaliana, and Rice
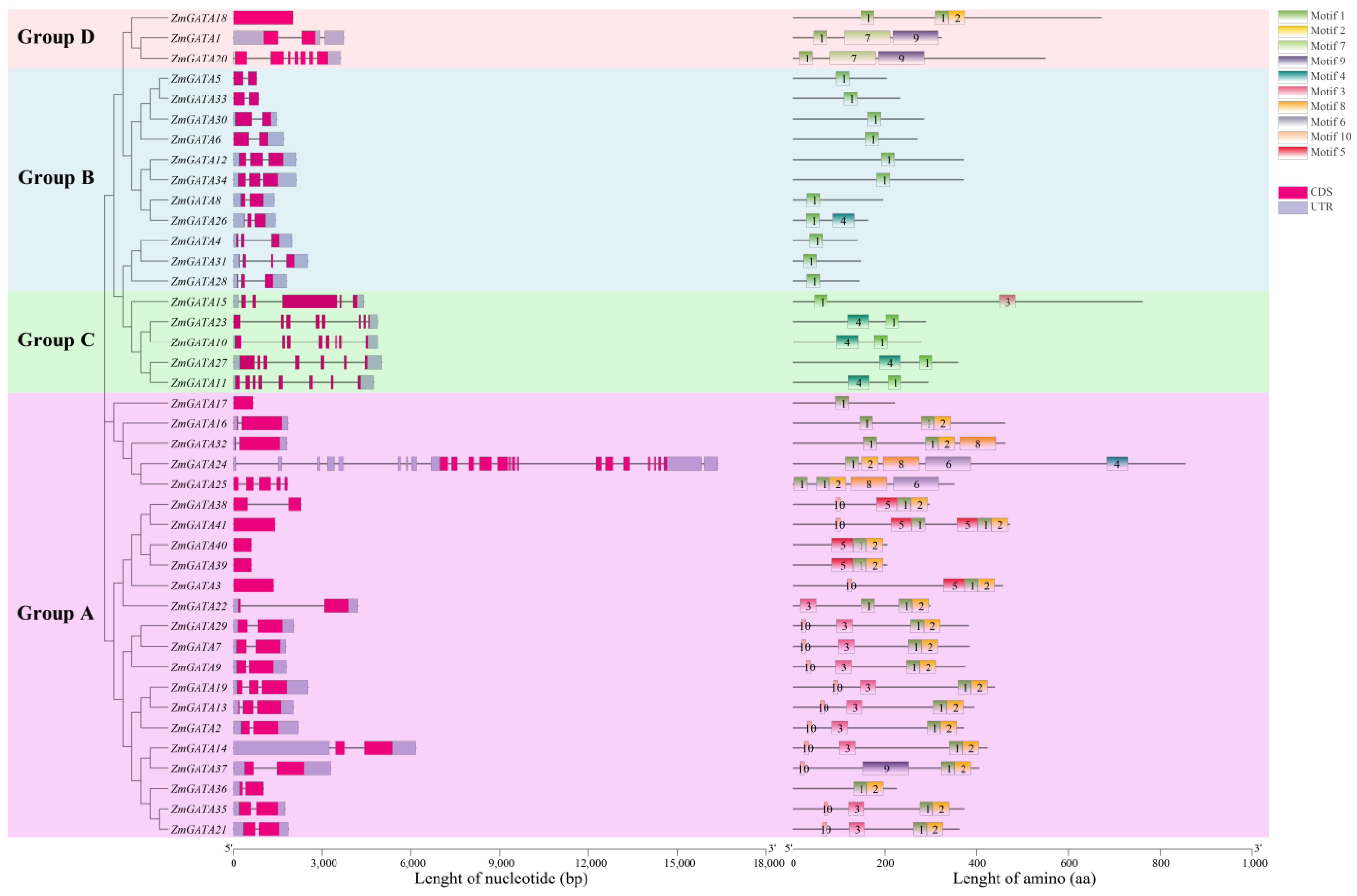
3.4. Gene Structure and Conservated Sequence Analysis of GATA Family in Maize
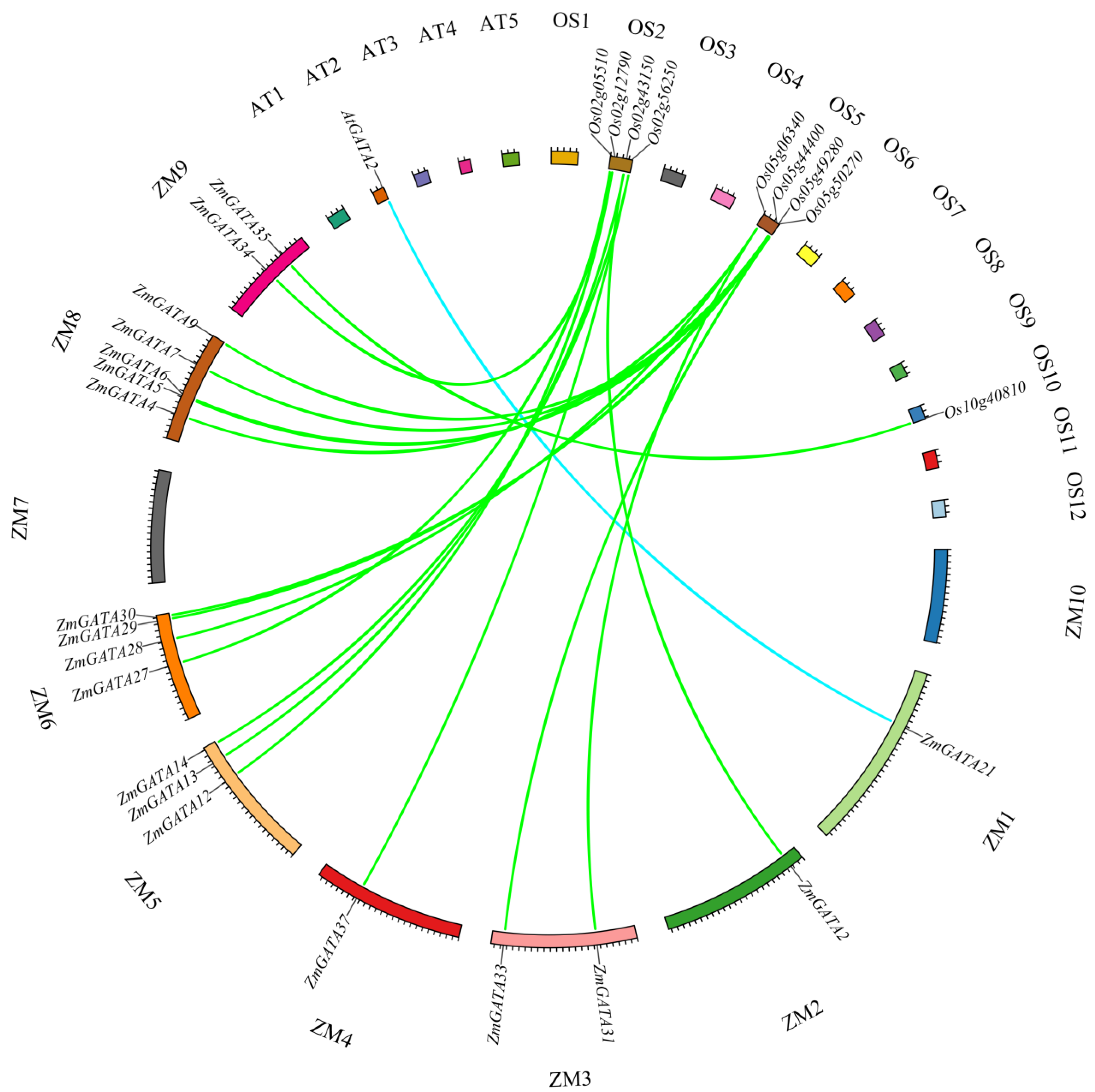
3.5. Collinearity Analysis of GATA Family Genes in Maize, A. thaliana, and Rice
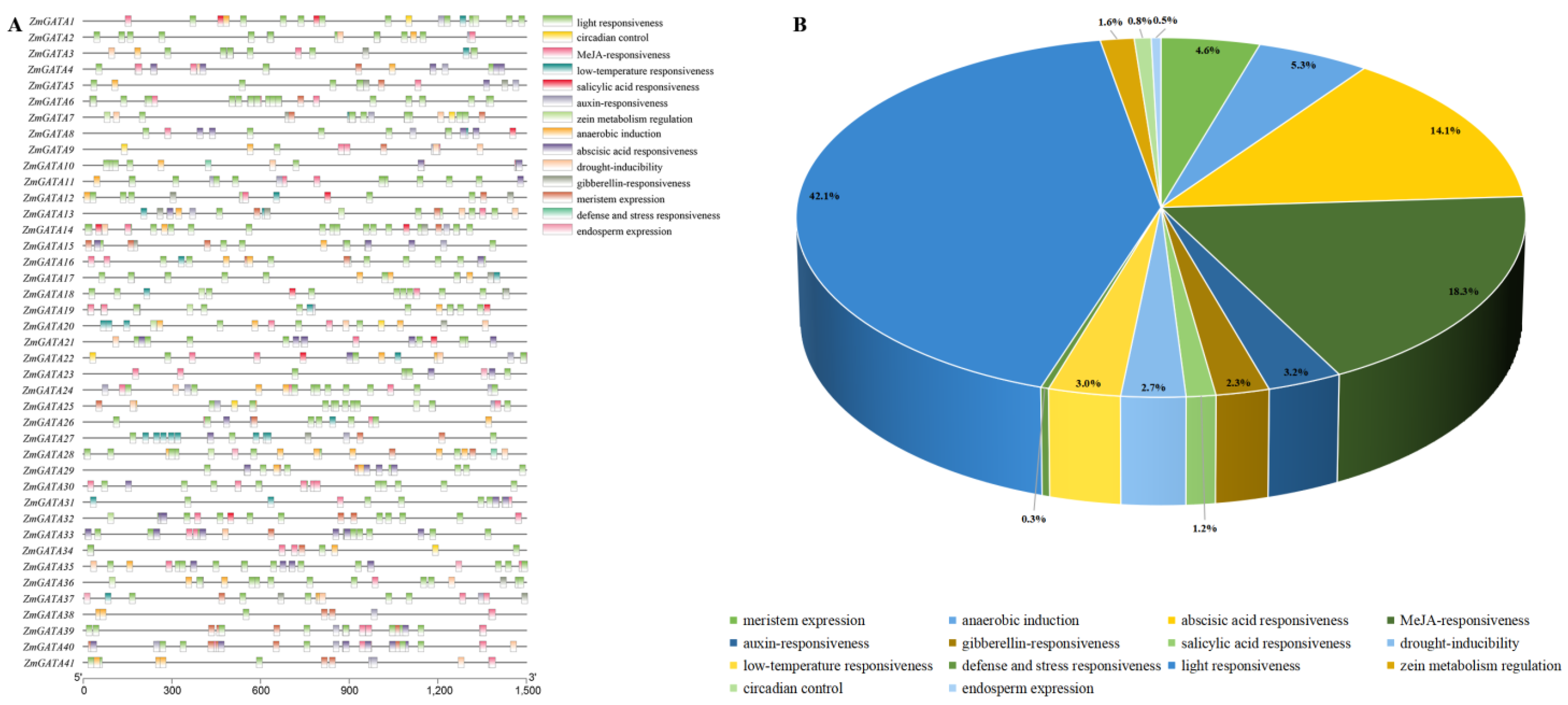
3.6. Analysis of Cis-Acting Elements of Promoter Sequences of GATA Family Genes in Maize
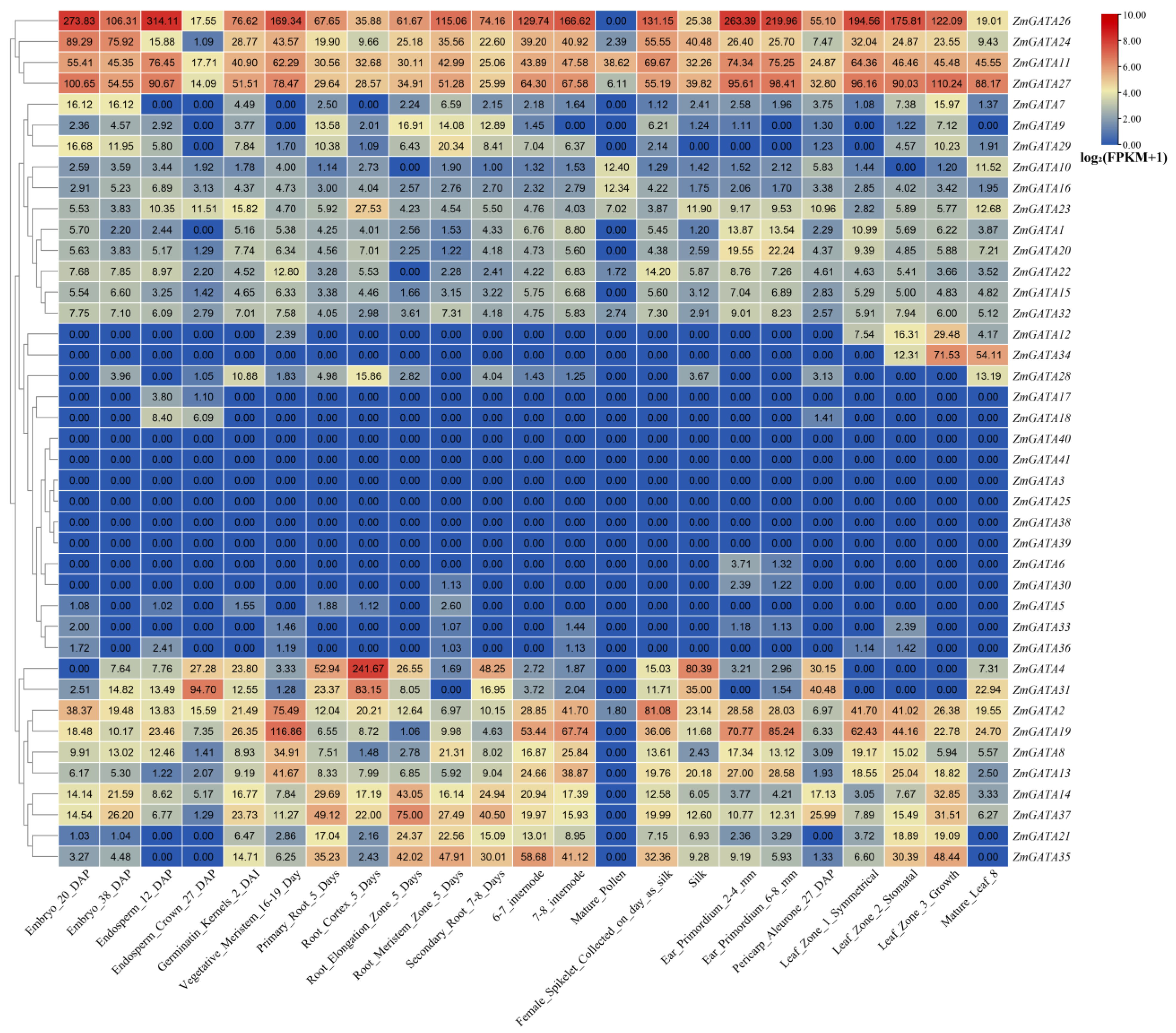
3.7. Tissue-Specific Expression Analysis of GATA Family Genes in Maize
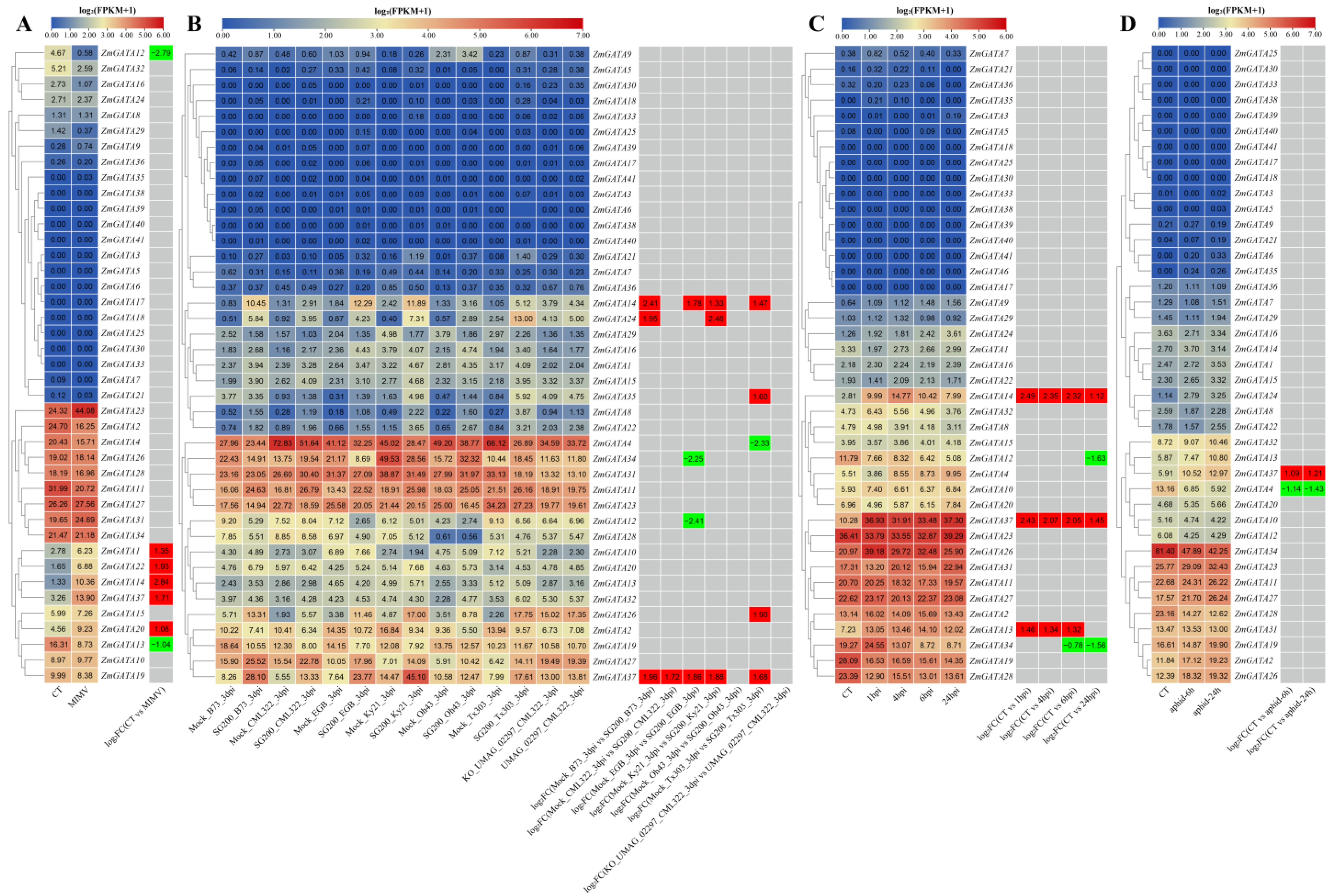
3.8. Expression Pattern Analysis of Maize GATA Family Genes under Abiotic Stress
3.9. Expression Pattern Analysis of Maize GATA Family Genes under Biotic Stress
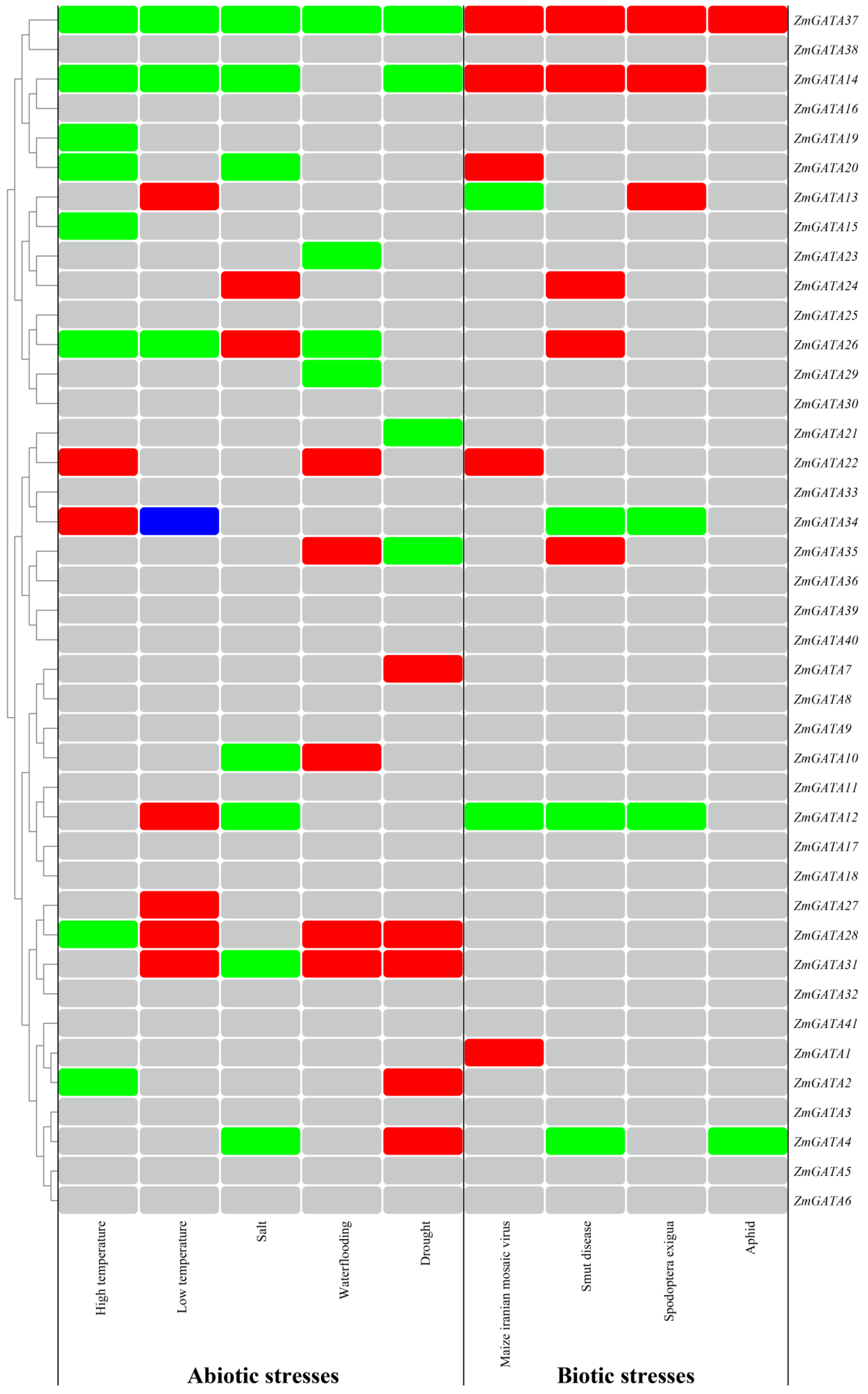
3.10. Analysis of Regulation Mode of Maize GATA Family Genes under Abiotic and Biotic Stresses
3.11. Protein–Protein Interaction Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Benz, B.F. Archaeological evidence of teosinte domestication from Guilá Naquitz, Oaxaca. Proc. Natl. Acad. Sci. USA 2001, 98, 2104–2106. [Google Scholar] [CrossRef] [PubMed]
- Dolgin, E. Maize genome mapped. Nat. News 2009, 1098. [Google Scholar] [CrossRef]
- Ramazan, S.; Nazir, I.; Yousuf, W.; John, R.; Allakhverdiev, S. Environmental stress tolerance in maize (Zea mays): Role of polyamine metabolism. Funct. Plant Biol. 2022, 50, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Guan, Y.; Chen, S.; Li, H. Genome-wide analysis of basic helix-loop-helix (bHLH) transcription factors in Brachypodium distachyon. BMC Genom. 2017, 18, 619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Ma, S.; Ye, N.; Jiang, M.; Cao, J.; Zhang, J. WRKY transcription factors in plant responses to stresses. J. Integr. Plant Biol. 2017, 59, 86–101. [Google Scholar] [CrossRef] [Green Version]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY transcription factors. Trends Plant Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef]
- Dröge-Laser, W.; Snoek, B.L.; Snel, B.; Weiste, C. The Arabidopsis bZIP transcription factor family—An update. Curr. Opin. Plant Biol. 2018, 45, 36–49. [Google Scholar] [CrossRef]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, J.; Hu, Z.; Guo, X.; Tian, S.; Chen, G. Genome-wide analysis of the MADS-box transcription factor family in Solanum lycopersicum. Int. J. Mol. Sci. 2019, 20, 2961. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Xi, H.; Park, S.; Yun, Y.; Park, J. Genome-wide comparative analyses of GATA transcription factors among seven Populus genomes. Sci. Rep. 2021, 11, 16578. [Google Scholar] [CrossRef]
- Reyes, J.C.; Muro-Pastor, M.I.; Florencio, F.J. The GATA family of transcription factors in Arabidopsis and rice. Plant Physiol. 2004, 134, 1718–1732. [Google Scholar] [CrossRef] [Green Version]
- Hudson, D.; Guevara, D.; Yaish, M.W.; Hannam, C.; Long, N.; Clarke, J.D.; Bi, Y.-M.; Rothstein, S.J. GNC and CGA1 modulate chlorophyll biosynthesis and glutamate synthase (GLU1/Fd-GOGAT) expression in Arabidopsis. PLoS ONE 2011, 6, e26765. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.-Y.; Chao, D.-Y.; Gao, J.-P.; Zhu, M.-Z.; Shi, M.; Lin, H.-X. A previously unknown zinc finger protein, DST, regulates drought and salt tolerance in rice via stomatal aperture control. Genes Dev. 2009, 23, 1805–1817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scazzocchio, C. The fungal GATA factors. Curr. Opin. Microbiol. 2000, 3, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Lowry, J.A.; Atchley, W.R. Molecular evolution of the GATA family of transcription factors: Conservation within the DNA-binding domain. J. Mol. Evol. 2000, 50, 103–115. [Google Scholar] [CrossRef]
- Yuan, Q.; Zhang, C.; Zhao, T.; Yao, M.; Xu, X. A genome-wide analysis of GATA transcription factor family in tomato and analysis of expression patterns. Int. J. Agric. Biol. 2018, 20, 1274–1282. [Google Scholar]
- Wang, T.; Yang, Y.; Lou, S.; Wei, W.; Zhao, Z.; Ren, Y.; Lin, C.; Ma, L. Genome-wide characterization and gene expression analyses of GATA transcription factors in Moso bamboo (Phyllostachys edulis). Int. J. Mol. Sci. 2019, 21, 14. [Google Scholar] [CrossRef] [Green Version]
- Evans, T.; Reitman, M.; Felsenfeld, G. An erythrocyte-specific DNA-binding factor recognizes a regulatory sequence common to all chicken globin genes. Proc. Natl. Acad. Sci. USA 1988, 85, 5976–5980. [Google Scholar] [CrossRef]
- Daniel-Vedele, F.; Caboche, M. A tobacco cDNA clone encoding a GATA-1 zinc finger protein homologous to regulators of nitrogen metabolism in fungi. Mol. Gen. Genet. MGG 1993, 240, 365–373. [Google Scholar] [CrossRef]
- Manfield, I.W.; Devlin, P.F.; Jen, C.-H.; Westhead, D.R.; Gilmartin, P.M. Conservation, convergence, and divergence of light-responsive, circadian-regulated, and tissue-specific expression patterns during evolution of the Arabidopsis GATA gene family. Plant Physiol. 2007, 143, 941–958. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Nutan, K.K.; Singla-Pareek, S.L.; Pareek, A. Abiotic stresses cause differential regulation of alternative splice forms of GATA transcription factor in rice. Front. Plant Sci. 2017, 8, 1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Hou, Y.; Hao, Q.; Chen, H.; Chen, L.; Yuan, S.; Shan, Z.; Zhang, X.; Yang, Z.; Qiu, D. Genome-wide survey of the soybean GATA transcription factor gene family and expression analysis under low nitrogen stress. PLoS ONE 2015, 10, e0125174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Zou, X.; Huang, Z.; Fan, S.; Qun, G.; Liu, A.; Gong, J.; Li, J.; Gong, W.; Shi, Y. Genome-wide identification and analysis of the evolution and expression patterns of the GATA transcription factors in three species of Gossypium genus. Gene 2019, 680, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Guo, Y.; Chen, Y.; Wu, D.; Jiang, L. Genome-wide identification, phylogenetic and expression pattern analysis of GATA family genes in Brassica napus. BMC Plant Biol. 2020, 20, 543. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Shao, H.; Li, K.; Zhang, D.; Fan, S.; Li, Y.; Han, M. Genome-wide identification, evolution, and expression analysis of GATA transcription factors in apple (Malus × domestica Borkh.). Gene 2017, 627, 460–472. [Google Scholar] [CrossRef]
- Peng, X.; Wu, Q.; Teng, L.; Tang, F.; Pi, Z.; Shen, S. Transcriptional regulation of the paper mulberry under cold stress as revealed by a comprehensive analysis of transcription factors. BMC Plant Biol. 2015, 15, 108. [Google Scholar] [CrossRef] [Green Version]
- Peng, W.; Li, W.; Song, N.; Tang, Z.; Liu, J.; Wang, Y.; Pan, S.; Dai, L.; Wang, B. Genome-wide characterization, evolution, and expression profile analysis of GATA transcription factors in Brachypodium distachyon. Int. J. Mol. Sci. 2021, 22, 2026. [Google Scholar] [CrossRef]
- Zhang, Z.; Ren, C.; Zou, L.; Wang, Y.; Li, S.; Liang, Z. Characterization of the GATA gene family in Vitis vinifera: Genome-wide analysis, expression profiles, and involvement in light and phytohormone response. Genome 2018, 61, 713–723. [Google Scholar] [CrossRef] [Green Version]
- Ravindran, P.; Verma, V.; Stamm, P.; Kumar, P.P. A novel RGL2–DOF6 complex contributes to primary seed dormancy in Arabidopsis thaliana by regulating a GATA transcription factor. Mol. Plant 2017, 10, 1307–1320. [Google Scholar] [CrossRef] [Green Version]
- Lu, G.; Casaretto, J.A.; Ying, S.; Mahmood, K.; Liu, F.; Bi, Y.-M.; Rothstein, S.J. Overexpression of OsGATA12 regulates chlorophyll content, delays plant senescence and improves rice yield under high density planting. Plant Mol. Biol. 2017, 94, 215–227. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Zhang, Y.; Zhang, L.L.; Huang, H.Y.; Yang, B.J.; Luan, S.; Xue, H.W.; Lin, W.H. OsGATA7 modulates brassinosteroids-mediated growth regulation and influences architecture and grain shape. Plant Biotechnol. J. 2018, 16, 1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, Y.; Zhou, Y.; Han, X.; Shen, C.; Wang, S.; Liu, C.; Yin, W.; Xia, X. The GATA transcription factor GNC plays an important role in photosynthesis and growth in poplar. J. Exp. Bot. 2020, 71, 1969–1984. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Ingvardsen, C.R.; Lübberstedt, T.; Xu, M. The Pic19 NBS-LRR gene family members are closely linked to Scmv1, but not involved in maize resistance to sugarcane mosaic virus. Genome 2008, 51, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Zhao, Y.; Li, X.; Wu, M.; Chai, W.; Sheng, L.; Wang, Y.; Dong, Q.; Jiang, H.; Cheng, B. Genomewide identification, classification and analysis of NAC type gene family in maize. J. Genet. 2015, 94, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Pan, J.; Zhang, D.; Jiang, S.; Cai, G.; Wang, L.; Li, D. Identification of mitogen-activated protein kinase kinase gene family and MKK–MAPK interaction network in maize. Biochem. Biophys. Res. Commun. 2013, 441, 964–969. [Google Scholar] [CrossRef]
- Jiang, L.; Hu, W.; Qian, Y.; Ren, Q.; Zhang, J. Genome-wide identification, classification and expression analysis of the Hsf and Hsp70 gene families in maize. Gene 2021, 770, 145348. [Google Scholar] [CrossRef]
- Jiang, L.; Yu, X.; Chen, D.; Feng, H.; Li, J. Identification, phylogenetic evolution and expression analysis of GATA transcription factor family in maize (Zea mays). Int. J. Agric. Biol. 2020, 23, 637–643. [Google Scholar]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.; Tosatto, S.C.; Paladin, L.; Raj, S.; Richardson, L.J. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39 (Suppl. S2), W29–W37. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent updates, new developments and status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. [Google Scholar] [CrossRef]
- Xiong, E.; Zheng, C.; Wu, X.; Wang, W. Protein subcellular location: The gap between prediction and experimentation. Plant Mol. Biol. Report. 2016, 34, 52–61. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Zhang, H.; Gao, S.; Lercher, M.J.; Chen, W.-H.; Hu, S. Evolview v2: An online visualization and management tool for customized and annotated phylogenetic trees. Nucleic Acids Res. 2016, 44, W236–W241. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.; Pirrung, M.; McCue, L.A. FQC Dashboard: Integrates FastQC results into a web-based, interactive, and extensible FASTQ quality control tool. Bioinformatics 2017, 33, 3137–3139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 genome project data processing subgroup. The sequence alignment/map (SAM) format and SAMtools. Bioinformatics 2009, 25, 2079. [Google Scholar]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Varet, H.; Brillet-Guéguen, L.; Coppée, J.-Y.; Dillies, M.-A. SARTools: A DESeq2-and EdgeR-based R pipeline for comprehensive differential analysis of RNA-Seq data. PLoS ONE 2016, 11, e0157022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coles, N.D.; McMullen, M.D.; Balint-Kurti, P.J.; Pratt, R.C.; Holland, J.B. Genetic control of photoperiod sensitivity in maize revealed by joint multiple population analysis. Genetics 2010, 184, 799–812. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, X.; Li, Y.; Zhang, Y.; Gou, Z.; Qi, X.; Zhang, J. Transcriptomic analysis revealed the common and divergent responses of maize seedling leaves to cold and heat stresses. Genes 2020, 11, 881. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Liu, C.; Zhang, D.; He, C.; Zhang, J.; Li, Z. Effects of maize organ-specific drought stress response on yields from transcriptome analysis. BMC Plant Biol. 2019, 19, 335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, F.; Tan, Z.; Fang, T.; Tang, K.; Liang, K.; Qiu, F. A comprehensive transcriptomics analysis reveals long non-coding RNA to be involved in the key metabolic pathway in response to waterlogging stress in maize. Genes 2020, 11, 267. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Wang, Y.; Zhang, Y.; Li, C.; Gong, S.; Yan, S.; Li, G.; Hu, G.; Ren, H.; Yang, J. Comparative transcriptome analysis of salt-sensitive and salt-tolerant maize reveals potential mechanisms to enhance salt resistance. Genes Genom. 2019, 41, 781–801. [Google Scholar] [CrossRef]
- Schurack, S.; Depotter, J.R.; Gupta, D.; Thines, M.; Doehlemann, G. Comparative transcriptome profiling identifies maize line specificity of fungal effectors in the maize—Ustilago maydis interaction. Plant J. 2021, 106, 733–752. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, A.; Izadpanah, K.; Dietzgen, R.G. Changes in maize transcriptome in response to maize Iranian mosaic virus infection. PLoS ONE 2018, 13, e0194592. [Google Scholar] [CrossRef] [Green Version]
- Tzin, V.; Hojo, Y.; Strickler, S.R.; Bartsch, L.J.; Archer, C.M.; Ahern, K.R.; Zhou, S.; Christensen, S.A.; Galis, I.; Mueller, L.A. Rapid defense responses in maize leaves induced by Spodoptera exigua caterpillar feeding. J. Exp. Bot. 2017, 68, 4709–4723. [Google Scholar] [CrossRef] [Green Version]
- Database Resources of the National Genomics Data Center, China National Center for Bioinformation in 2023. Nucleic Acids Res. 2023, 51, D18–D28. [CrossRef] [PubMed]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.T.; Pyysalo, S. The STRING database in 2023: Protein–protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 2023, 51, D638–D646. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.P.; Robin Buell, C. Advances in plant genome sequencing. Plant J. 2012, 70, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Monaco, M.K.; Stein, J.; Naithani, S.; Wei, S.; Dharmawardhana, P.; Kumari, S.; Amarasinghe, V.; Youens-Clark, K.; Thomason, J.; Preece, J. Gramene 2013: Comparative plant genomics resources. Nucleic Acids Res. 2014, 42, D1193–D1199. [Google Scholar] [CrossRef] [Green Version]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Wu, N.; Song, W.; Yin, G.; Qin, Y.; Yan, Y.; Hu, Y. Soybean (Glycine max) expansin gene superfamily origins: Segmental and tandem duplication events followed by divergent selection among subfamilies. BMC Plant Biol. 2014, 14, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, K.; Wu, W.-w.; Diao, Q.-y. Application and research progress on transcriptomics. Biotechnol. Bull. 2019, 35, 1. [Google Scholar]
- Zhang, H.; Wu, T.; Li, Z.; Huang, K.; Kim, N.-E.; Ma, Z.; Kwon, S.-W.; Jiang, W.; Du, X. OsGATA16, a GATA transcription factor, confers cold tolerance by repressing OsWRKY45–1 at the seedling stage in rice. Rice 2021, 14, 42. [Google Scholar] [CrossRef]
- Nutan, K.K.; Rathore, R.S.; Tripathi, A.K.; Mishra, M.; Pareek, A.; Singla-Pareek, S.L. Integrating the dynamics of yield traits in rice in response to environmental changes. J. Exp. Bot. 2020, 71, 490–506. [Google Scholar] [CrossRef]
- Zhao, T.; Wu, T.; Pei, T.; Wang, Z.; Yang, H.; Jiang, J.; Zhang, H.; Chen, X.; Li, J.; Xu, X. Overexpression of SlGATA17 promotes drought tolerance in transgenic tomato plants by enhancing activation of the phenylpropanoid biosynthetic pathway. Front. Plant Sci. 2021, 12, 634888. [Google Scholar] [CrossRef]
- He, H.; Zhu, S.; Jiang, Z.; Ji, Y.; Wang, F.; Zhao, R.; Bie, T. Comparative mapping of powdery mildew resistance gene Pm21 and functional characterization of resistance-related genes in wheat. Theor. Appl. Genet. 2016, 129, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Jia, L.; Yang, D.; Hu, Y.; Njogu, M.K.; Wang, P.; Lu, X.; Yan, C. Genome-wide identification, phylogenetic and expression pattern analysis of gata family genes in cucumber (Cucumis sativus L.). Plants 2021, 10, 1626. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Locus Name | CDS Size (bp) | Number of Amino Acids (aa) | Molecular Weight (kD) | pI | Instability Index | Aliphatic Index | Grand Average of Hydropathicity | Prediction of Subcellular Location |
|---|---|---|---|---|---|---|---|---|---|
| ZmGATA1 | Zm00001d002790 | 972 | 323 | 35.83 | 8.55 | 56.48 | 61.33 | −0.591 | Extracell |
| ZmGATA2 | Zm00001d002811 | 1116 | 371 | 39.52 | 7.11 | 68.05 | 57.44 | −0.682 | Nucleus |
| ZmGATA3 | Zm00001d005005 | 1371 | 456 | 46.84 | 6.37 | 64.87 | 55.18 | −0.468 | Chloroplast |
| ZmGATA4 | Zm00001d009193 | 420 | 139 | 14.87 | 9.82 | 65.52 | 65.40 | −0.612 | Chloroplast |
| ZmGATA5 | Zm00001d009604 | 612 | 203 | 21.78 | 8.96 | 70.73 | 54.48 | −0.504 | Chloroplast |
| ZmGATA6 | Zm00001d009668 | 813 | 270 | 28.14 | 8.90 | 66.06 | 52.33 | −0.339 | Chloroplast |
| ZmGATA7 | Zm00001d010785 | 1155 | 384 | 40.37 | 5.63 | 56.44 | 71.77 | −0.462 | Nucleus |
| ZmGATA8 | Zm00001d011771 | 588 | 195 | 21.24 | 9.71 | 61.25 | 57.28 | −0.839 | Nucleus |
| ZmGATA9 | Zm00001d012757 | 1128 | 375 | 39.41 | 5.57 | 61.17 | 66.45 | −0.342 | Nucleus |
| ZmGATA10 | Zm00001d013331 | 837 | 278 | 29.77 | 8.60 | 52.50 | 67.88 | −0.453 | Nucleus |
| ZmGATA11 | Zm00001d014656 | 882 | 293 | 30.87 | 9.23 | 45.07 | 64.44 | −0.462 | Nucleus |
| ZmGATA12 | Zm00001d016361 | 1113 | 370 | 39.51 | 9.35 | 64.31 | 63.16 | −0.431 | Nucleus |
| ZmGATA13 | Zm00001d017409 | 1185 | 394 | 41.93 | 8.43 | 55.95 | 73.86 | −0.392 | Nucleus |
| ZmGATA14 | Zm00001d018421 | 1269 | 422 | 43.43 | 5.55 | 68.03 | 65.17 | −0.319 | Nucleus |
| ZmGATA15 | Zm00001d022142 | 2283 | 760 | 86.75 | 8.64 | 47.99 | 72.05 | −0.417 | Nucleus |
| ZmGATA16 | Zm00001d023539 | 1386 | 461 | 49.37 | 9.44 | 77.46 | 65.23 | −0.652 | Chloroplast |
| ZmGATA17 | Zm00001d023540 | 666 | 221 | 23.33 | 8.25 | 68.06 | 58.05 | −0.618 | Nucleus |
| ZmGATA18 | Zm00001d023541 | 2016 | 671 | 73.79 | 7.37 | 74.14 | 66.66 | −0.732 | Nucleus |
| ZmGATA19 | Zm00001d025953 | 1317 | 438 | 46.17 | 9.07 | 69.15 | 59.86 | −0.548 | Chloroplast |
| ZmGATA20 | Zm00001d025988 | 1650 | 549 | 60.54 | 6.23 | 58.35 | 70.18 | −0.586 | Nucleus |
| ZmGATA21 | Zm00001d029896 | 1086 | 361 | 37.45 | 7.34 | 58.15 | 47.01 | −0.522 | Nucleus |
| ZmGATA22 | Zm00001d031135 | 900 | 299 | 33.33 | 8.63 | 61.34 | 51.24 | −0.925 | Nucleus |
| ZmGATA23 | Zm00001d033523 | 867 | 288 | 30.65 | 4.61 | 56.22 | 63.85 | −0.732 | Cytoplasm |
| ZmGATA24 | Zm00001d033945 | 2565 | 854 | 94.10 | 8.93 | 49.17 | 75.66 | −0.395 | Nucleus |
| ZmGATA25 | Zm00001d033946 | 1053 | 350 | 38.62 | 9.24 | 47.42 | 81.17 | −0.369 | Chloroplast |
| ZmGATA26 | Zm00001d034751 | 492 | 163 | 17.99 | 9.97 | 77.21 | 58.22 | −0.887 | Nucleus |
| ZmGATA27 | Zm00001d036494 | 1077 | 358 | 38.13 | 5.09 | 45.52 | 68.55 | −0.568 | Nucleus |
| ZmGATA28 | Zm00001d037605 | 432 | 143 | 15.57 | 9.99 | 72.01 | 62.87 | −0.628 | Nucleus |
| ZmGATA29 | Zm00001d038801 | 1149 | 382 | 39.65 | 5.65 | 55.76 | 73.72 | −0.364 | Cytoplasm |
| ZmGATA30 | Zm00001d039113 | 855 | 284 | 29.95 | 8.57 | 64.20 | 54.19 | −0.351 | Chloroplast |
| ZmGATA31 | Zm00001d040775 | 444 | 147 | 16.05 | 9.43 | 66.01 | 65.65 | −0.660 | Nucleus |
| ZmGATA32 | Zm00001d041883 | 1386 | 461 | 49.38 | 10.03 | 67.15 | 62.52 | −0.733 | Chloroplast |
| ZmGATA33 | Zm00001d043969 | 702 | 233 | 23.57 | 7.45 | 50.40 | 57.73 | −0.137 | Mitochondrion |
| ZmGATA34 | Zm00001d046354 | 1113 | 370 | 38.98 | 8.50 | 55.17 | 67.68 | −0.285 | Nucleus |
| ZmGATA35 | Zm00001d047081 | 1122 | 373 | 38.55 | 8.22 | 56.25 | 45.76 | −0.521 | Nucleus |
| ZmGATA36 | Zm00001d048391 | 681 | 226 | 23.84 | 6.40 | 72.93 | 59.91 | −0.514 | Nucleus |
| ZmGATA37 | Zm00001d051981 | 1218 | 405 | 42.28 | 5.33 | 69.74 | 64.02 | −0.424 | Nucleus |
| ZmGATA38 | Zm00001d052412 | 894 | 297 | 31.02 | 6.44 | 73.57 | 64.88 | −0.497 | Nucleus |
| ZmGATA39 | Zm00001d052413 | 615 | 204 | 22.13 | 10.23 | 74.88 | 62.75 | −0.702 | Nucleus |
| ZmGATA40 | Zm00001d052430 | 615 | 204 | 22.21 | 10.23 | 73.62 | 62.25 | −0.732 | Nucleus |
| ZmGATA41 | Zm00001d052431 | 1419 | 472 | 49.21 | 9.26 | 74.40 | 60.11 | −0.579 | Nucleus |
| Motif | Sequence | Number of Amino Acids | Pfam Annotation |
|---|---|---|---|
| motif 1 | HCGTTKTPQWRSGPLGPKTLCNACGVRYK | 29 | GATA |
| motif 2 | GRLLPEYRPAASPTFVPSQHSNSHRKVMZ | 29 | - |
| motif 3 | KRLNYPHRVASLMRFREKRKERNFDKKIRYSVRKEVALRMQRRKGQF | 47 | - |
| motif 4 | GGNGGNNRSAALPVALAPPSGSTGGAVRRRRPVPRPRNRQVQRTCS | 46 | - |
| motif 5 | ELYEPSDDLAELEWLSNIMDD | 21 | - |
| motif 6 | CGEDAVRLVGEYGVDAYPFSAQRRRELESMDDARRGGGRLQELLGCEERDYVISADDIKIPIADLAGKTVGLYFGAHWCPPCHVFTKQLKEVYNELKILR | 100 | - |
| motif 7 | KKPNHIIMENGPFSGQNFRKMGDVDPSYRSSSGSAVSYSESCAPYGAADASEMTGSAQSHAWESLVPSRKRSCVTRPKPSPVEKLAKELNFIMHEEKLYY | 100 | - |
| motif 8 | EHPRAMDVLQFPQRWQAYTALRSAGKSVEIIFVSLDRDEASFRDHFQGMSWLAVPFDAAGLLRQKLCARFAIERIPALI | 79 | - |
| motif 9 | EEDLLYHSETPIGSFEIGSGSVLLRHPNSKSLEEESEASSIPADNKSYITSESYSGSASFVIHNGNKAAINLNAPNARPKKSPLHMEDNARRCKLFYERQ | 100 | - |
| motif 10 | MSNQPPHASLQDDLPDCDGDDPLALAIRLFPAHTTGAGLSPAALGIGRVAEPPRREQEPLANSTYGVRGAGPDPWGLRLSRSVLGGLDGFDVDTFFADD | 99 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Y.; Huang, J.; Yu, L.; Wang, C.; Zhang, X.; Cheng, X.; Yu, H.; Zhang, K. Identification, Characterization, and Expression Profiling of Maize GATA Gene Family in Response to Abiotic and Biotic Stresses. Agronomy 2023, 13, 1921. https://doi.org/10.3390/agronomy13071921
Hu Y, Huang J, Yu L, Wang C, Zhang X, Cheng X, Yu H, Zhang K. Identification, Characterization, and Expression Profiling of Maize GATA Gene Family in Response to Abiotic and Biotic Stresses. Agronomy. 2023; 13(7):1921. https://doi.org/10.3390/agronomy13071921
Chicago/Turabian StyleHu, Yuchao, Jingyi Huang, Li Yu, Changjin Wang, Xinwei Zhang, Xinxin Cheng, Haibing Yu, and Kaijing Zhang. 2023. "Identification, Characterization, and Expression Profiling of Maize GATA Gene Family in Response to Abiotic and Biotic Stresses" Agronomy 13, no. 7: 1921. https://doi.org/10.3390/agronomy13071921