

Understanding the Influence of Applying Two Culture Filtrates to Control Gray Mold Disease (Botrytis cinerea) in Tomato


 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains
2.2. In Vitro Plant-Growth-Promoting Activities
2.3. In Vitro Evaluation of T. longibrachiatum and Pseudomonas sp. Culture Filtrates against Botrytis cinerea
2.4. In Vivo Evaluation of T. longibrachiatum, Pseudomonas sp. and/or Salicylic Acid on Tomato Seedlings in the Presence of Botrytis cinerea
2.4.1. Effect of Culture Filtrate on Tomato Seedling Gray Mold Disease Severity
2.4.2. Determination of Phenotypic and Plant Growth Parameters
2.5. In Vivo Evaluation of T. longibrachiatum, Pseudomonas sp. and/or Salicylic Acid on Tomato Fruits Inoculated with Botrytis cinerea
2.5.1. Effect on Gray Mold Severity
2.5.2. Determination of Morphometric Fruit Quality
2.5.3. Determination of Physiochemical Fruit Parameters
2.5.4. Determination of Antioxidant Enzymatic Activities
2.5.5. Determination of Stress Markers
2.6. Statistical Analysis
3. Results
3.1. In Vitro Plant-Growth-Promoting Activities
3.2. In Vitro Evaluation of T. longibrachiatum and Pseudomonas sp. Culture Filtrates against Botrytis cinerea
3.3. In Vivo Evaluation of T. longibrachiatum, Pseudomonas sp., and/or Salicylic Acid on Tomato Seedlings in the Presence of Botrytis cinerea
3.3.1. Effect of Culture Filtrates on Gray Mold Disease Incidence
3.3.2. Effect of Culture Filtrate on Tomato Seedlings’ Phenotype and Growth
3.4. In Vivo Evaluation of T. longibrachiatum, Pseudomonas sp., and/or Salicylic Acid on Tomato Fruits Inoculated with Botrytis cinerea
3.4.1. Effect of Culture Filtrates on Fruit Gray Mold Disease Severity
3.4.2. Effect of Culture Filtrates on Tomato Fruit Morphometric and Quality
3.4.3. Effect of Culture Filtrates on Antioxidant Enzymatic Activities
3.4.4. Effect of Culture Filtrates on Protein, Total Phenols, and MDA Contents
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ali, M.Y.; Sina, A.A.; Khandker, S.S.; Neesa, L.; Tanvir, E.M.; Kabir, A.; Khalil, M.I.; Gan, S.H. Nutritional Composition and Bioactive Compounds in Tomatoes and Their Impact on Human Health and Disease: A Review. Foods 2020, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Zhang, C.; Wu, X.; Liu, L.; Zhang, H. Efficacy assessment of Pantoeajilinensis D25 fermentation broth against Botrytis cinerea. Process Biochem. 2021, 111, 241–248. [Google Scholar] [CrossRef]
- Elad, Y.; Williamson, B.; Tudzynski, P.; Delen, N. Botrytis spp. and diseases they cause in agricultural systems–an introduction. In Botrytis: Biology, Pathology and Control; Springer: Berlin/Heidelberg, Germany, 2007; pp. 1–8. [Google Scholar]
- Elad, Y.; Pertot, I.; Cotes Prado, A.M.; Stewart, A. Plant hosts of Botrytis spp. In Botrytis—The Fungus, the Pathogen and Its Management in Agricultural Systems; Fillinger, S., Elad, Y., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 413–486. [Google Scholar]
- Hua, L.; Yong, C.; Zhanquan, Z.; Boqiang, L.; Guozheng, Q.; Shiping, T. Pathogenic mechanisms and control strategies of Botrytis cinerea causing post-harvest decay in fruits and vegetables. Food Qual. Saf. 2018, 2, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Fillinger, S.; Elad, Y. Botrytis-the Fungus, the Pathogen and Its Management in Agricultural Systems; Springer: New York, NY, USA, 2016. [Google Scholar]
- Adnan, M.; Hamada, M.; Li, G.; Luo, C. Detection and molecular characterization of resistance to the dicarboximide and benzamide fungicides in Botrytis cinerea from tomato in Hubei Province, China. Plant Dis. 2018, 102, 1299–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adnan, M.; Hamada, M.S.; Hahn, M.; Li, G.-Q.; Luo, C.-X. Fungicide resistance of Botrytis cinerea from strawberry to procymidone and zoxamide in Hubei, China. Phytopathol. Res. 2019, 1, 17. [Google Scholar] [CrossRef]
- Zemmouri, B.; Lammoglia, S.-K.; Bouras, F.-Z.; Seghouani, M.; Rebouh, N.Y.; Latati, M. Modelling human health risks from pesticide use in innovative legume-cereal intercropping systems in Mediterranean conditions. Ecotoxicol. Environ. Saf. 2022, 238, 113590. [Google Scholar] [CrossRef]
- Reglinski, T.; Elmer, P.A.G.; Taylor, J.T.; Wood, P.N.; Hoyte, S.M. Inhibition of Botrytis cinerea growth and suppression of botrytis bunch rot in grapes using chitosan. Plant Pathol. 2010, 59, 882–890. [Google Scholar] [CrossRef]
- Massart, S.; Martinez-Medina, M.; Jijakli, M.H. Biological control in the microbiome era: Challenges and opportunities. Biol. Control 2015, 89, 98–108. [Google Scholar] [CrossRef]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Rebouh, N.Y.; Aliat, T.; Polityko, P.M.; Kherchouche, D.; Boulelouah, N.; Temirbekova, S.K.; Afanasyeva, Y.V.; Kucher, D.E.; Plushikov, V.G.; Parakhina, E.A.; et al. Environmentally Friendly Wheat Farming: Biological and Economic Efficiency of Three Treatments to Control Fungal Diseases in Winter Wheat (Triticum aestivum L.) under Field Conditions. Plants 2022, 11, 1566. [Google Scholar] [CrossRef]
- Vos, C.M.; De Cremer, K.; Cammue, B.P.; De Coninck, B. The toolbox of Trichoderma spp. in the biocontrol of Botrytis cinerea disease. Mol. Plant Pathol. 2015, 16, 400–412. [Google Scholar] [CrossRef] [Green Version]
- Santoyo, G.; Orozco-Mosqueda, M.C.; Govindappa, M.; Santoyo, G.; Orozco-mosqueda, M.C.; Govindappa, M. Mechanisms of biocontrol and plant growth-promoting activity in soil bacterial species of Bacillus and Pseudomonas: A review. Biocontr. Sci. Technol. 2012, 22, 855–872. [Google Scholar] [CrossRef]
- Köhl, J.; Goossen-van de Geijn, H.; Groenenboom-de, L.; Haas, B.; Henken, R.; Hauschild, U.; Hilscher, C.; Lombaers-van der Plas, T.; Van den Bosch, M.W. Stepwise screening of candidate antagonists for biological control of Blumeriagraminis f. sp. tritici. Biol. Control 2019, 136, 104008. [Google Scholar] [CrossRef]
- Nygren, K.; Dubey, M.; Zapparata, A.; Iqbal, M.; Tzelepis, G.D.; Durling, M.B.; Jensen, D.F.; Karlsson, M. The mycoparasitic fungus Clonostachys rosea responds with both common and specific gene expression during interspecific interaction with fungal prey. Evol. Appl. 2018, 11, 931–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharyya, C.; Banerjee, S.; Acharya, U.; Mitra, A.; Mallick, I.; Haldar, A.; Haldar, S.; Ghosh, A.; Ghosh, A. Evaluation of plant growth promotion properties and induction of antioxidative defense mechanism by tea rhizobacteria of Darjeeling, India. Sci. Rep. 2020, 10, 15536. [Google Scholar] [CrossRef] [PubMed]
- El-Mageed, T.A.A.; El-Mageed, S.A.A.; El-Saadony, M.T.; Abdelaziz, S.; Abdou, N.M. Plant Growth-Promoting Rhizobacteria Improve Growth, Morph-Physiological Responses, Water Productivity, and Yield of Rice Plants Under Full and Deficit Drip Irrigation. Rice 2022, 15, 16. [Google Scholar] [CrossRef] [PubMed]
- Pérez-García, L.A.; Sáenz-Mata, J.; Fortis-Hernández, M.; Navarro-Muñoz, C.E.; Palacio-Rodríguez, R.; Preciado-Rangel, P. Plant-Growth-Promoting Rhizobacteria Improve Germination and Bioactive Compounds in Cucumber Seedlings. Agronomy 2023, 13, 315. [Google Scholar] [CrossRef]
- Nurbailis, A.; Djamaan, H.; Rahma, Y.I. Potential of culture filtrate from Trichoderma spp. as biofungicide to Colletotrichum gloeosporioides causing anthracnose disease in chili. Biodiversitas 2019, 20, 2915–2920. [Google Scholar] [CrossRef]
- Rhouma, A.; Ben Salem, I.; M’Hamdi, M.; Boughalleb-M’Hamdi, N. Antagonistic potential of certain soilborne fungal bioagents against Monosporascus root rot and vine decline of watermelon and promotion of its growth. Nov. Res. Microbiol. J. 2018, 2, 85–100. [Google Scholar]
- Mohammedi, Z. Etude de Pouvoir Antimicrobien et Antioxydant des Huiles Essentielles et Flavonoides de Quelques Plantes de la Région de Tlemcen. Master’s Thesis, Université Abou BakrBelkaid Tlemcen, Tlemcen, Algeria, 2005; p. 105. [Google Scholar]
- Segarra, G.; Casanova, E.; Borrero, C.; Avilés, M.; Trillas, I. The suppressive effects of composts used as growth media against Botrytis cinerea in cucumber plants. Eur. J. Plant Pathol. 2007, 117, 393–402. [Google Scholar] [CrossRef]
- Rhouma, A.; Mehaoua, M.S.; Mougou, I.; Rhouma, H.; Shah, K.K.; Bedjaoui, H. Combining melon varieties with chemical fungicides for integrated powdery mildew control in Tunisia. Eur. J. Plant Pathol. 2023, 165, 189–201. [Google Scholar] [CrossRef]
- El-Shafey, N.M.; AbdElgawad, H.R. Antioxidants Released from Cichorium pumilum Jacq. Amendment Mitigate Salinity Stress in Maize. Jordan J. Biol. Sci. 2020, 13, 525–533. [Google Scholar]
- Righini, H.; Francioso, O.; Di Foggia, M.; Quintana, A.M.; Roberti, R. Preliminary Study on the Activity of Phycobiliproteins against Botrytis cinerea. Mar. Drugs 2020, 18, 600. [Google Scholar] [CrossRef]
- Rhouma, A.; Khrieba, M.I.; Salih, Y.A.; Rhouma, H.; Bedjaoui, H. Efficacy of fungicides for control of powdery mildew on grapevines in Chott Sidi Abdel Salam oasis, southeastern Tunisia. J. Oasis Agric. Sustain. Dev. 2021, 3, 1–7. [Google Scholar] [CrossRef]
- Geasa, M.M.M.; Hassan, M.H.A. Effect of Mechanical Damage on Tomato Fruits under Storage Conditions. J. Soil Sci. Agric. Eng. 2022, 31, 93–98. [Google Scholar] [CrossRef]
- LópezCamelo, A.F.; Gómez, P.A. Comparison of color indexes for tomato ripening. Hortic. Bras. 2004, 22, 534–537. [Google Scholar] [CrossRef]
- Hajlaoui, F.; Hajlaoui, H. Effect of Exogenous Selenium Intake on Yield and Quality of Tomatoes Grown Under Salt Stress. IOP Conf. Ser. Earth Environ. Sci. 2021, 904, 012047. [Google Scholar] [CrossRef]
- Naeem, M.; Basit, A.; Ahmad, I.; Mohamed, H.I.; Wasila, H. Effect of Salicylic Acid and Salinity Stress on the Performance of Tomato Plants. Gesunde. Pflanzen. 2020, 72, 393–402. [Google Scholar] [CrossRef]
- Mzibra, A.; Aasfar, A.; Khouloud, M.; Farrie, Y.; Boulif, R.; Kadmiri, I.M.; Bamouh, A.; Douira, A. Improving growth, yield, and quality of tomato plants (Solanum lycopersicum L.) by the application of moroccan seaweed-based biostimulants under greenhouse conditions. Agronomy 2021, 11, 1373. [Google Scholar] [CrossRef]
- Velazhahan, R.; Vidhyasekaran, P. Role of phenolic compounds, peroxidase and polyphenol-oxidase in resistance of groundnut to rust. Acta Phytopathol. Entomol. Hung. 1994, 29, 23–29. [Google Scholar]
- Bhuvaneshwari, V.; Amsaveni, R.; Kalaiselvi, M.; Rajeshwari, R.; Paul, P.K. Induced resistance by neem extracts in plants. Int. J. Biosci. Nanosci. 2015, 12, 221–224. [Google Scholar]
- Zhou, X.R.; Xiao, Y.J.; Meng, X.H.; Liu, B.J. Full inhibition of Whangkeumbae pear polyphenol oxidase enzymatic browning reaction by L-cysteine. Food Chem. 2018, 266, 1–8. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Haraguchi, H.; Saito, T.; Okamura, N.; Yagi, A. Inhibition of lipid peroxidation and superoxide generation by diterpenoids from Rosmarinus officinalis. Planta Med. 1995, 61, 333–346. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Zhang, B.; Liu, H.; Han, J.; Zhang, Y. Identification of endophytic Bacillus velezensis ZSY-1 strain and antifungal activity of its volatile compounds against Alternaria solani and Botrytis cinerea. Biol. Control 2017, 105, 27–39. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Gao, Y.; Gao, T.; Zhang, D. Inhibitory Abilities of Bacillus Isolates and Their Culture Filtrates against the Gray Mold Caused by Botrytis cinerea on Postharvest Fruit. Plant Pathol. J. 2019, 35, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Lijiahong, G.; Yalun, F.; Xiaohua, P.; Zhengkun, Y.; Mengke, Z.; Zhiyu, S.; Ning, G.; Shuangchen, C.; Junliang, C.; Bing, B.; et al. Biocontrol potential of Trichoderma harzianum against Botrytis cinerea in tomato plants. Biol. Control 2022, 174, 105019. [Google Scholar] [CrossRef]
- Zhao, X.; Zhou, J.; Tian, R.; Liu, Y. Microbial volatile organic compounds: Antifungal mechanisms, applications, and challenges. Front. Microbiol. 2022, 13, 922450. [Google Scholar] [CrossRef]
- Rosenzweig, J.L.; Weinger, K.; Poirier-Solomon, L.; Rushton, M. Use of a disease severity index for evaluation of healthcare costs and management of comorbidities of patients with diabetes mellitus. Am. J. Manag. Care 2002, 8, 950–958. [Google Scholar] [PubMed]
- Yadav, T.; Kumar, A.; Yadav, R.; Yadav, G.; Kumar, R.; Kushwaha, M. Salicylic acid and thiourea mitigate the salinity and drought stress on physiological traits governing yield in pearl millet-wheat. Saudi J. Biol. Sci. 2020, 27, 2010–2017. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, S. The effects of salicylic acid on the distribution of 14C-assimilation and photosynthesis in young grape plants under heat stress. Int. Symp. Biotechnol. Temp. Fruit Crops Trop. Species 2005, 738, 779–785. [Google Scholar] [CrossRef]
- Shoaib, A.; Munir, M.; Javaid, A.; Awan, A.Z.; Rafiq, M. Anti-mycotic potential of Trichoderma spp. and leaf biomass of Azadirachtaindica against the charcoal rot pathogen Macrophominaphaseolina (Tassi) Goid. in cowpea. Egypt J. Biol. Pest Cont. 2018, 28, 26. [Google Scholar] [CrossRef] [Green Version]
- Hunziker, L.; Bönisch, D.; Groenhagen, U.; Bailly, A.; Schulz, S.; Weisskopf, L. Pseudomonas strains naturally associated with potato plants produce volatiles with a high potential for inhibition of Phytophthora infestans. Appl. Environ. Microbiol. 2015, 81, 821–830. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.S.; Dutta, S.; Ann, M.; Raaijmakers, J.M.; Park, K. Promotion of plant growth by Pseudomonas fluorescens strain SS101 via novel volatile organic compounds. Biochem. Biophys. Res. Commun. 2015, 461, 361–365. [Google Scholar] [CrossRef]
- Boamah, S.; Zhang, S.; Xu, B.; Li, T.; Calderón-Urrea, A. Trichoderma longibrachiatum (TG1) Enhances Wheat Seedlings Tolerance to Salt Stress and Resistance to Fusarium pseudograminearum. Front. Plant Sci. 2021, 12, 741231. [Google Scholar] [CrossRef]
- Ould Amer, S.; Aliat, T.; Kucher, D.E.; Bensaci, O.A.; Rebouh, N.Y. Investigating the Potential of Arbuscular Mycorrhizal Fungi in Mitigating Water Deficit Effects on Durum Wheat (Triticum durum Desf.). Agriculture 2023, 13, 552. [Google Scholar] [CrossRef]
- Abdelmoteleb, A.; Gonzalez-Mendoza, D.; Zayed, O. Cell-free culture filtrate of Trichoderma longibrachiatum AD-1 as an alternative approach to control Fusarium solani and induce defense response Phaseolus vulgaris L. plants. Rhizosphere 2023, 25, 100648. [Google Scholar] [CrossRef]
- Hajji-Hedfi, L.; Hlaoua, W.; Al-Judaibi, A.A.; Rhouma, A.; Horrigue-Raouani, N.; Abdel-Azeem, A.M. Comparative effectiveness of filamentous fungi in biocontrol of Meloidogyne javanica and activated defense mechanisms on tomato. J. Fungi. 2023, 9, 37. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Di Mattia, E.; El-Nakhel, C.; Cardarelli, M. Co-inoculation of Glomus intraradices and Trichoderma atroviride acts as a biostimulant to promote growth, yield and nutrient uptake of vegetable crops. J. Sci. Food Agric. 2015, 95, 1706–1715. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Cisneros, M.F.; Ornelas-Paz, J.J.; Olivas-Orozco, G.I.; Acosta-Muñiz, C.H.; Sepúlveda-Ahumada, D.R.; Pérez-Corral, D.A.; Rios-Velasco, C.; Salas-Marina, M.A.; Fernández-Pavía, S.P. Effect of Trichoderma spp. and phytopathogenic fungi on plant growth and tomato fruit quality. Rev. Mex. Fitopatol. 2018, 36, 444–456. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.P.; Singh, H.B.; Singh, D.K. Effect of Trichoderma harzianum on mineral component and antioxidant activity of tomato fruits. Int. J. Plant Res. 2013, 26, 237–244. [Google Scholar] [CrossRef]
- Vukelić, I.D.; Prokić, L.T.; Racić, G.M.; Pešić, M.B.; Bojović, M.M.; Sierka, E.M.; Kalaji, H.M.; Panković, D.M. Effects of Trichoderma harzianum on photosynthetic characteristics and fruit quality of tomato plants. Int. J. Mol. Sci. 2021, 22, 6961. [Google Scholar] [CrossRef] [PubMed]
- Borrero, C.; Trillas, M.I.; Delgado, A.; Avilés, M. Effect of ammonium/nitrate ratio in nutrient solution on control of Fusarium wilt of tomato by Trichoderma asperellum T34. Plant Pathol. 2012, 61, 132–139. [Google Scholar] [CrossRef]
- Silva-Beltrán, N.P.; Ruiz-Cruz, S.; Cira-Chávez, L.A.; Estrada-Alvarado, M.I.; Ornelas-Paz, J.D.J.; López-Mata, M.A.; Del-Toro-Sánchez, C.L.; Ayala-Zavala, J.F.; Márquez-Ríos, E. Total phenolic, flavonoid, tomatine, and tomatidine contents and antioxidant and antimicrobial activities of extracts of tomato plant. Int. J. Anal. Chem. 2015, 2015, 284071. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.-H.; Park, S.; Park, S.; Oh, B.-J.; Back, K.; Han, O.; Kim, J.-I.; Kim, Y.S. Overexpression of a defensin enhances resistance to a fruit-specific anthracnose fungus in pepper. PLoS ONE 2014, 9, e97936. [Google Scholar] [CrossRef]
- Konappa, N.; Krishnamurthy, S.; Arakere, U.C.; Chowdappa, S.; Ramachandrappa, N.S. Efficacy of indigenous plant growth-promoting rhizobacteria and Trichoderma strains in eliciting resistance against bacterial wilt in a tomato. Egypt J. Biol. Pest. Control 2020, 30, 106. [Google Scholar] [CrossRef]
- Oszust, K.; Cybulska, J.; Frąc, M. How do Trichoderma genus fungi win a nutritional competition battle against soft fruit pathogens? A report on niche overlap nutritional potentiates. Int. J. Mol. Sci. 2020, 21, 4235. [Google Scholar] [CrossRef]
- Gholamnezhad, J.; Sanjarian, F.; Goltapeh, E.M.; Safaei, N.; Razavi, K.H. Effect of salicylic acid on enzyme activity in wheat on immediate early time after infection with Mycosphaerella graminicola. Plant Sci. 2016, 47, 1–8. [Google Scholar] [CrossRef] [Green Version]


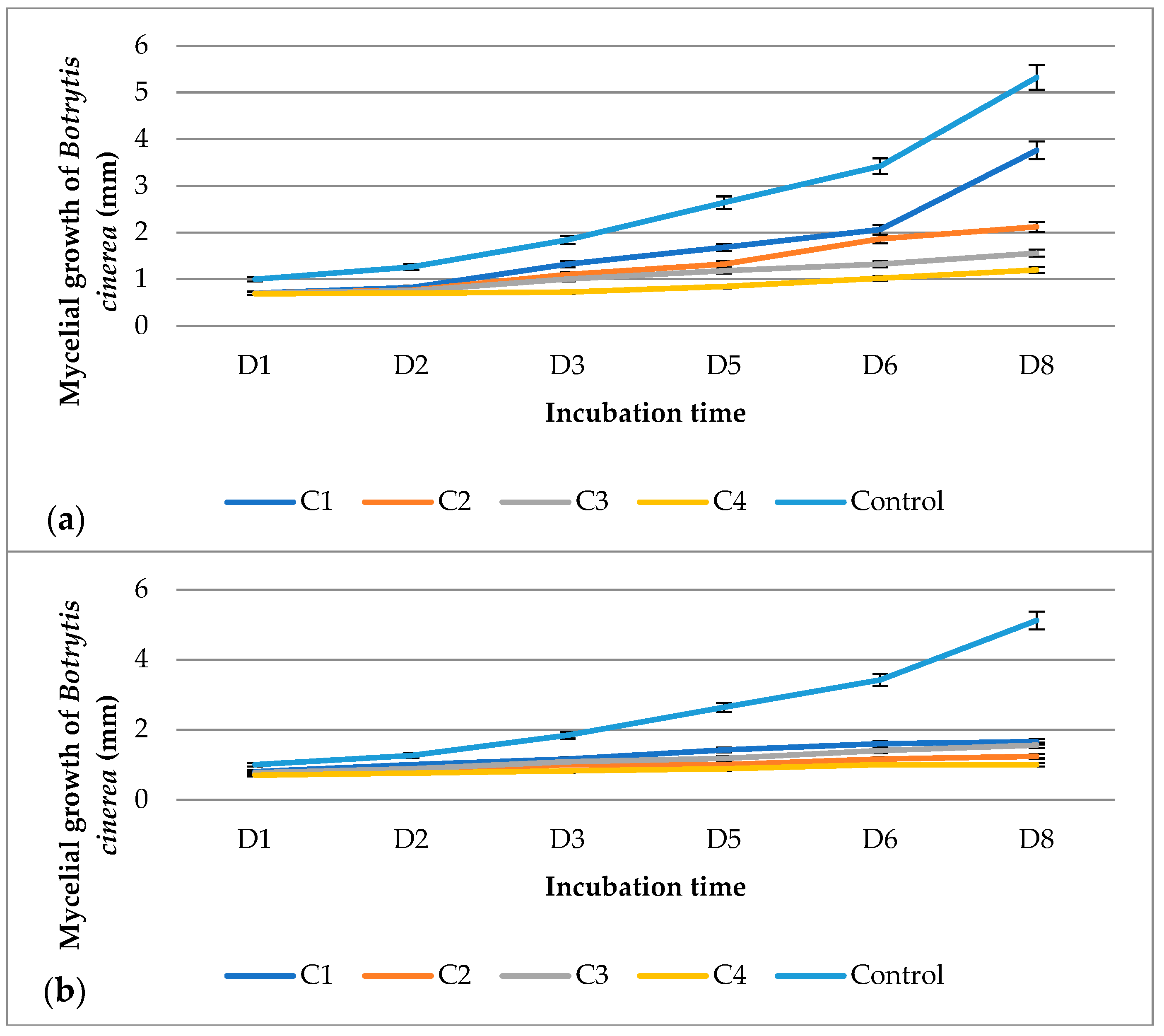{kind=link}
| Microorganism | Cat | Pec | Amy | Pro | Chit | Glu | AIA | HCN | N | P |
|---|---|---|---|---|---|---|---|---|---|---|
| T. longibrachiatum | + | - | - | + | + | + | + | + | - | + |
| Pseudomonas sp. | + | - | + | - | - | - | + | + | - | + |
| Treatments | Mycelial Growth Rate (mm h−1) | Mycelial Growth Inhibition (%) | ||
|---|---|---|---|---|
| T. longibrachiatum | Pseudomonas sp. | T. longibrachiatum | Pseudomonas sp. | |
| C1 | 1.37 ± 0.03 b a | 1.01 ± 0.07 b | 41.80 ± 0.62 d | 67.58 ± 0.24 b |
| C2 | 1.15 ± 0.01 c | 0.85 ± 0.12 bc | 57.67 ± 0.32 c | 75.78 ± 0.57 ab |
| C3 | 0.99 ± 0.04 d | 0.97 ± 0.14 bc | 68.52 ± 0.54 b | 69.53 ± 0.39 ab |
| C4 | 0.83 ± 0.004 e | 0.80 ± 0.03 c | 79.63 ± 0.81 a | 80.47 ± 0.90 a |
| Control | 2.14 ± 0.04 a | 1.81 ± 0.03 a | nd | nd |
| p-value b | <0.01 | <0.01 | <0.01 | <0.01 |
| Treatments | DSI | Resistance Level | Chl a (mg g−1 Fresh Weight) | Chl b (mg g−1 Fresh Weight) | Chl T (mg g−1 Fresh Weight) | Fresh Weight (g) | Seedling Length (cm) |
|---|---|---|---|---|---|---|---|
| Negative control | 0 ± 0 f a | EE | 6.13 ± 0.66 a | 0.85 ± 0.24 cd | 6.98 ± 0.41 a | 0.61 ± 0.09 a | 13.18 ± 0.85 a |
| Positive control | 92.60 ± 1.71 a | HI | 1.66 ± 0.11 de | 0.68 ± 0.04 de | 2.34 ± 0.09 g | 0.17 ± 0.06 c | 7.08 ± 0.81 e |
| T. longibrachiatum filtrate | 9.40 ± 1.52 e | E | 4.08 ± 0.02 b | 2.43 ± 0.02 b | 6.52 ± 0.02 b | 0.69 ± 0.09 a | 11.5 ± 0.61 b |
| Pseudomonas sp. filtrate | 10.80 ± 1.38 e | E | 2.13 ± 0.01 c | 0.92 ± 0.01 c | 3.05 ± 0.01 e | 0.44 ± 0.07 b | 11.6 ± 1.14 b |
| Salicylic acid | 37.80 ± 1.59 b | I | 2.10 ± 0.02 cd | 0.63 ± 0.009 e | 2.73 ± 0.02 f | 0.38 ± 0.08 b | 11.38 ± 0.44 bc |
| T. longibrachiatum filtrate + Salicylic acid | 15.60 ± 1.05 d | E | 1.36 ± 0.03 e | 2.34 ± 0.02 b | 3.70 ± 0.01 d | 0.26 ± 0.04 c | 9.54 ± 0.79 d |
| Pseudomonas sp. filtrate + Salicylic acid | 22.20 ± 1.48 c | E | 2.56 ± 0.02 c | 2.72 ± 0.03 a | 5.28 ± 0.01 c | 0.43 ± 0.08 b | 10.26 ± 1.21 cd |
| p-value b | <0.01 | nd | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Chl a | Chl b | Chl T | SL | FWS | |
|---|---|---|---|---|---|
| Chl b | −0.068 | ||||
| Chl T | 0.868 ** | 0.436 * | |||
| SL | 0.679 ** | 0.022 | 0.624 ** | ||
| FWS | 0.585 ** | 0.327 | 0.690 ** | 0.773 ** | |
| DSI | −0.521 * | −0.385 | −0.661 ** | −0.822 ** | −0.796 ** |
| Treatments | PFA | DSI | Resistance Level |
|---|---|---|---|
| Negative control | 0 ± 0 e a | 0 ± 0 e | EE |
| Positive control | 92.99 ± 0.88 a | 94.37 ± 0.12 a | HE |
| T. longibrachiatum filtrate | 24.52 ± 0.30 c | 23.1 ± 0.29 c | E |
| Pseudomonas sp. filtrate | 33.21 ± 0.78 b | 27.31 ± 0.25 bc | I |
| Salicylic acid | 40 ± 0.51 b | 31.97 ± 0.08 b | I |
| T. longibrachiatum filtrate + Salicylic acid | 13.97 ± 0.33 d | 13.01 ± 0.56 d | E |
| Pseudomonas sp. filtrate + Salicylic acid | 23.29 ± 0.07 c | 17.51 ± 0.15 d | E |
| p-value b | <0.01 | <0.01 | Nd |
| Treatments | Fruit Firmness | pH | Electrical Conductivity (mS cm−1) | Water Content (%) | Juice Yield (%) | Titratable Acidity (g/10 mL Juice) | Sugar Content (°Brix) | Color Density | |
|---|---|---|---|---|---|---|---|---|---|
| L | a/b | ||||||||
| Negative control | 3.07 ± 0.15 a a | 3.59 ± 0.01 b | 2.14 ± 0.23 d | 98.15 ± 0.10 f | 43.47 ± 0.05 e | 1.18 ± 0.02 a | 6.27 ± 0.15 a | 39.47 ± 0.75 a | 1.09 ± 0.07 a |
| Positive control | 0 ± 0 e | 4.37 ± 0.55 a | 2.63 ± 0.19 a | 99.59 ± 0.5 a | 30.66 ± 0.15 g | 0.89 ± 0.01 d | 4.93 ± 0.11 c | 41.80 ± 0.60 a | 1.23 ± 0.05 a |
| T. longibrachiatum filtrate | 2.1 ± 0.17 c | 3.74 ± 0.01 b | 2.57 ± 0.02 b | 98.43 ± 0.01 e | 69.22 ± 0.05 a | 0.83 ± 0.01 e | 5.30 ± 0.43 c | 33.87 ± 0.49 b | 1.16 ± 0.13 a |
| Pseudomonas sp. filtrate | 2.5 ± 0.28 b | 3.55 ± 0.01 b | 2.34 ± 0.11 c | 98.96 ± 0.05 d | 56.56 ± 0.05 b | 0.83 ± 0.01 e | 5.87 ± 0.05 b | 36.33 ± 0.53 b | 0.87 ± 0.10 b |
| Salicylic acid | 1.7 ± 0.05 d | 3.73 ± 0.02 b | 1.76 ± 0.01 g | 99.42 ± 0.02 b | 53.92 ± 0.06 d | 1.18 ± 0.02 a | 5.07 ± 0.11 c | 34.87 ± 0.23 b | 1.14 ± 0.12 a |
| T. longibrachiatum filtrate + Salicylic acid | 2.33 ± 0.10 b | 3.74 ± 0.05 b | 1.92 ± 0.01 f | 97.65 ± 0.04 g | 39.68 ± 0.11 f | 1.02 ± 0.01 c | 4.97 ± 0.06 c | 35.33 ± 0.36 b | 0.89 ± 0.08 b |
| Pseudomonas sp. filtrate + Salicylic acid | 3.03 ± 0.06 a | 3.56 ± 0.06 b | 2.12 ± 0.05 e | 99.13 ± 0.04 c | 56.02 ± 0.05 c | 1.08 ± 0.05 b | 5.97 ± 0.21 ab | 36.40 ± 0.29 b | 1.19 ± 0.17 a |
| p-value b | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Treatments | POX (Units mg−1 min−1) | PPO (Units mg−1 min−1) | CAT (µmol H2O2 mg protein−1) | APX (µmol mg−1 min−1) | TPC (µg g−1) | TP (mg g−1) | MDA (µmol g−1) |
|---|---|---|---|---|---|---|---|
| Negative control | 4.06 ± 0.08 ab a | 10.92 ± 0.03 e | 44.58 ± 0.23 a | 39.39 ± 0.92 b | 5.48 ± 0.27 a | 8.45 ± 0.47 c | 1.75 ± 0.02 b |
| Positive control | 1.96 ± 0.24 b | 19.03 ± 0.15 b | 32.12 ± 0.15 e | 15.78 ± 0.25 f | 2.88 ± 0.06 d | 37.13 ± 0.13 a | 5.73 ± 0.04 a |
| T. longibrachiatum filtrate | 3.79 ± 0.07 b | 15.15 ± 0.21 c | 38.36 ± 0.09 c | 32.61 ± 0.57 c | 4.58 ± 0.08 bc | 12.64 ± 0.93 b | 2.28 ± 0.98 b |
| Pseudomonas sp. filtrate | 3.76 ± 0.07 b | 13.74 ± 0.52 d | 36.32 ± 0.10 d | 25.68 ± 0.63 d | 4.60 ± 0.34 bc | 12.33 ± 0.39 b | 2.79 ± 0.16 b |
| Salicylic acid | 3.78 ± 0.28 b | 19.93 ± 0.11 a | 36.28 ± 0.24 d | 44.17 ± 0.45 a | 4.82 ± 0.42 bc | 11.94 ± 0.26 b | 2.65 ± 0.01 b |
| T. longibrachiatum filtrate + Salicylic acid | 4.17 ± 0.13 a | 14.75 ± 0.09 c | 43.84 ± 0.18 ab | 30.37 ± 0.73 c | 4.35 ± 0.29 c | 12.64 ± 0.38 b | 2.62 ± 0.71 b |
| Pseudomonas sp. filtrate + Salicylic acid | 4.16 ± 0.11 a | 7.22 ± 0.57 f | 42.56 ± 0.09 b | 22.82 ± 0.75 e | 4.94 ± 0.05 b | 8.99 ± 0.35 c | 1.90 ± 0.04 b |
| p-value b | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| APX | POX | PPO | TPC | MDA | TP | FF | pH | EC | WC | JY | TA | Brix | L | a/b | PFA | DSI | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CAT | 0.360 | 0.793 ** | −0.712 | 0.675 ** | −0.629 ** | −0.725 ** | 0.817 ** | −0.556 ** | −0.481 * | −0.757 ** | 0.076 | 0.520 * | 0.461 * | −0.258 | −0.202 | −0.877 ** | −0.848 ** |
| APX | 0.573 ** | 0.141 | 0.659 ** | −0.500 * | −0.626 ** | 0.407 | −0.428 | −0.639 ** | −0.311 | 0.324 | 0.604 ** | 0.104 | −0.441 * | −0.062 | −0.589 ** | −0.610 ** | |
| POX | −0.577 ** | 0.815 ** | −0.753 ** | −0.965 ** | 0.911 ** | −0.751 ** | −0.614 ** | −0.589 ** | 0.522 * | 0.401 | 0.455 * | −0.627 ** | −0.298 | −0.921 ** | −0.960 ** | ||
| PPO | −0.547 * | 0.509 * | 0.569 ** | −0.795 ** | 0.538 * | 0.056 | 0.328 | −0.241 | −0.179 | −0.765 ** | 0.043 | 0.039 | 0.613 ** | 0.611 ** | |||
| TPC | −0.785 ** | −0.895 ** | 0.873 ** | −0.730 ** | −0.529 * | −0.396 | 0.505 * | 0.538 * | 0.635 ** | −0.408 | −0.247 | −0.868 ** | −0.886 ** | ||||
| MDA | 0.791 ** | −0.754 ** | 0.639 ** | 0.414 | 0.431 | −0.505 * | −0.348 | −0.567 ** | 0.468 * | 0.332 | 0.755 ** | 0.775 ** | |||||
| TP | −0.925 ** | 0.812 ** | 0.581 ** | 0.492 * | −0.612 ** | −0.415 | −0.537 * | 0.620 ** | 0.286 | 0.910 ** | 0.954 ** | ||||||
| FF | −0.796 ** | −0.433 * | −0.542 * | 0.477 * | 0.355 | 0.699 ** | −0.427 | −0.293 | −0.928 ** | −0.945 ** | |||||||
| pH | 0.415 | 0.347 | −0.522 * | −0.249 | −0.517 * | 0.497 * | 0.285 | 0.714 ** | 0.781 ** | ||||||||
| EC | 0.212 | 0.027 | −0.795 ** | −0.014 | 0.363 | 0.242 | 0.485 * | 0.538 * | |||||||||
| WC | −0.015 | −0.084 | −0.136 | 0.269 | 0.443 * | 0.748 ** | 0.685 ** | ||||||||||
| JY | −0.207 | 0.252 | −0.721 ** | 0.006 | −0.410 | −0.465 * | |||||||||||
| TA | 0.222 | 0.022 | 0.135 | −0.397 | −0.419 | ||||||||||||
| Brix | 0.147 | −0.127 | −0.539 * | −0.547 * | |||||||||||||
| L | 0.273 | 0.465 * | 0.527 * | ||||||||||||||
| a/b | 0.321 | 0.327 | |||||||||||||||
| PFA | 0.982 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hajji-Hedfi, L.; Rhouma, A.; Hajlaoui, H.; Hajlaoui, F.; Rebouh, N.Y. Understanding the Influence of Applying Two Culture Filtrates to Control Gray Mold Disease (Botrytis cinerea) in Tomato. Agronomy 2023, 13, 1774. https://doi.org/10.3390/agronomy13071774
Hajji-Hedfi L, Rhouma A, Hajlaoui H, Hajlaoui F, Rebouh NY. Understanding the Influence of Applying Two Culture Filtrates to Control Gray Mold Disease (Botrytis cinerea) in Tomato. Agronomy. 2023; 13(7):1774. https://doi.org/10.3390/agronomy13071774
Chicago/Turabian StyleHajji-Hedfi, Lobna, Abdelhak Rhouma, Hichem Hajlaoui, Fedi Hajlaoui, and Nazih Y. Rebouh. 2023. "Understanding the Influence of Applying Two Culture Filtrates to Control Gray Mold Disease (Botrytis cinerea) in Tomato" Agronomy 13, no. 7: 1774. https://doi.org/10.3390/agronomy13071774