
Quantifying and Disentangling the Competition Effect of a Weed Community in a Long-Term Biennial Cereal-Legume Rotation


Abstract
:1. Introduction
2. Material and Methods
2.1. Study Site and Experimental Information
2.2. Population Dynamic Models
2.3. Model Fitting
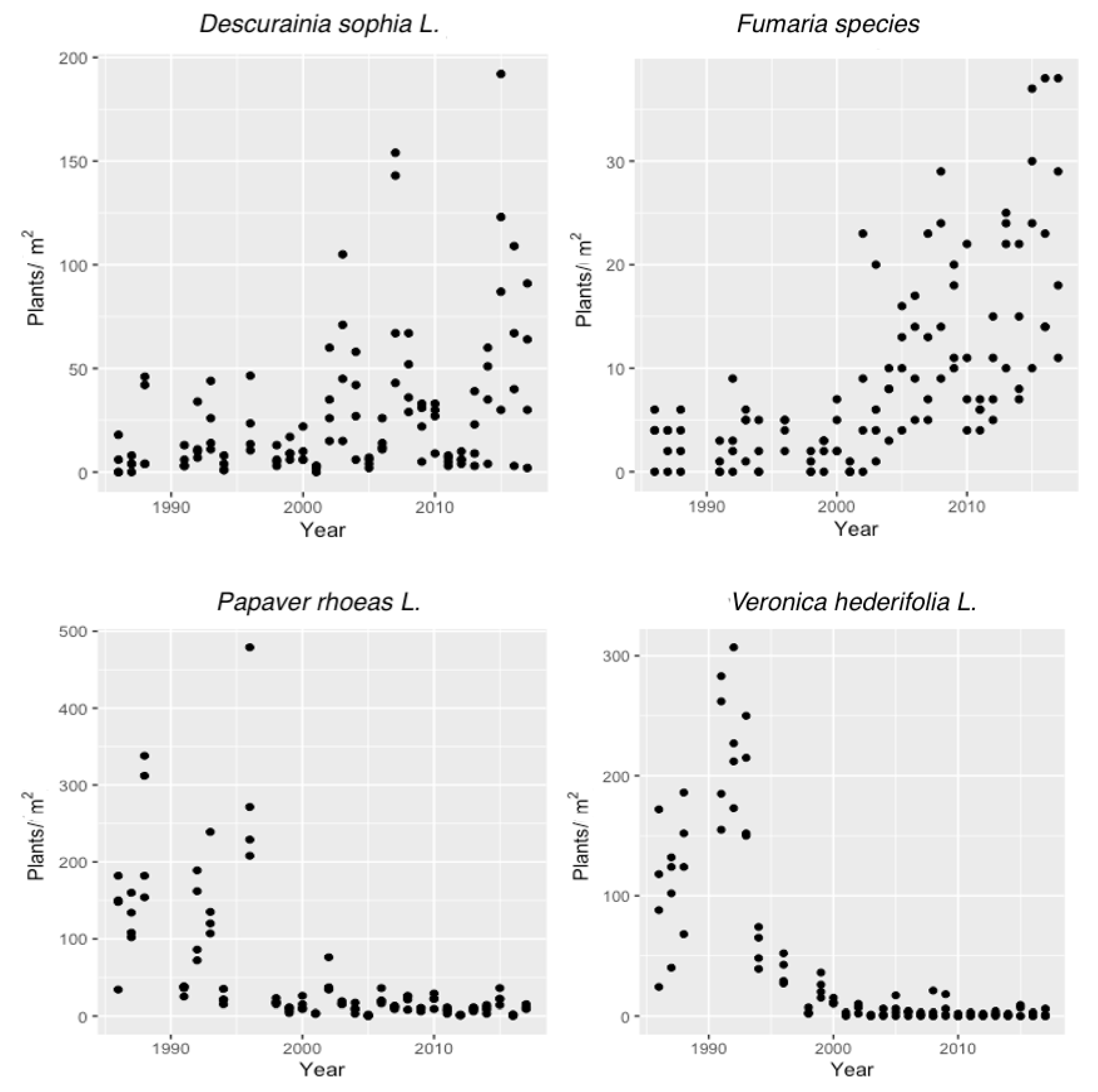
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oerke, E. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Appleby, A.P.; Muller, F.; Carpy, S. Weed control. In Agrochemicals; Muller, F., Ed.; Wiley: New York, NY, USA, 2000; pp. 687–707. [Google Scholar]
- Zimdahl, R. Weed-Crop Competition: A Review; Blackwell Publishing: Oxford, UK, 2004. [Google Scholar]
- Swanton, C.; Nkoa, R.; Blackshaw, R. Experimental Methods for Crop–Weed Competition Studies. Weed Sci. 2015, 63, 2–11. [Google Scholar] [CrossRef]
- Torner, C.; Sanchez del Arco, M.J.; Satorre, E.H.; Fernandez-Quintanilla, C.C. A comparison of the growth patterns and the competitive ability of four annual weeds. Agronomie 2000, 20, 147–156. [Google Scholar] [CrossRef]
- Storkey, J.; Westbury, D.B. Managing arable weeds for biodiversity. Pest Manag. Sci. 2007, 65, 517–523. [Google Scholar] [CrossRef]
- Aarts, H.F.M.; De Visser, C.L.M. A management information system for weed control in winter wheat. Proc. Br. Crop Prot. Conf. Weeds 1985, 2, 679–686. [Google Scholar]
- Hakansson, A. Growth in plant stands of different density. In Proceedings of the VIIIth International Symposium on the Biology, Ecology and Systematics of Weeds, Kyoto, Japan, 10–14 July 1989; pp. 631–640. [Google Scholar]
- Wilson, B.J. Yield responses of winter cereals to the control of bread leaf weeds. In Proceedings of the EWRS Symposium on Economic Weed Control, Wageningen, The Netherlands, 25–28 June 1986; pp. 75–82. [Google Scholar]
- Jensen, P.K. Weed size hierarchies in Denmark. Weed Res. 1991, 31, 1–7. [Google Scholar] [CrossRef]
- Berti, A.; Zanin, G. Density Equivalent: A method for forecasting yield loss caused by mixed weed populations. Weed Res. 1994, 34, 327–332. [Google Scholar] [CrossRef]
- Guglielmini, A.C.; Verdu, A.M.C.; Satorre, E.H. Competitive ability of five common weed species in competition with soybean. Int. J. Pest Manag. 2017, 63, 30–36. [Google Scholar] [CrossRef]
- Hernandez Plaza, E.; Navarrete, L.; Gonzalez-Andujar, J.L. Intensity of soil disturbance shapes functional diversity of weed communities: The long-term effect of different tillage system. Agric. Ecosyst. Environ. 2015, 207, 101–108. [Google Scholar] [CrossRef]
- Royama, T. Animal Population Ecology: An Analytical Approach; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Ferrero, R.; Lima, M.; Davis, A.S.; Gonzalez-Andujar, J.L. Weed diversity affects soybean and maize yield in a long term experiment in Michigan, USA. Front. Plant Sci. 2017, 8, 236. [Google Scholar] [CrossRef]
- Ferrero, R.; Lima, M.; Gonzalez-Andujar, J.L. Crop production structure and stability under climate change in South America. Ann. Appl. Biol. 2018, 172, 65–73. [Google Scholar] [CrossRef]
- Gonzalez-Andujar, J.L.; Aguilera, M.J.; Davis, A.S.; Navarrete, L. Disentangling weed diversity and weather impact on long-term crop productivity in a wheat-legume rotation. Field Crops Res. 2019, 232, 24–29. [Google Scholar] [CrossRef]
- Lima, M.; Navarrete, L.; Gonzalez-Andujar, J.L. Climate effects and feedback structure determining weed population dynamics in a long-term experiment. PLoS ONE 2012, 7, e30569. [Google Scholar] [CrossRef]
- Schabenberger, O.; Pierce, F.J. Contemporary Statistical Models for the Plant and Soil Sciences; Taylor & Francis, CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Freckleton, R.P.; Watkinson, A.R. Nonmanipulative of plant community dynamics. Trends Ecol. Evol. 2001, 16, 301–307. [Google Scholar] [CrossRef]
- Storkey, J.; Neve, P. What good is weed diversity? Weed Res. 2018, 58, 239–243. [Google Scholar] [CrossRef]
- Freckleton, R.P.; Watkinson, A.R. Asymmetric competition between plant species. Funct. Ecol. 2001, 15, 615–623. [Google Scholar] [CrossRef]
- Adler, P.B.; Smull, D.; Beard, K.H.; Choi, R.T.; Furniss, T.; Kulmatiski, A.; Meiners, J.M.; Tredennick, A.T.; Veblen, K.E. Competition and coexistence in plant communities: Intraspecific competition is stronger than interspecific competition. Ecol. Lett. 2018, 21, 1319–1329. [Google Scholar] [CrossRef]
- Kemper, R.; Rinke, N.; Gerhards, R.; Böhm, H. Weed suppression and crop yield performance in sole and intercrops of common vetch and spring wheat depending on seed density ratio in organic farming. J. Kult. 2020, 72, 12–24. [Google Scholar]
- Van Delden, A.; Lotz, L.; Bastiaans, L.; Franke, A.C.; Smid, H.G.; Groeneveld, R.M.W.; Kropff, M.J. The influence of nitrogen supply on the ability of wheat and potato to suppress Stellaria media growth and reproduction. Weed Res. 2002, 42, 429–445. [Google Scholar] [CrossRef]
- Liebman, M.; Davis, A.S. Integration of soil, crop and weed management in low-external-input farming systems. Weed Res. 2000, 40, 27–47. [Google Scholar] [CrossRef]
- Wilson, B.J.; Wright, K.J. Predicting the growth and competitive effects of annual weeds in wheat. Weed Res. 1990, 30, 201–221. [Google Scholar] [CrossRef]
- Marshall, E.J.P.; Brown, V.K.; Boatman, N.D.; Lutman, P.J.W.; Squire, G.R.; Ward, L.K. The role of weeds in supporting biological diversity within crop fields. Weed Res. 2003, 43, 77–89. [Google Scholar] [CrossRef]
- Blackshaw, R.E.; Brandt, R.N. Nitrogen fertilizer rate effects on weed competitiveness is species dependent. Weed Sci. 2008, 56, 743–747. [Google Scholar] [CrossRef]
- Berquer, A.; Bretagnolle, V.; Martin, O.; Gaba, S. Disentangling the effect of nitrogen input and weed control on crop-weed competition suggests a potential agronomic trap in conventional farming. Agric. Ecosyst. Environ. 2023, 345, 108–232. [Google Scholar] [CrossRef]
- Song, J.S.; Im, J.H.; Kim, J.W.; Kim, D.G.; Lim, Y.; Yook, M.J.; Lim, S.H.; Kim, D.S. Modeling the Effects of Nitrogen Fertilizer and Multiple Weed Interference on Soybean Yield. Agronomy 2021, 11, 515. [Google Scholar] [CrossRef]
- Di Tomaso, J.M. Approaches for improving crop competitiveness through the manipulation of fertilization strategies. Weed Sci. 1995, 43, 491–497. [Google Scholar] [CrossRef]
- Angonin, C.; Caussanel, J.P.; Meynard, J.M. Competition between winter wheat and Veronica hederifolia: Influence of weed density and the amount and timing of nitrogen application. Weed Res. 1996, 36, 175–187. [Google Scholar] [CrossRef]
- Baylis, J.M.; Watkinson, A.R.; Lintell Smith, G.; Firbank, L. Reduced nitrogen inputs in wheat: The effects on weed-crop competition and weed population dynamics. In Proceedings of the 1st International Weed Control Congress, Melbourne, Australia, 17–21 February 1992; Volume 2, pp. 76–80. [Google Scholar]
- Lehoczky, E.; Kismanyoky, A. Study on the weediness of winter wheat in a long-term fertilization field experiment. Comm. Appl. Biol. Sci. 2006, 71, 793–796. [Google Scholar]
- Kneževic, M.; Stipesevic, B.; Kneževic, I.; Loncaric, Z. Weed populations of winter wheat as affected by tillage and nitrogen. Ekologia 2007, 26, 190–200. [Google Scholar]
- Blackshaw, R.E.; Molnar, L.J.; Larney, F.J. Fertilizer, manure and compost effects on weed growth and competition with winter wheat in western Canada. Crop Prot. 2005, 24, 971–980. [Google Scholar] [CrossRef]
- Mokhtassi-Bidgoli, A.; AghaAlikhani, M.; Nassiri-Mahallati, M.; Zand, E.; Gonzalez-Andujar, J.L.; Azari, A. Agronomic performance, seed quality and nitrogen uptake of Descurainia sophia in response to different nitrogen rates and water regimes. Ind. Crops Prod. 2013, 44, 583–592. [Google Scholar] [CrossRef]
- Holderbaum, J.F.; Decker, A.M.; Meisinger, J.J.; Mulford, F.R.; Vough, L.R. Fall-Seeded Legume Cover Crops for No-Tillage Corn in the Humid East. Agron. J. 1990, 82, 117–124. [Google Scholar] [CrossRef]
- Chantre, G.R.; Gonzalez-Andujar, J.L. Decision Support Systems for Weed Management; Springer International Publishing: Cham, Switzerland, 2020. [Google Scholar]
- Swanton, C. Weed science and the clock of the long now. Weed Sci. 2022, 70, 369. [Google Scholar] [CrossRef]


{kind=link}
| CEREAL | LEGUME | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pseudo-R2 | rmax | c | aDES | aFUM | aPAP | aVER | Pseudo-R2 | rmax | c | aDES | aFUM | aPAP | aVER | ||
| DES | 0.61 | 29.50 | 0.37 | 0.49 | −0.21 | −0.05 | 0.06 | DES | 0.63 | 6.71 | −0.78 | 0.72 | −0.36 | 0.13 | −0.03 |
| FUM | 0.57 | 25.00 | 0.73 | 0.20 | 0.06 | −0.16 | 0.19 | FUM | 0.89 | 5.00 | −0.86 | −0.09 | 0.51 | 0.36 | 0.09 |
| PAP | 0.55 | 15.00 | 0.31 | 0.30 | −0.18 | 0.18 | −0.07 | PAP | 0.72 | 12.90 | 0.18 | 0.00 | 0.01 | 0.48 | −0.27 |
| VER | 0.61 | 10.60 | 0.97 | 0.20 | −0.19 | −0.35 | 0.30 | VER | 0.86 | 3.35 | −1.89 | 0.89 | 0.15 | −0.10 | 0.55 |
| CEREAL | Average | LEGUME | Average | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Species | DES | FUM | PAP | VER | DES | FUM | PAP | VER | ||
| DES | 1 | 0.43 | 0.10 | 0.12 | 0.21 | 1 | 0.51 | 0.18 | 0.05 | 0.25 |
| FUM | 3.14 | 1 | 2.63 | 3.11 | 2.96 | 0.18 | 1 | 0.71 | 0.17 | 0.35 |
| PAP | 1.64 | 1.02 | 1 | 0.40 | 1.02 | 0.01 | 0.03 | 1 | 0.56 | 0.20 |
| VER | 0.67 | 0.64 | 1.16 | 1 | 0.82 | 1.64 | 0.28 | 0.18 | 1 | 0.70 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzalez-Andujar, J.L.; Aguilera, M.J.; Van Acker, R. Quantifying and Disentangling the Competition Effect of a Weed Community in a Long-Term Biennial Cereal-Legume Rotation. Agronomy 2023, 13, 1432. https://doi.org/10.3390/agronomy13051432
Gonzalez-Andujar JL, Aguilera MJ, Van Acker R. Quantifying and Disentangling the Competition Effect of a Weed Community in a Long-Term Biennial Cereal-Legume Rotation. Agronomy. 2023; 13(5):1432. https://doi.org/10.3390/agronomy13051432
Chicago/Turabian StyleGonzalez-Andujar, Jose L., Maria J. Aguilera, and Rene Van Acker. 2023. "Quantifying and Disentangling the Competition Effect of a Weed Community in a Long-Term Biennial Cereal-Legume Rotation" Agronomy 13, no. 5: 1432. https://doi.org/10.3390/agronomy13051432