

Effect of Irrigation Water Salinity on Soil Characteristics and Microbial Communities in Cotton Fields in Southern Xinjiang, China
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
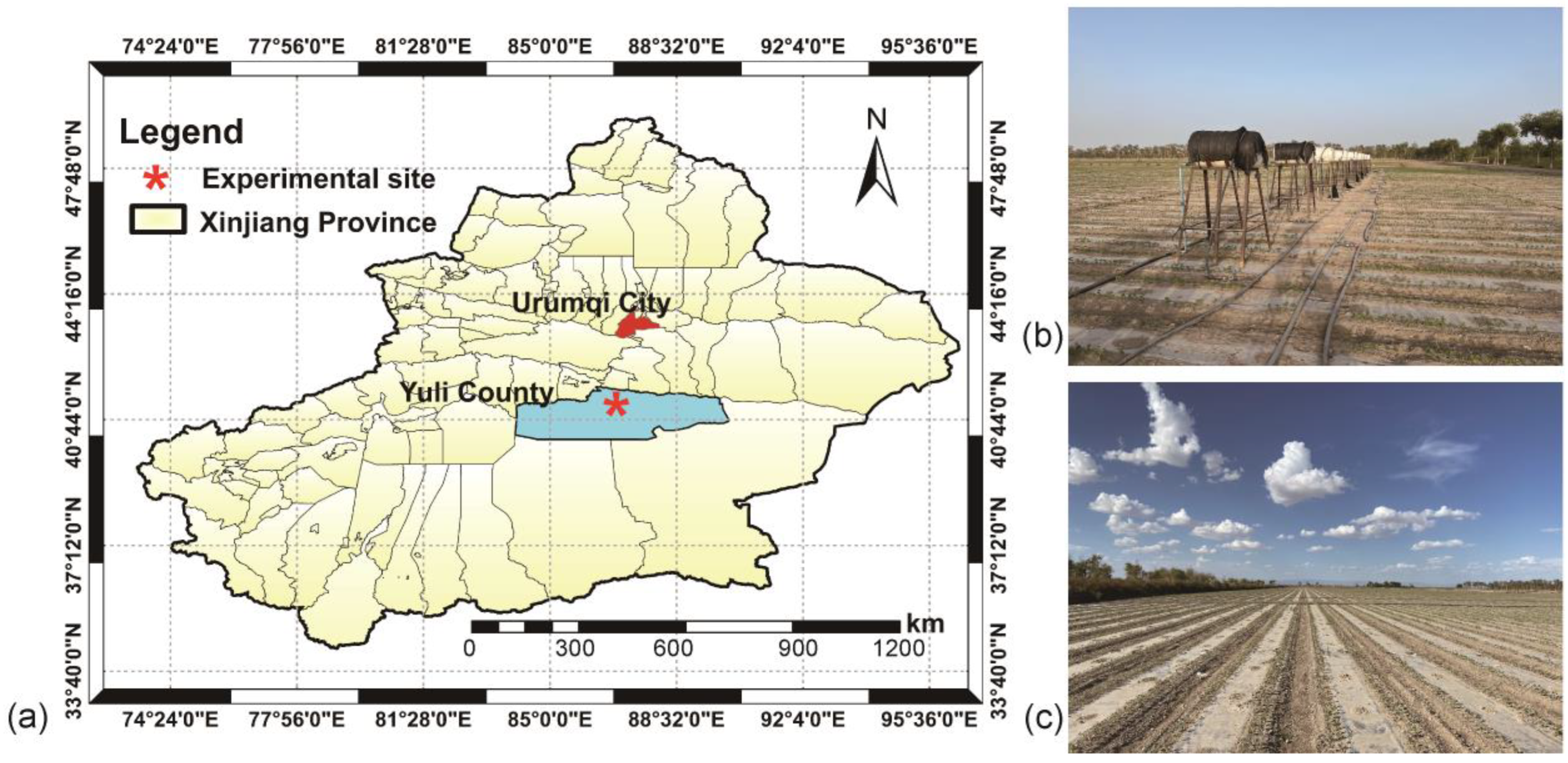
2.1. Study Site and Experimental Setup
2.2. Soil Sampling
2.3. Soil Physicochemical Analysis
2.4. Hydro-Chemical Ions
2.5. Microbial Biomass and Enzymatic Assay
2.6. Cotton Seed Yield and Biomass
2.7. Measurement of Soil Microbial Communities
2.8. Data Tools and Statistical Analyses
3. Results
3.1. Soil Physicochemical Properties
3.2. Microbial Biomass and Enzymatic Activity
3.3. The Richness and Diversity of Bacterial and Fungal Communities
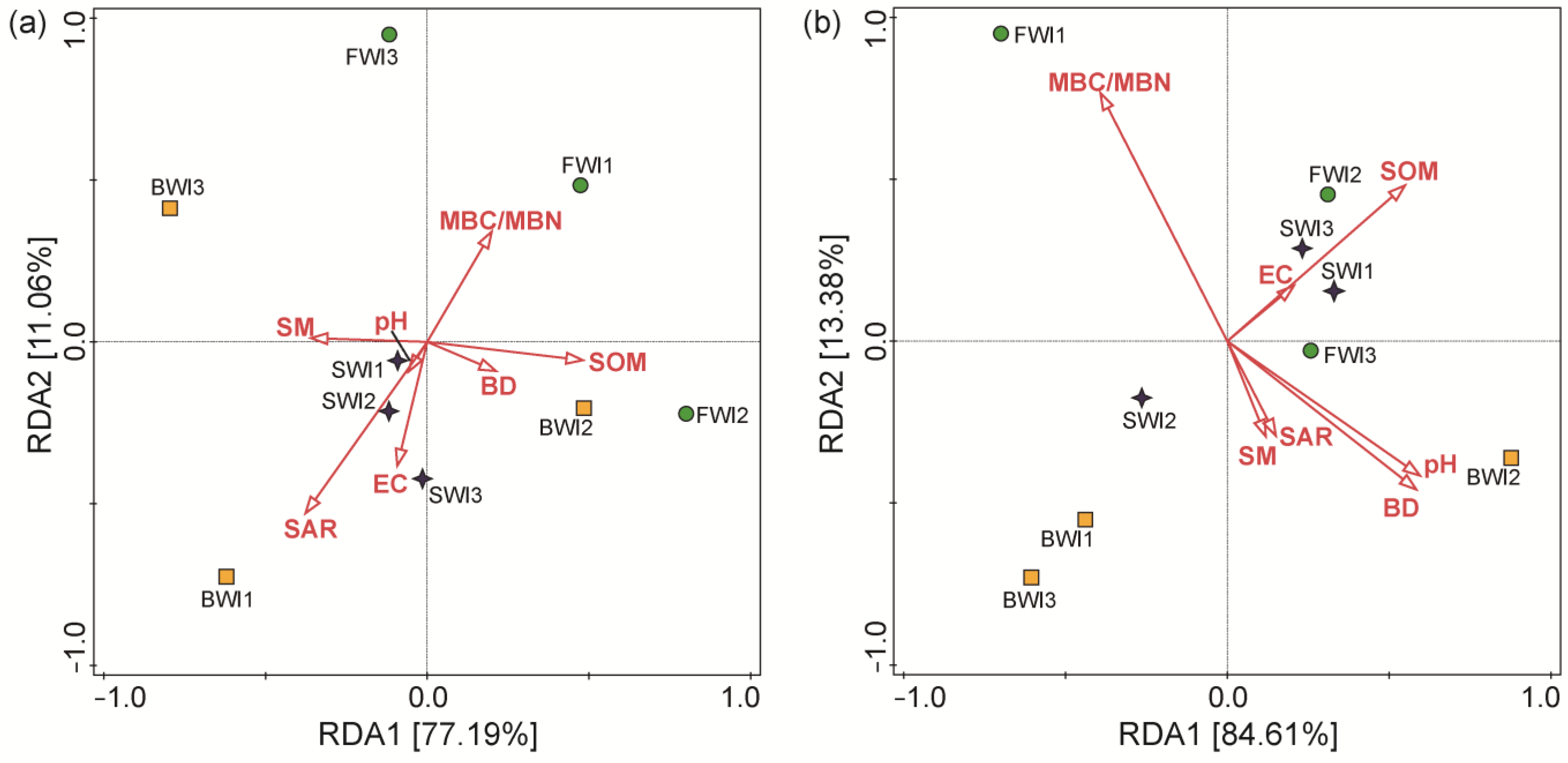
3.4. Microbial Community Composition
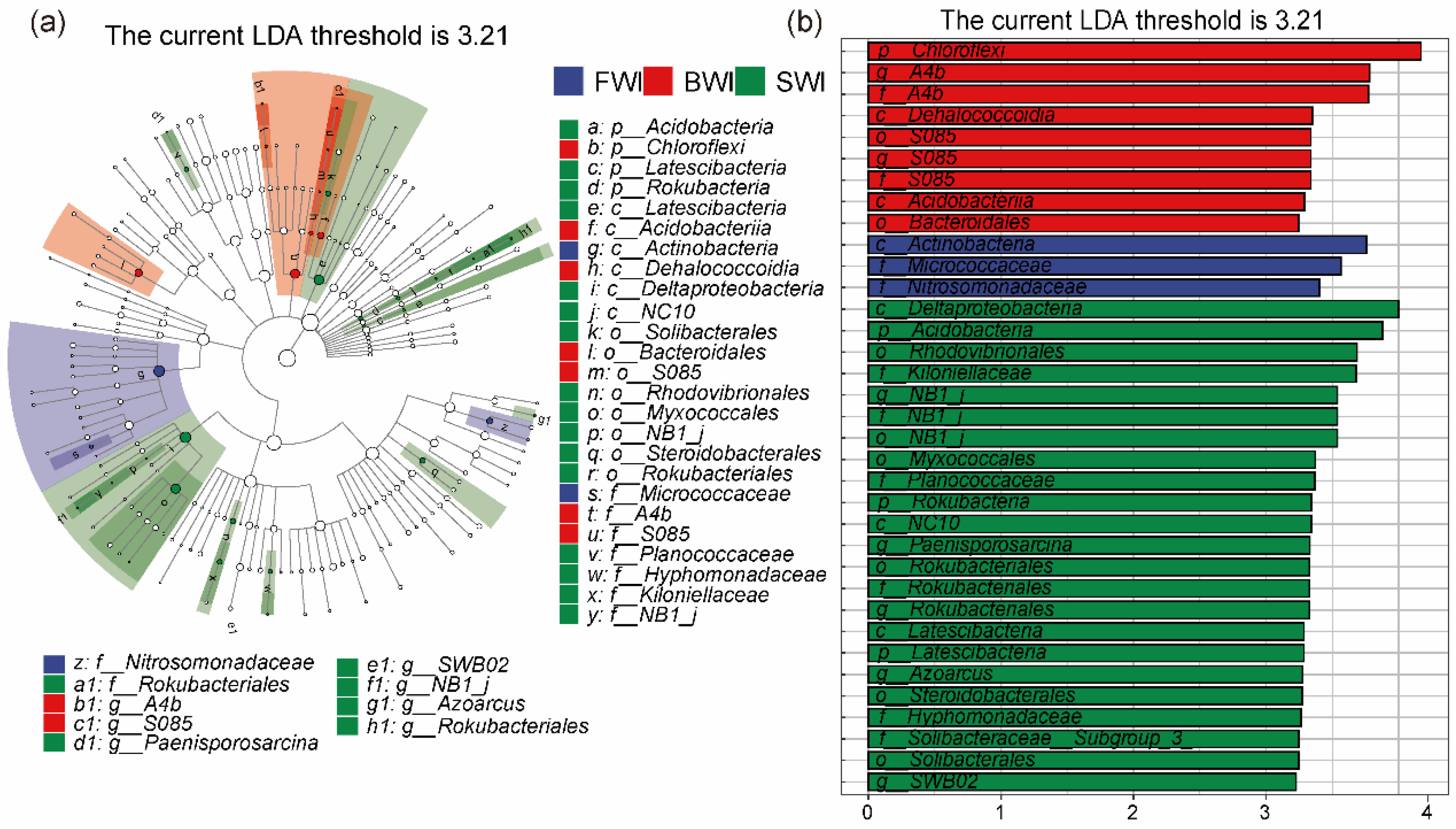
3.5. Analysis of Biomarkers in Microbial Communities
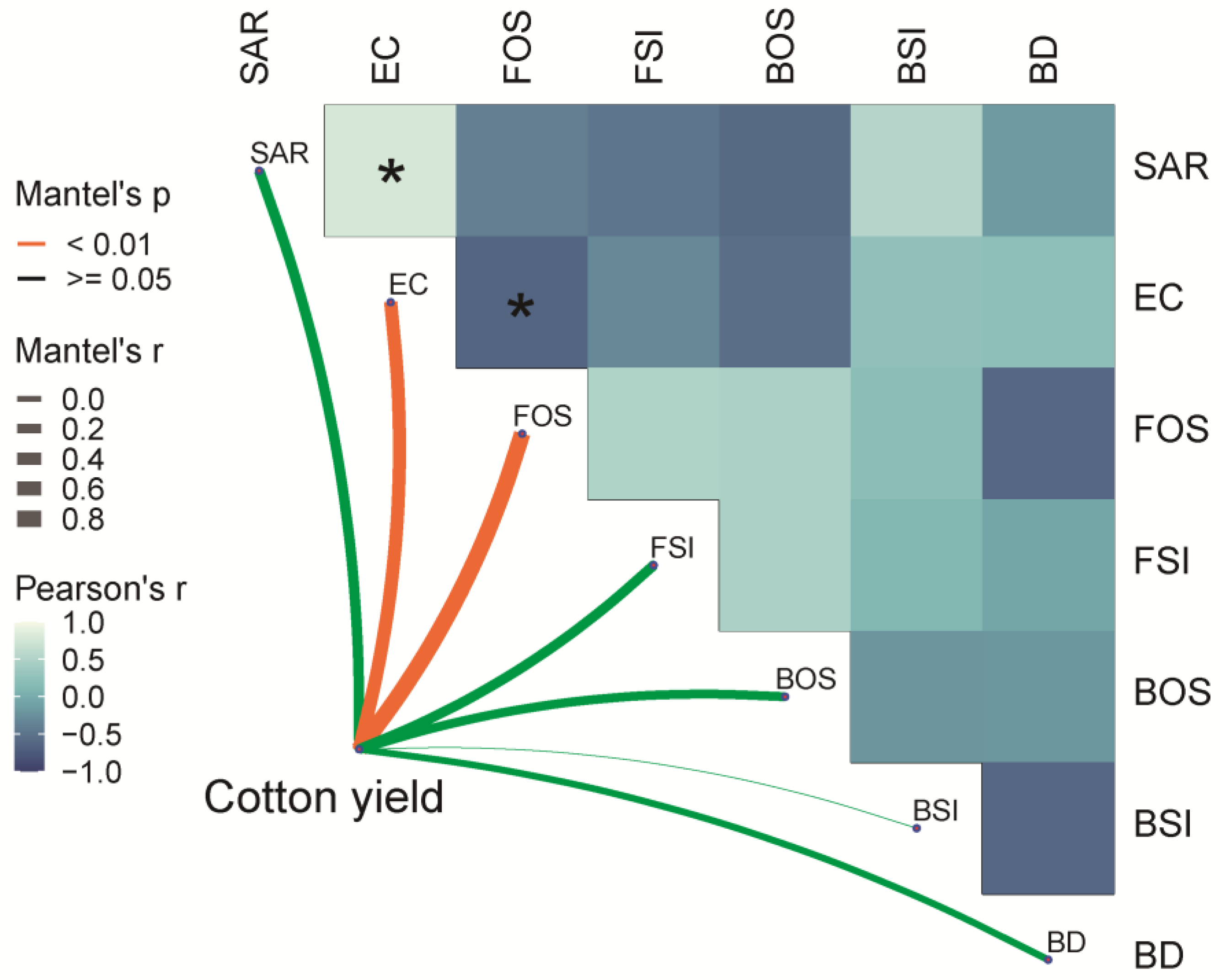
3.6. Correlation Analysis of Yield Index with Soil Physicochemical Properties and Microbial Community
4. Discussion
4.1. Effect of Saline Water Irrigation on Physicochemical Characteristics and Biological Properties of Soil
4.2. Effect of Irrigation with Saline Water on Bacterial and Fungal Communities
4.3. Relationship between Microorganisms and Soil Physicochemical Properties
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lambers, H. Introduction, dryland salinity: A key environmental issue in southern Australia. Plant Soil 2003, 257, 5–7. [Google Scholar] [CrossRef]
- Chen, W.; Jin, M.; Ferré, T.P.A.; Liu, Y.; Xian, Y.; Shan, T.; Ping, X. Spatial distribution of soil moisture, soil salinity, and root density beneath a cotton field under mulched drip irrigation with brackish and fresh water. Field Crop. Res. 2018, 215, 207–221. [Google Scholar] [CrossRef]
- Li, X.; Jin, M.; Zhou, N.; Huang, J.; Jiang, S.; Telesphore, H. Evaluation of evapotranspiration and deep percolation under mulched drip irrigation in an oasis of Tarim basin, China. J. Hydrol. 2016, 538, 677–688. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Kang, Y.; Wan, S.; Hu, W.; Liu, S.; Liu, S. Salt distribution and the growth of cotton under different drip irrigation regimes in a saline area. Agric. Water Manag. 2011, 100, 58–69. [Google Scholar] [CrossRef]
- Cheng, M.; Wang, H.; Fan, J.; Wang, X.; Sun, X.; Yang, L.; Zhang, S.; Xiang, Y.; Zhang, F. Crop yield and water productivity under salty water irrigation: A global meta-analysis. Agric. Water Manag. 2021, 256, 107105. [Google Scholar] [CrossRef]
- Chunxia, W.; Guang, Y.; Junfeng, L.; Xinlin, H.; Lianqing, X.; Aihua, L. Effects of timing and duration under brackish water mulch drip irrigation on cotton yield in northern Xinjiang, China. Int. J. Agric. Biol. Eng. 2017, 10, 115–122. [Google Scholar] [CrossRef]
- Yang, G.; Li, F.; Tian, L.; He, X.; Gao, Y.; Wang, Z.; Ren, F. Soil physicochemical properties and cotton (Gossypium hirsutum L.) yield under brackish water mulched drip irrigation. Soil Tillage Res. 2020, 199, 104592. [Google Scholar] [CrossRef]
- Gong, X.; Liu, C.; Li, J.; Luo, Y.; Yang, Q.; Zhang, W.; Yang, P.; Feng, B. Responses of rhizosphere soil properties, enzyme activities and microbial diversity to intercropping patterns on the Loess Plateau of China. Soil Tillage Res. 2019, 195, 104355. [Google Scholar] [CrossRef]
- Zhang, H.; Luo, G.; Wang, Y.; Fei, J.; Xiangmin, R.; Peng, J.; Tian, C.; Zhang, Y. Crop rotation-driven change in physicochemical properties regulates microbial diversity, dominant components, and community complexity in paddy soils. Agric. Ecosyst. Environ. 2023, 343, 108278. [Google Scholar] [CrossRef]
- Chen, J.; Liu, X.; Zheng, J.; Zhang, B.; Lu, H.; Chi, Z.; Pan, G.; Li, L.; Zheng, J.; Zhang, X.; et al. Biochar soil amendment increased bacterial but decreased fungal gene abundance with shifts in community structure in a slightly acid rice paddy from Southwest China. Appl. Soil Ecol. 2013, 71, 33–44. [Google Scholar] [CrossRef]
- Hall, R.M.; Penke, N.; Kriechbaum, M.; Kratschmer, S.; Jung, V.; Chollet, S.; Guernion, M.; Nicolai, A.; Burel, F.; Fertil, A.; et al. Vegetation management intensity and landscape diversity alter plant species richness, functional traits and community composition across European vineyards. Agr. Syst. 2020, 177, 102706. [Google Scholar] [CrossRef]
- Liu, H.; Xu, W.; Li, J.; Yu, Z.; Zeng, Q.; Tan, W.; Mi, W. Short-term effect of manure and straw application on bacterial and fungal community compositions and abundances in an acidic paddy soil. J. Soils Sediments 2021, 21, 3057–3071. [Google Scholar] [CrossRef]
- Selim, T.; Berndtsson, R.; Persson, M.; Somaida, M.; El-Kiki, M.; Hamed, Y.; Mirdan, A.; Zhou, Q. Influence of geometric design of alternate partial root-zone subsurface drip irrigation (APRSDI) with brackish water on soil moisture and salinity distribution. Agric. Water Manag. 2012, 103, 182–190. [Google Scholar] [CrossRef]
- Li, X.; Jin, M.; Huang, J.; Yuan, J. The soil–water flow system beneath a cotton field in arid north-west China, serviced by mulched drip irrigation using brackish water. Hydrogeol. J. 2015, 23, 35–46. [Google Scholar] [CrossRef]
- Qi, Z.; Feng, H.; Zhao, Y.; Zhang, T.; Yang, A.; Zhang, Z. Spatial distribution and simulation of soil moisture and salinity under mulched drip irrigation combined with tillage in an arid saline irrigation district, northwest China. Agric. Water Manag. 2018, 201, 219–231. [Google Scholar] [CrossRef]
- Wang, R.; Wan, S.; Sun, J.; Xiao, H. Soil salinity, sodicity and cotton yield parameters under different drip irrigation regimes during saline wasteland reclamation. Agric. Water Manag. 2018, 209, 20–31. [Google Scholar] [CrossRef]
- Wichern, J.; Wichern, F.; Joergensen, R.G. Impact of salinity on soil microbial communities and the decomposition of maize in acidic soils. Geoderma 2006, 137, 100–108. [Google Scholar] [CrossRef]
- Dong, Y.; Zhang, J.; Chen, R.; Zhong, L.; Lin, X.; Feng, Y. Microbial community composition and activity in saline soils of coastal agro-ecosystems. Microorganisms 2022, 10, 835–846. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Li, X.; Jin, M.; Wang, R.; Chen, J.; Guo, S. Reduced co-occurrence and ion-specific preferences of soil microbial hub species after ten years of irrigation with brackish water. Soil Tillage Res. 2020, 199, 104599. [Google Scholar] [CrossRef]
- Chowdhury, N.; Nakatani, A.S.; Setia, R.; Marschner, P. Microbial activity and community composition in saline and non-saline soils exposed to multiple drying and rewetting events. Plant Soil 2011, 348, 103–113. [Google Scholar] [CrossRef]
- Chaudhry, V.; Rehman, A.; Mishra, A.; Chauhan, P.S.; Nautiyal, C.S. Changes in bacterial community structure of agricultural land due to long-term organic and chemical amendments. Microb. Ecol. 2012, 64, 450–460. [Google Scholar] [CrossRef]
- Yan, N.; Marschner, P. Response of soil respiration and microbial biomass to changing EC in saline soils. Soil Biol. Biochem. 2013, 65, 322–328. [Google Scholar] [CrossRef]
- Zhalnina, K.; Dias, R.; de Quadros, P.D.; Davis-Richardson, A.; Camargo, F.A.O.; Clark, I.M.; McGrath, S.P.; Hirsch, P.R.; Triplett, E.W. Soil pH Determines Microbial Diversity and Composition in the Park Grass Experiment. Microb. Ecol. 2015, 69, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Soman, C.; Li, D.; Wander, M.M.; Kent, A.D. Long-term fertilizer and crop-rotation treatments differentially affect soil bacterial community structure. Plant Soil 2017, 413, 145–159. [Google Scholar] [CrossRef]
- Zhang, K.; Shi, Y.; Cui, X.; Yue, P.; Li, K.; Liu, X.; Tripathi, B.M.; Chu, H.; Lozupone, C. Salinity is a key determinant for soil microbial communities in a desert ecosystem. mSystems 2019, 4, e218–e225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oren, A. The bioenergetic basis for the decrease in metabolic diversity at increasing salt concentrations: Implications for the functioning of salt lake ecosystems. Hydrobiologia 2001, 466, 61–72. [Google Scholar] [CrossRef]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef]
- Kabiri, V.; Raiesi, F.; Ghazavi, M.A. Tillage effects on soil microbial biomass, SOM mineralization and enzyme activity in semi-arid Calcixerepts. Agric. Ecosyst. Environ. 2016, 232, 73–84. [Google Scholar] [CrossRef]
- Yang, P.; Zia-Khan, S.; Wei, G.; Zhong, R.; Aguila, M. Winter Irrigation Effects in Cotton Fields in Arid Inland Irrigated Areas in the North of the Tarim Basin, China. Water 2016, 8, 47. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Li, Y.; Si, B.; Wang, Y.; Chen, X.; Wang, X.; Chen, H.; Wang, H.; Zhang, F.; Bai, Y.; et al. Optimizing biochar application to improve soil physical and hydraulic properties in saline-alkali soils. Sci. Total Environ. 2021, 771, 144802. [Google Scholar] [CrossRef]
- Carter, D.L.; Mortland, M.M.; Kemper, W.D. Methods of Soil Analysis Part I: Physical and Mineralogical Methods; Amer Society of Agronomy: Madison, WI, USA, 1986. [Google Scholar]
- Bao, S.D. Soil Agricultural Chemistry Analysis; Agriculture Press: Beijing, China, 1999. [Google Scholar]
- Gillman, G.P.; Sumpter, E.A. Modification to the compulsive exchange method for measuring exchange characteristics of soils. Aust. J. Soil. Res. 1986, 24, 61–66. [Google Scholar] [CrossRef]
- Brady, N.C.; Weil, R.R.; Brady, N.C. Elements of Nature and Properties of Soils; Prentice Hall: Hoboken, NJ, USA, 2000. [Google Scholar]
- Yan, M.; Li, T.; Li, X.; Liu, Y.; Zhang, J. Microbial biomass and activity restrict soil function recovery of a post-mining land in eastern Loess Plateau. Catena 2021, 199, 105107. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Gordon, K. Supplement 1: Microbes and Health || Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; He, Z.; Shi, W. Cotton/mung bean intercropping improves crop productivity, water use efficiency, nitrogen uptake, and economic benefits in the arid area of Northwest China. Agric. Water Manag. 2020, 240, 106277. [Google Scholar] [CrossRef]
- Li, Y.; Yao, N.; Liang, J.; Wang, X.; Niu, B.; Jia, Y.; Jiang, F.; Yu, Q.; Liu, D.L.; Feng, H.; et al. Rational biochar application rate for cotton nutrient content, growth, yields, productivity, and economic benefits under film-mulched trickle irrigation. Agric. Water Manag. 2023, 276, 108079. [Google Scholar] [CrossRef]
- Liang, J.; Shi, W. Cotton/halophytes intercropping decreases salt accumulation and improves soil physicochemical properties and crop productivity in saline-alkali soils under mulched drip irrigation: A three-year field experiment. Field Crop. Res. 2021, 262, 108027. [Google Scholar] [CrossRef]
- Bellemain, E.; Carlsen, T.; Brochmann, C.; Coissac, E.; Kauserud, H. Its as an environmental DNA barcode for fungi: An in silico approach reveals potential PCR biases. BMC Microbiol. 2010, 10, 189–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Caporaso, J.G. QIIME 2: Reproducible, interactive, scalable, and extensible microbiome data science. PeerJ 2018, 25, 16–70. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 11–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Kaehler, B.D.; Ram, R.J.; Matthew, D.; Evan, B.; Rob, K.; Huttley, G.A.; Gregory, C.J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2′s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Guo, H.; Shi, X.; Ma, L.; Yang, T.; Min, W. Long-term irrigation with saline water decreases soil nutrients, diversity of bacterial communities, and cotton yields in a gray desert soil in China. Pol. J. Environ. Stud. 2020, 29, 4077–4088. [Google Scholar] [CrossRef]
- Zhang, A.; Zheng, C.; Li, K.; Dang, H.; Cao, C.; Rahma, A.E.; Zhang, J.; Feng, D. Responses of soil water-salt variation and cotton growth to drip irrigation with saline water in the low plain near the Bohai Sea. Irrig. Drain. 2020, 69, 448–459. [Google Scholar] [CrossRef]
- Li, X.; Jin, M.; Zhou, N.; Jiang, S.; Hu, Y. Inter-dripper variation of soil water and salt in a mulched drip irrigated cotton field: Advantages of 3-D modelling. Soil Tillage Res. 2018, 184, 186–194. [Google Scholar] [CrossRef]
- Singh, K. Microbial and enzyme activities of saline and sodic soils. Land Degrad. Dev. 2015, 27, 706–718. [Google Scholar] [CrossRef]
- Doongar, R.; Chaudhary, A.P.; Rathore, B.J. Effects of seawater irrigation on soil microbial community structure and physiological function. Int. J. Environ. Sci. Technol. 2016, 13, 2199–2208. [Google Scholar] [CrossRef] [Green Version]
- Zahran, H.H. Diversity, adaptation and activity of the bacterial flora in saline environments. Biol. Fertil. Soils 1997, 25, 211–223. [Google Scholar] [CrossRef]
- Zhai, H.; Cao, C.; Liu, M. Effects of long-term brackish water irrigation on soil enzyme activity and reaction kinetics. Agric. Water Manag. 2018, 36, 95–101, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Bowles, T.M.; Acosta-Martínez, V.; Calderón, F.; Jackson, L.E. Soil enzyme activities, microbial communities, and carbon and nitrogen availability in organic agroecosystems across an intensively-managed agricultural landscape. Soil Biol. Biochem. 2014, 68, 252–262. [Google Scholar] [CrossRef]
- Rath, K.M.; Rousk, J. Salt effects on the soil microbial decomposer community and their role in organic carbon cycling: A review. Soil Biol. Biochem. 2015, 81, 108–123. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, L.; Yang, J.; Duan, Y.; Luo, Y.; Taherzadeh, M.J.; Li, Y.; Li, H.; Awasthi, M.K.; Zhao, Z. The diversity of microbial community and function varied in response to different agricultural residues composting. Sci. Total Environ. 2020, 715, 136983. [Google Scholar] [CrossRef]
- Yan, N.; Marschner, P. Response of microbial activity and biomass to increasing salinity depends on the final salinity, not the original salinity. Soil Biol. Biochem. 2012, 53, 50–55. [Google Scholar] [CrossRef]
- He, H.; Wang, Z.; Guo, L.; Zheng, X.; Zhang, J.; Li, W.; Fan, B. Distribution characteristics of residual film over a cotton field under long-term film mulching and drip irrigation in an oasis agroecosystem. Soil Tillage Res. 2018, 180, 194–203. [Google Scholar] [CrossRef]
- Wang, Q.; Shan, Y. Review of research development on water and soil regulation with brackish water irrigation. J. Agric. Mach. 2015, 46, 117–126. [Google Scholar] [CrossRef]
- Chen, W.; Jin, M.; Ferré, T.P.A.; Liu, Y.; Huang, J.; Xian, Y. Soil conditions affect cotton root distribution and cotton yield under mulched drip irrigation. Field Crop. Res. 2020, 249, 107743. [Google Scholar] [CrossRef]
- Singh, G.; Mukerji, K. Root Exudates as Determinant of Rhizospheric Microbial Biodiversity. In Microbial Activity in the Rhizoshere; Mukerji, K.G., Manoharachary, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Gao, Y.; Cui, J.; Ren, G.; Wei, S.; Yang, P.; Yin, C.; Liang, H.; Chang, J. Changes in the root-associated bacteria of sorghum are driven by the combined effects of salt and sorghum development. Environ. Microbiome 2021, 16, 14. [Google Scholar] [CrossRef] [PubMed]
- Eisenhauer, N.; Scheu, S.; Jousset, A. Bacterial diversity stabilizes community productivity. PLoS ONE 2012, 7, e34517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naether, A.; Foesel, B.U.; Naegele, V.; Wust, P.K.; Weinert, J.; Bonkowski, M.; Alt, F.; Oelmann, Y.; Polle, A.; Lohaus, G. Environmental factors affect Acidobacterial communities below the subgroup level in grassland and forest soils. Appl. Environ. Microb. 2012, 78, 7398. [Google Scholar] [CrossRef] [Green Version]
- Yin, C.; Hulbert, S.H.; Schroeder, K.L.; Mavrodi, O.; Mavrodi, D.; Dhingra, A.; Schillinger, W.F.; Paulitz, T.C. Role of Bacterial Communities in the Natural Suppression of Rhizoctonia solani Bare Patch Disease of Wheat (Triticum aestivum L.). Appl. Environ. Microb. 2013, 79, 7428–7438. [Google Scholar] [CrossRef] [Green Version]
- Paungfoo-Lonhienne, C.; Yeoh, Y.K.; Kasinadhuni, N.; Lonhienne, T.; Robinson, N.; Hugenholtz, P.; Ragan, M.A.; Schmidt, S. Nitrogen fertilizer dose alters fungal communities in sugarcane soil and rhizosphere. Sci. Rep. 2015, 5, 8678. [Google Scholar] [CrossRef] [Green Version]
- Couturier, M.; Tangthirasunun, N.; Xie, N.; Brun, S.; Berrin, J.G. Plant biomass degrading ability of the coprophilic ascomycete fungus Podospora anserina. Biotechnol. Adv. 2016, 34, 976–983. [Google Scholar] [CrossRef]
- Setia, R.; Marschner, P.; Baldock, J.; Chittleborough, D.; Smith, P.; Smith, J. Salinity effects on carbon mineralization in soils of varying texture. Soil Biol. Biochem. 2011, 43, 1908–1916. [Google Scholar] [CrossRef]
- Kakumanu, M.L.; Williams, M.A. Osmolyte dynamics and microbial communities vary in response to osmotic more than matric water deficit gradients in two soils. Soil Biol. Biochem. 2014, 79, 14–24. [Google Scholar] [CrossRef]
- Ma, B.; Gong, J. A meta-analysis of the publicly available bacterial and archaeal sequence diversity in saline soils. World J. Microb. Biotechnol. 2013, 29, 2325–2334. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Encinas, C.; Neria-Gonzalez, I.; Alcantara-Hernandez, R.J.; Estrada-Alvarado, I.; Zavala-Diaz, D.L.S.F.; Dendooven, L.; Marsch, R. Changes in the bacterial populations of the highly alkaline saline soil of the former lake Texcoco (Mexico) following flooding. Extremophiles 2009, 13, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Chen, M.; Feng, H.; Wei, M.; Song, F.; Lou, Y.; Cui, X.; Wang, H.; Zhuge, Y. Organic and inorganic fertilizers respectively drive bacterial and fungal community compositions in a fluvo-aquic soil in northern China. Soil Tillage Res. 2020, 198, 104540. [Google Scholar] [CrossRef]
- Muneer, M.A.; Huang, X.; Hou, W.; Zhang, Y.; Cai, Y.; Munir, M.Z.; Wu, L.; Zheng, C. Response of fungal diversity, community composition, and functions to nutrients management in red soil. J. Fungi 2021, 7, 554–572. [Google Scholar] [CrossRef]
- Huang, X.; Muneer, M.A.; Li, J.; Hou, W.; Ma, C.; Jiao, J.; Cai, Y.; Chen, X.; Wu, L.; Zheng, C. Integrated nutrient management significantly improves pomelo (citrus grandis) root growth and nutrients uptake under acidic soil of southern China. Agronomy 2021, 11, 1231. [Google Scholar] [CrossRef]
- Ding, J.; Jiang, X.; Guan, D.; Zhao, B.; Ma, M.; Zhou, B.; Cao, F.; Yang, X.; Li, L.; Li, J. Influence of inorganic fertilizer and organic manure application on fungal communities in a long-term field experiment of Chinese Mollisols. Appl. Soil Ecol. 2017, 111, 114–122. [Google Scholar] [CrossRef]
- Rath, K.M.; Maheshwari, A.; Bengtson, P.; Rousk, J. Comparative Toxicities of Salts on Microbial Processes in Soil. Appl. Environ. Microb. 2016, 82, 2012–2020. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.; Xue, D.; Li, X.; Deng, Y.; Rui, J.; Feng, K.; Wang, Z. The responses and adaptations of microbial communities to salinity in farmland soils: A molecular ecological network analysis. Appl. Soil Ecol. 2017, 120, 239–246. [Google Scholar] [CrossRef]
- Lau, J.A.; Lennon, J.T. Rapid responses of soil microorganisms improve plant fitness in novel environments. Proc. Natl. Acad. Sci. USA 2012, 109, 14058–14062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shade, A.; Peter, H.; Allison, S.D.; Baho, D.L.; Berga, M.; Bürgmann, H.; Huber, D.H.; Langenheder, S.; Lennon, J.T.; Martiny, J.B.H.; et al. Fundamentals of microbial community resistance and resilience. Front. Microbiol. 2012, 3, 417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rath, K.M.; Maheshwari, A.; Rousk, J. Linking Microbial Community Structure to Trait Distributions and Functions Using Salinity as an Environmental Filter. mBio 2019, 10, e1607–e1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, H.; Lili, Y.; Cecilie, H.; Baojian, W.; Ji, C.; Li, Z.; Jiawen, Y.; Xinlin, H. Linking microbial community compositions to cotton nitrogen utilization along soil salinity gradients. Field Crop. Res. 2022, 288, 108697. [Google Scholar] [CrossRef]
- Zhou, Z.; Hua, J.; Xue, J. Salinity drives shifts in soil microbial community composition and network complexity along vegetation community succession in coastal tidal flats. Estuar. Coast. Shelf Sci. 2022, 276, 108005. [Google Scholar] [CrossRef]
- Navarro, A.; Elia, A.; Conversa, G.; Campi, P.; Mastrorilli, M. Potted mycorrhizal carnation plants and saline stress: Growth, quality and nutritional plant responses. Sci. Hortic. 2012, 140, 139–148. [Google Scholar] [CrossRef]
- Gómez-Bellot, M.J.; Ortuño, M.F.; Nortes, P.A.; Vicente-Sánchez, J.; Bañón, S.; Sánchez-Blanco, M.J. Mycorrhizal euonymus plants and reclaimed water: Biomass, water status and nutritional responses. Sci. Hortic. 2015, 186, 61–69. [Google Scholar] [CrossRef]
- Nie, S.; Lei, X.; Zhao, L.; Brookes, P.C.; Wang, F.; Chen, C.; Yang, W.; Xing, S. Fungal communities and functions response to long-term fertilization in paddy soils. Appl. Soil Ecol. 2018, 130, 251–258. [Google Scholar] [CrossRef]
- Fontaine, S.; Henault, C.; Aamor, A.; Bdioui, N.; Bloor, J.M.G.; Maire, V.; Mary, B.; Revaillot, S.; Maron, P.A. Fungi mediate long term sequestration of carbon and nitrogen in soil through their priming effect. Soil Biol. Biochem. 2011, 43, 86–96. [Google Scholar] [CrossRef]
- Swift, M.J.; Heal, O.W.; Anderson, J.M. Decomposition in terrestrial ecosystems. Stud. Ecol. 1979, 5, 2772–2774. [Google Scholar] [CrossRef]
- Zhang, X.; Fu, G.; Xing, S.; Fu, W.; Liu, X.; Wu, H.; Zhou, X.; Ma, Y.; Zhang, X.; Chen, B. Structure and diversity of fungal communities in long-term copper-contaminated agricultural soil. Sci. Total Environ. 2022, 806, 151302. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, J.; Zhao, B.; Yan, P.; Zhou, G.; Xin, X. Effects of straw amendment and moisture on microbial communities in Chinese fluvo-aquic soil. J. Soils Sediments 2014, 14, 1829–1840. [Google Scholar] [CrossRef]
- Wei, K.; Zhang, J.; Wang, Q.; Guo, Y.; Mu, W. Irrigation with ionized brackish water affects cotton yield and water use efficiency. Ind. Crop. Prod. 2022, 175, 114244. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Layer (cm) | Soil Texture | Bulk Density (g cm−3) | Field Capacity (%) | Soil Salinity (g kg−1) | pH (1:5) | Ions (g L−1) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| K+ | Ca2+ | Na+ | Mg2+ | CO32− | Cl− | HCO32− | SO42− | ||||||
| 0–20 | Sandy loam | 1.59 | 21.75 | 3.67 | 8.33 | 0.08 | 0.08 | 0.90 | 0.10 | 0.01 | 1.14 | 0.69 | 0.49 |
| 20–40 | Silt loam | 1.58 | 16.78 | 3.26 | 8.37 | 0.07 | 0.09 | 0.72 | 0.13 | 0.02 | 0.86 | 0.67 | 0.57 |
| 40–60 | Sandy loam | 1.63 | 19.89 | 3.07 | 8.34 | 0.07 | 0.09 | 0.67 | 0.12 | 0.01 | 0.85 | 0.54 | 0.58 |
| 60–80 | Sandy | 1.64 | 12.20 | 2.32 | 8.32 | 0.08 | 0.08 | 0.52 | 0.06 | 0.01 | 0.64 | 0.56 | 0.26 |
| Year | Treatment | EC | TDS | K+ | Ca2+ | Na+ | Mg2+ | CO32− | Cl− | HCO32− | SO42− |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2019 | FWI | 0.12 | 1.00 | 0.02 ± 0.002 | 0.07 ± 0.01 | 0.15 ±0.02 | 0.04 ± 0.01 | - | 0.32 ± 0.05 | 0.22 ± 0.02 | 0.13 ± 0.04 |
| BWI | 0.56 | 3.00 | 0.06 ± 0.011 | 0.14 ± 0.01 | 0.68 ± 0.08 | 0.21 ± 0.03 | - | 0.96 ± 0.03 | 0.38 ± 0.03 | 0.54 ± 0.06 | |
| SWI | 1.57 | 7.00 | 0.14 ± 0.018 | 0.11 ± 0.07 | 1.68 ± 0.13 | 0.33 ± 0.01 | - | 2.77 ± 0.08 | 0.31 ± 0.10 | 1.62 ± 0.07 | |
| 2020 | FWI | 0.12 | 1.00 | 0.01 ± 0.003 | 0.05 ± 0.01 | 0.17 ± 0.01 | 0.06 ± 0.01 | - | 0.31 ± 0.05 | 0.21 ± 0.02 | 0.19 ±0.02 |
| BWI | 0.56 | 3.00 | 0.06 ± 0.009 | 0.14 ± 0.01 | 0.63 ± 0.07 | 0.26 ± 0.02 | - | 1.01 ± 0.02 | 0.36 ± 0.03 | 0.51 ± 0.08 | |
| SWI | 1.57 | 7.00 | 0.10 ± 0.016 | 0.16 ± 0.01 | 1.67 ± 0.15 | 0.28 ± 0.03 | - | 2.76 ± 0.08 | 0.47 ± 0.08 | 1.52 ± 0.15 | |
| 2021 | FWI | 0.12 | 1.00 | 0.02 ± 0.002 | 0.07 ± 0.01 | 0.16 ± 0.01 | 0.06 ± 0.01 | - | 0.28 ± 0.05 | 0.26 ± 0.02 | 0.16 ±0.03 |
| BWI | 0.56 | 3.00 | 0.04 ± 0.015 | 0.14 ± 0.01 | 0.68 ± 0.09 | 0.28 ± 0.02 | - | 0.96 ± 0.03 | 0.41 ± 0.02 | 0.48 ± 0.08 | |
| SWI | 1.57 | 7.00 | 0.09 ± 0.015 | 0.16 ± 0.01 | 1.55 ± 0.22 | 0.29 ± 0.02 | - | 2.78 ± 0.05 | 0.49 ± 0.08 | 1.60 ± 0.10 |
| Date | TN | TDS | K+ | Ca2+ | Na+ | Mg2+ | CO32− | Cl− | HCO32− | SO42− | SAR | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2019 | Jun | - | 7.45 | 0.02 | 0.11 | 1.79 | 0.33 | 0.02 | 2.47 | 0.31 | 2.39 | 3.82 |
| Jul | - | 7.76 | 0.03 | 0.16 | 1.98 | 0.34 | - | 2.72 | 0.62 | 1.89 | 3.96 | |
| Aug | - | 8.86 | 0.01 | 0.20 | 2.13 | 0.68 | 0.02 | 3.20 | 0.54 | 2.07 | 3.21 | |
| 2020 | Jun | - | 10.68 | 0.02 | 0.32 | 3.14 | 0.53 | - | 4.65 | 0.33 | 1.70 | 4.82 |
| Jul | - | 8.37 | 0.02 | 0.37 | 2.38 | 0.41 | - | 3.44 | 0.37 | 1.39 | 3.81 | |
| Aug | - | 7.53 | 0.01 | 0.34 | 2.06 | 0.31 | - | 3.13 | 0.35 | 1.32 | 3.61 | |
| 2021 | Jun | 2.79 | 11.97 | 0.06 | 0.58 | 3.65 | 0.45 | - | 5.05 | 0.35 | 1.83 | 5.09 |
| Jul | - | 9.98 | 0.07 | 0.44 | 3.01 | 0.45 | - | 4.11 | 0.35 | 1.55 | 4.51 | |
| Aug | - | 11.13 | 0.07 | 0.55 | 3.39 | 0.47 | - | 4.62 | 0.34 | 1.70 | 4.75 | |
| Variable | Unit | Treatment | ||
|---|---|---|---|---|
| FWI | BWI | SWI | ||
| TN | g kg−1 | 0.42 ab | 0.38 b | 0.46 a |
| AP | mg kg−1 | 12.79 a | 12.97 a | 14.30 a |
| AK | mg kg−1 | 180.60 a | 188.20 a | 236.17 a |
| SOM | g kg−1 | 7.01 ab | 6.30 b | 7.71 a |
| SOC | g kg−1 | 4.07 ab | 3.66 b | 4.47 a |
| SM | % | 16.31 a | 16.52 a | 17.85 a |
| pH | 8.11 a | 8.25 a | 8.26 a | |
| EC | mS cm−1 | 0.93 b | 1.03 b | 3.18 a |
| K+ | g kg−1 | 0.07 b | 0.06 b | 0.12 a |
| Ca2+ | g kg−1 | 0.53 a | 0.27 b | 0.58 a |
| Na+ | g kg−1 | 0.69 c | 0.92 b | 1.94 a |
| Mg2+ | g kg−1 | 0.23 b | 0.19 c | 0.42 a |
| CO32− | g kg−1 | - | - | - |
| HCO3− | g kg−1 | 0.15 c | 0.20 a | 0.16 b |
| Cl− | g kg−1 | 0.89 c | 1.17 b | 2.49 a |
| SO42− | g kg−1 | 1.12 b | 0.56 c | 1.34 a |
| SAR | 1.11 c | 1.92 b | 2.75 a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, B.; Bai, Y.; Guo, S.; He, Z.; Wang, B.; Liu, H.; Zhai, J.; Cao, H. Effect of Irrigation Water Salinity on Soil Characteristics and Microbial Communities in Cotton Fields in Southern Xinjiang, China. Agronomy 2023, 13, 1679. https://doi.org/10.3390/agronomy13071679
Ding B, Bai Y, Guo S, He Z, Wang B, Liu H, Zhai J, Cao H. Effect of Irrigation Water Salinity on Soil Characteristics and Microbial Communities in Cotton Fields in Southern Xinjiang, China. Agronomy. 2023; 13(7):1679. https://doi.org/10.3390/agronomy13071679
Chicago/Turabian StyleDing, Bangxin, Yungang Bai, Shuchen Guo, Zijian He, Bei Wang, Hongbo Liu, Jiangrui Zhai, and Hongxia Cao. 2023. "Effect of Irrigation Water Salinity on Soil Characteristics and Microbial Communities in Cotton Fields in Southern Xinjiang, China" Agronomy 13, no. 7: 1679. https://doi.org/10.3390/agronomy13071679