Spider Plant (Cleome gynandra L.): An Emerging Weed in the Sweet Corn–Brassica Cropping System


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Collection, Multiplication, and Preparation
2.2. Seed Biology Study
General Seed Germination Protocol
2.3. Seed Germination Studies
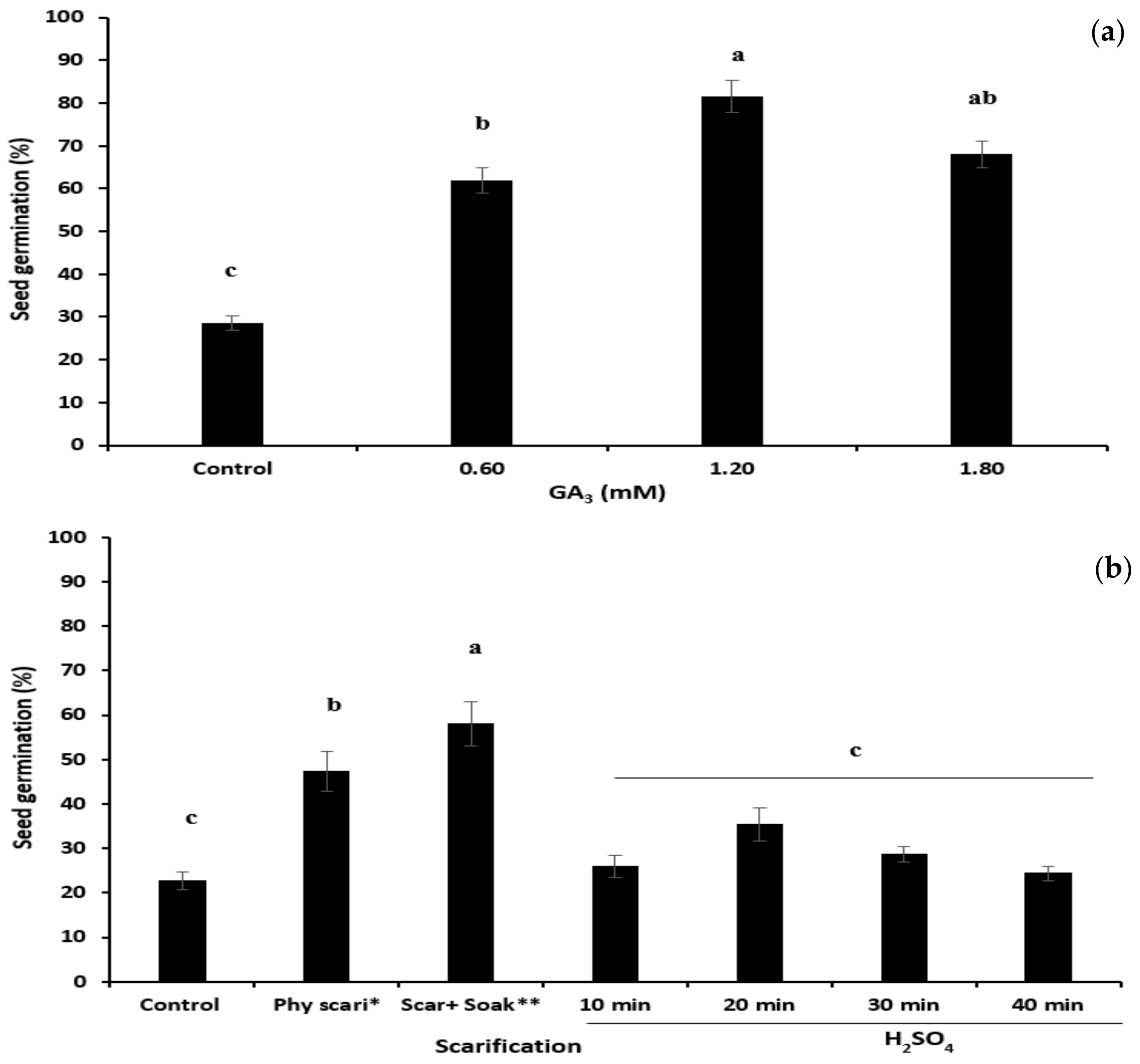
2.3.1. Determination of Appropriate Dormancy-Breaking Treatments
Leaching
Gibberellic Acid
Scarification
Dry After-Ripening
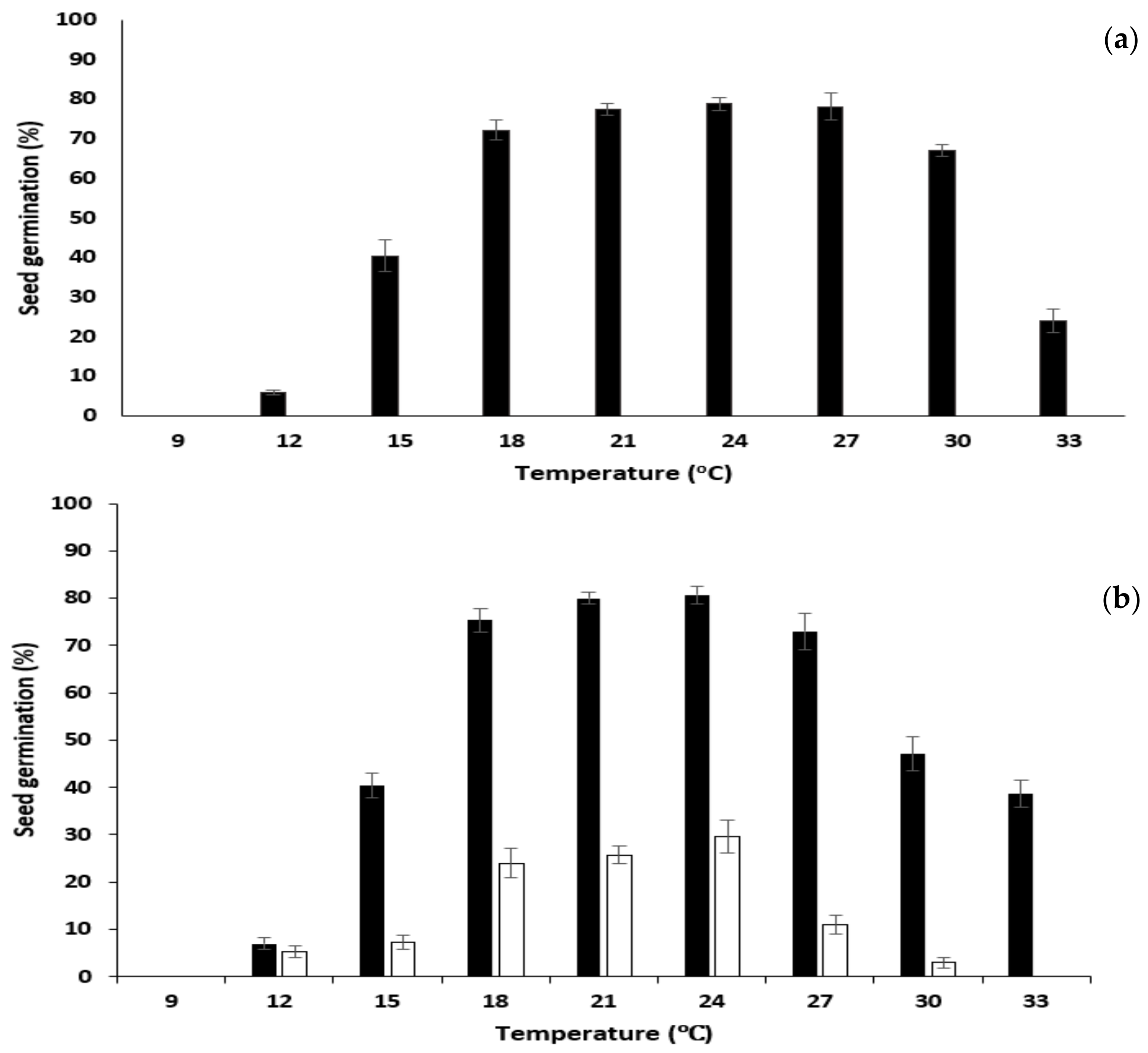
2.3.2. Optimal Germination Conditions
Constant Temperature
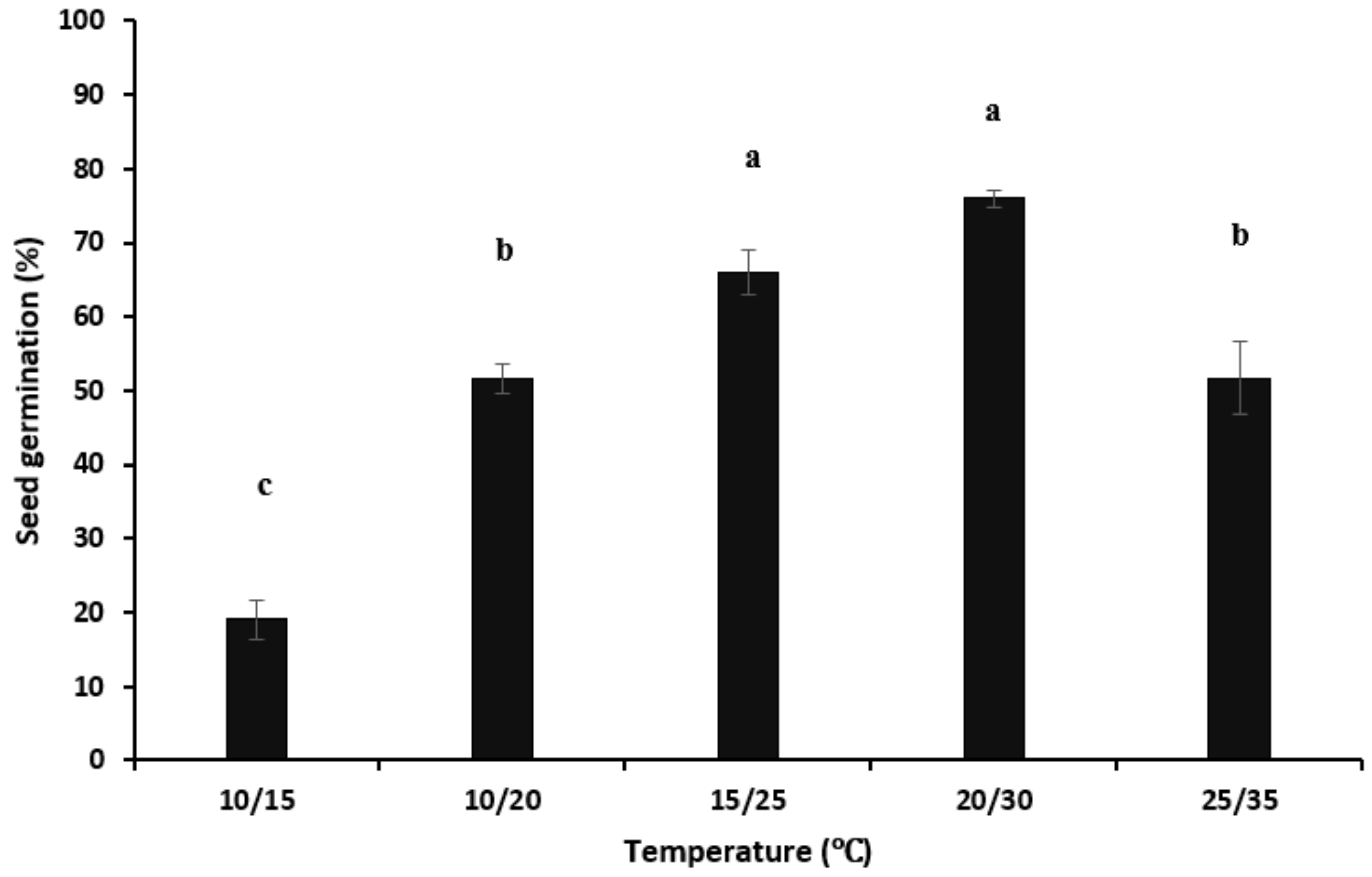
Alternating Temperatures
2.3.3. Seed Germination under Abiotic Stress
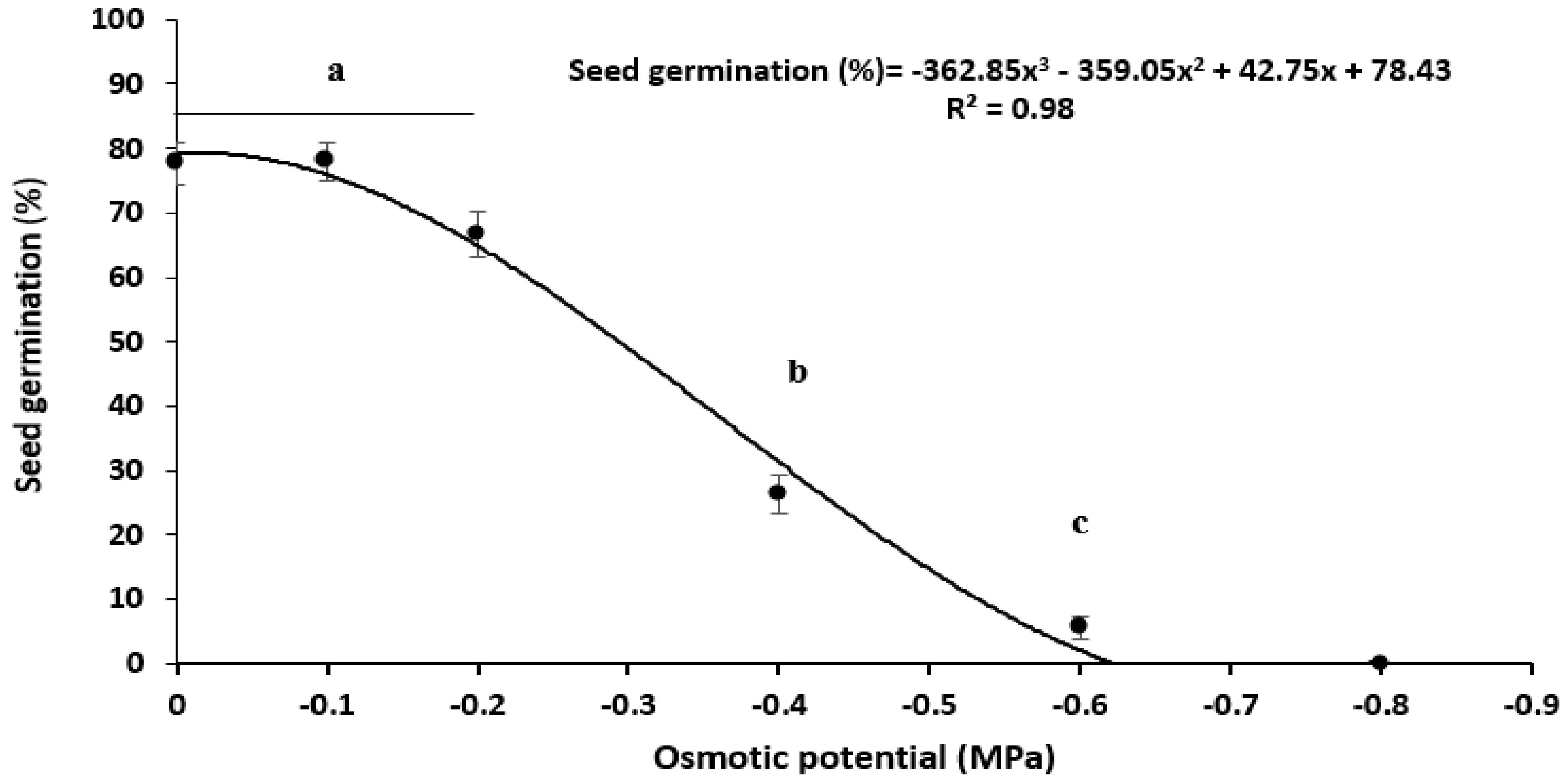
Moisture Stress
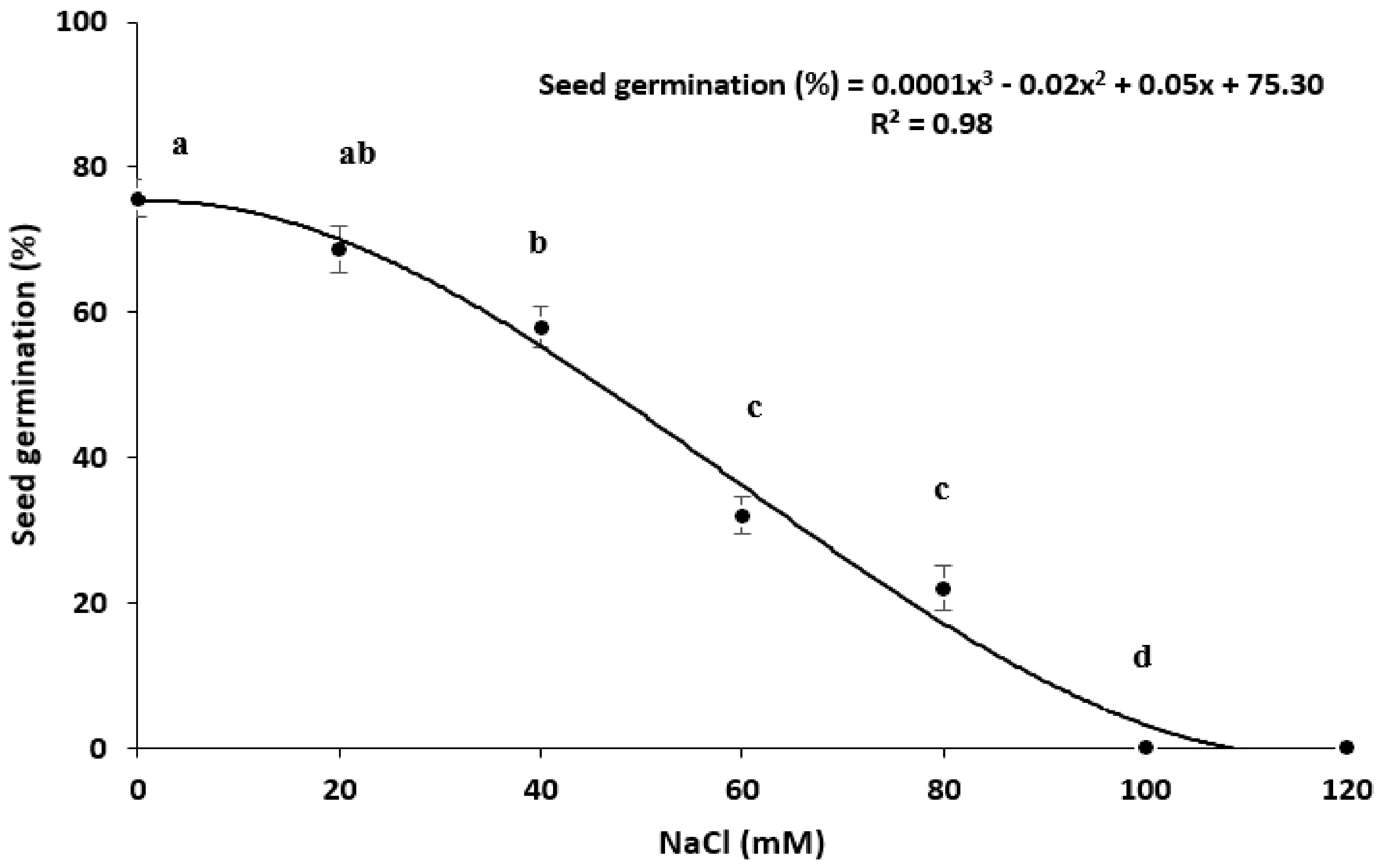
Salinity Stress
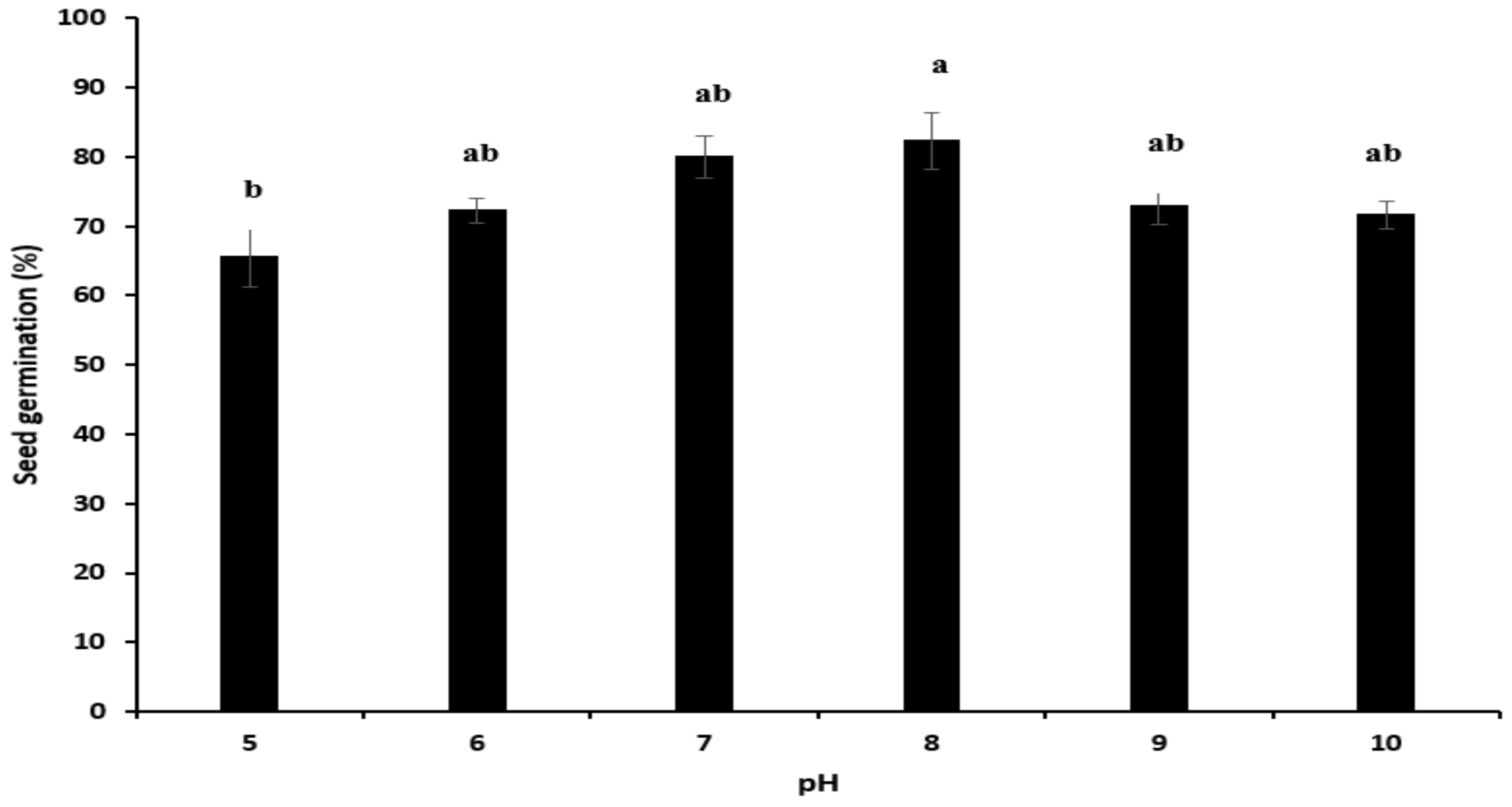
pH Stress
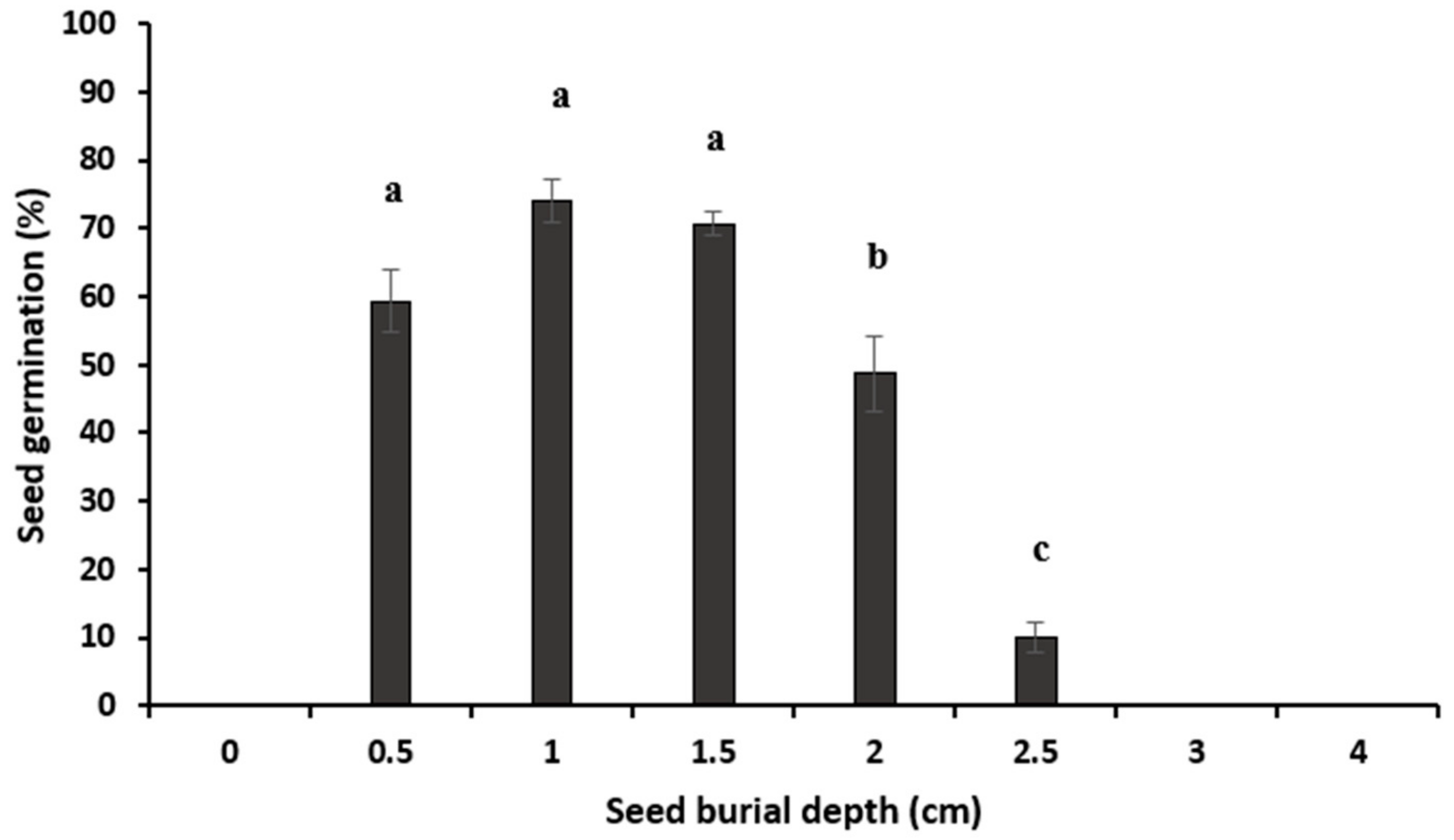
2.3.4. Seed Germination under Variable Burial Depth
Burial Depth
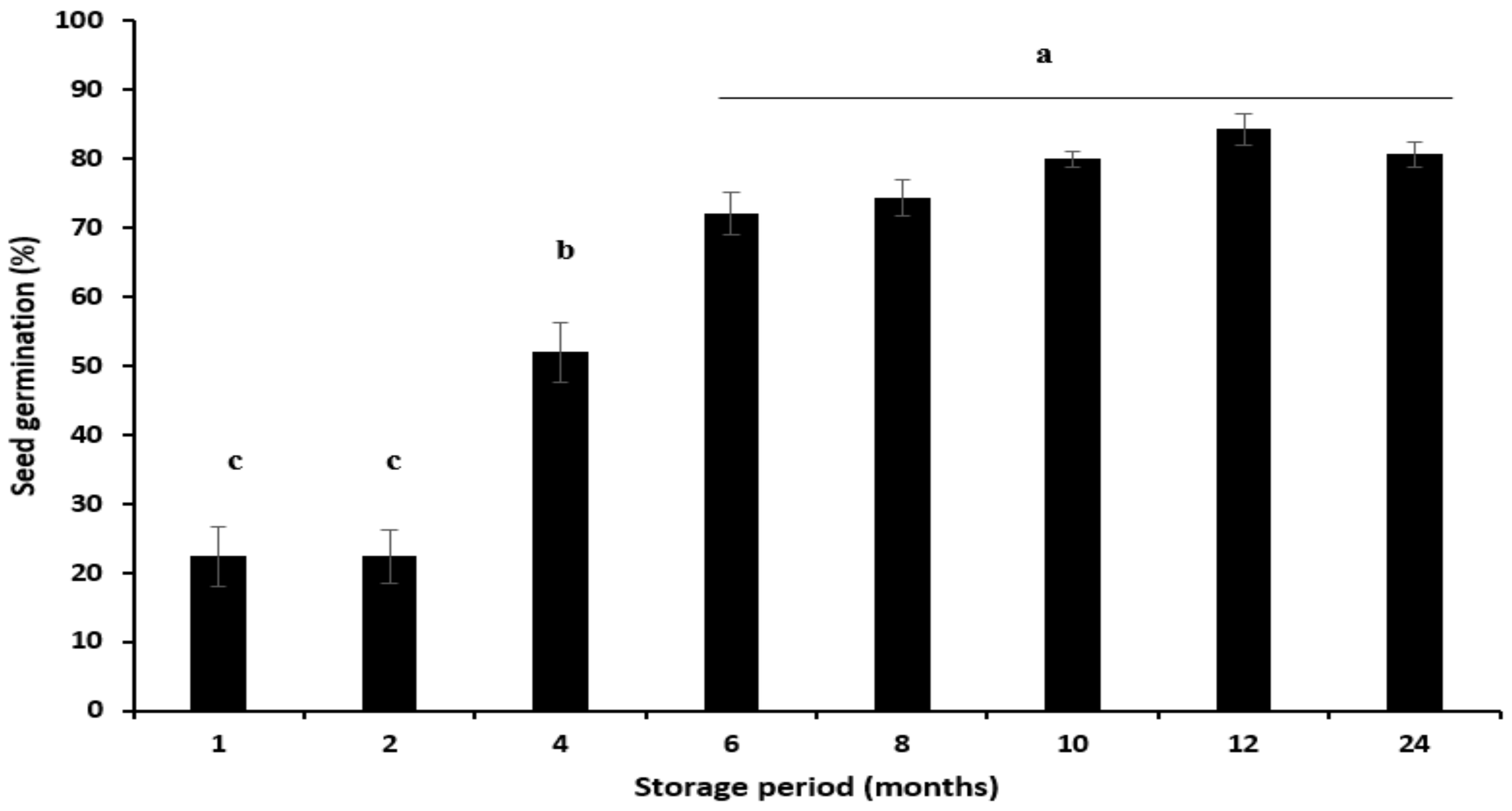
2.3.5. Seed Germination After-Ripening
After-Ripening
2.4. Experimental Design and Statistical Analysis
3. Results
3.1. Determination of Appropriate Dormancy-Breaking Treatments
3.2. Optimal Germination Conditions
3.3. Seed Germination under Abiotic Stress
3.3.1. pH Stress
3.3.2. Moisture Stress
3.3.3. Salinity Stress
3.4. Seed Germination under Variable Burial Depth
3.5. Seed Germination After-Ripening
After-Ripening
4. Discussion
4.1. Determination of an Appropriate Dormancy-Breaking Treatment
4.2. Optimal Germination Conditions
4.3. Seed Germination under Abiotic Stress
4.4. Seed Germination under Variable Burial Depth
4.5. Seed Germination After-Ripening
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kokwaro, J.O.; Riley, B.W.; Brokensha, D. Flowering Plant Families of East Africa: An Introduction to Plant Taxonomy; East African Educational Publishers: Nairobi, Kenya, 1996; Volume 66, p. 321. [Google Scholar]
- Kuhn, U. Dicot Weed: Capparaceae; Hafliger, T.J., Wolf, M., Eds.; CIBA Geigy LTD: Basel, Switzerland, 1988. [Google Scholar]
- Bremer, K.; Wanntorp, H.E. Phylogenetic systematics in botany. Taxon 1978, 27, 317–329. [Google Scholar] [CrossRef]
- Cweya, J.A.; Heller, J.; Engels, J. Cat’s Whiskers, Cleome gynandra L.; International Plant Genetic Resource Institute: Rome, Italy, 1997. [Google Scholar]
- Holm, L.R.G.; Plucknett, D.L.; Pancho, J.V.; Herberger, J.P. A Geographical Atlas of World Weeds, Reprint ed.; John Wiley & Sons: New York, NY, USA, 1991. [Google Scholar]
- Chweya, J.A.; Mnzava, N.A. Promoting the Conservation and Use of Underutilized and Neglected Crops. 11. Cat’s Whiskers (Cleome gynandra L.); International Plant Genetic Resources Institute: Rome, Italy, 1997; pp. 1–54. [Google Scholar]
- Brown, G.K. Introduction to the Census of the Queensland Flora 2021; Queensland Department of Environment and Science: Brisbane, Australia, 2021; pp. 1–29.
- Turner, G.; McMahon, G. Weeds in Australian Cane Fields; Bureau of Sugar Experiment Stations: Indooroopilly, Australia, 1989; pp. 1–84.
- Group, T.S. Regional Food Sector Strategy: Prepared for Lockyer Valley Regional Council; The Stafford Group: Sydney, Australia, 2013; pp. 1–76. [Google Scholar]
- Shilla, O.; Dinssa, F.F.; Omondi, E.O.; Winkelmann, T.; Abukutsa-Onyango, M.O. Cleome gynandra L. origin, taxonomy and morphology: A review. Afr. J. Agric. Res. 2019, 14, 1568–1583. [Google Scholar] [CrossRef]
- Australia, P.H. Domestic Quarantine for the Producers. Available online: https://interstatequarantine.org.au/producers/moving-plant-goods/ (accessed on 17 April 2023).
- Yepes, J.H. Study of weed Cleome gynandra L. Rev. Comalfi 1978, 5, 49–53. [Google Scholar]
- Boonsong, E. Effects of seed maturity, seed storage and pre-germination treatments on seed germination of Cleome (Cleome gynandra L.). Sci. Hortic. 2009, 119, 236–240. [Google Scholar] [CrossRef]
- Ochuodho, J.O.; Modi, A.T.; Beukes, M. The dormancy and germination of Cleome gynandra L. In Proceedings of the Fourth Workshops on Sustainable Horticultural Production in the Tropics, Department of Horticulture, Moi University, Eldoret, Kenya, 24 November 2004. [Google Scholar]
- Bohringer, R.; Lourens, A.; Jansen van Vuuren, P.J. The influence of various constant temperatures on the germination of Cleome gynandra seed. J. South Afr. Soc. Hortic. Sci. 1999, 9, 21–24. [Google Scholar]
- Holm, L.; Doll, J.; Holm, E.; Pancho, J.V.; Herberger, J.P. World Weeds: Natural Histories and Distribution; Wiley: New York, NY, USA, 1997. [Google Scholar]
- Navie, S.C. The Biology of Parthenium hysterophorus L. in Australia. Ph.D. Thesis, The University of Queensland, Brisbane, Australia, 2002. [Google Scholar]
- Tamado, T.; Schutz, W.; Milberg, P. Germination ecology of the weed Parthenium hysterophorus in eastern Ethiopia. Ann. Appl. Biol. 2002, 140, 263–270. [Google Scholar] [CrossRef]
- Peters, J. Tetrazolium Testing Handbook: Contribution No. 29 to the Handbook on Seed Testing; AOSA: Lincoln, NE, USA, 2000. [Google Scholar]
- Elias, S.G. Seed Testing Principles and Practices; Michigan State University Press: East Lansing, MI, USA, 2012. [Google Scholar]
- Michel, B.E. Evaluation of the water potentials of solutions of polyethylene glycol 8000 both in the absence and presence of other solutes. Plant Physiol. 1983, 72, 66–70. [Google Scholar] [CrossRef]
- Rengasamy, P. World salinization with emphasis on Australia. J. Exp. Bot. 2006, 57, 1017–1023. [Google Scholar] [CrossRef]
- De Caritat, P.; Cooper, M.; Wilford, J. The pH of Australian soils: Field results from a national survey. Soil Res. 2011, 49, 173–182. [Google Scholar] [CrossRef]
- Chachalis, D.; Reddy, K.N. Factors affecting Campsis radicans seed germination and seedling emergence. Weed Sci. 2000, 48, 212–216. [Google Scholar] [CrossRef]
- Bewley, J.D.; Black, M. Physiology and Biochemistry of Seeds in Relation to Germination; Springer: Berlin/Heidelberg, Germany, 1978. [Google Scholar]
- Bewley, J.D.; Bradford, K.; Hilhorst, H.; Nonogaki, H. Seeds, 3rd ed.; Springer: New York, NY, USA, 2012. [Google Scholar]
- Kelly, K.M.; Staden, J.V.; Bell, W.E. Seed coat structure and dormancy. Plant Growth Regul. 1992, 11, 201–209. [Google Scholar] [CrossRef]
- Taylorson, R.B. Recent Advances in the Development and Germination of Seeds; Plenum Press: New York, NY, USA, 1989. [Google Scholar]
- Mekenian, M.R.; Willemsen, R.W. Germination characteristics of Raphanus raphanistrum. I. Laboratory studies. Bull. Torrey Bot. Club 1975, 102, 243–252. [Google Scholar] [CrossRef]
- Sozzi, G.O.; Chiesa, A. Improvement of caper (Capparis spinosa L.) seed germination by breaking seed coat-induced dormancy. Sci. Hortic. 1995, 62, 255–261. [Google Scholar] [CrossRef]
- Murray, D.R. Seed Physiology; Academic: Sydney, Australia, 1984. [Google Scholar]
- Werker, W. Seed dormancy as explained by the anatomy of embryo envelopes. Isr. J. Bot. 1980, 29, 22–44. [Google Scholar]
- Bachelard, E.P. Effects of Gibberellic acid, Kinetin, and light on the germination of dormant seeds of some Eucalypt species. Aust. J. Bot. 1967, 15, 393–401. [Google Scholar] [CrossRef]
- Bell, D.T.; Rokich, D.P.; McChesney, C.J.; Plummer, J.A. Effects of temperature, light and gibberellic acid on the germination of seeds of 43 species native to Western Australia. J. Veg. Sci. 1995, 6, 797–806. [Google Scholar] [CrossRef]
- Khan, A.A.; Saminy, C. The Physiology and Biochemistry of Seed Development, Dormancy and Germination. In Hormones in Relation to Priming and Secondary Seed Dormancy; Khan, A.A., Ed.; Elsevier Biomedical: Amsterdam, The Netherlands, 1982; pp. 203–241. [Google Scholar]
- Halmer, P.; Bewley, J.D.; Thorpe, T.A. An Enzyme to Degrade Lettuce Endosperm Cell Walls. Appearance of a Mannanase Following Phytochrome- and Gibberellin-induced Germination. Planta 1976, 130, 189–196. [Google Scholar] [CrossRef]
- Jones, R.L.; Stoddard, J.L. Gibberellins and Seed Germination. In The Physiology and Biochemistry of Seed Dormancy and Germination; Khan, A.A., Ed.; North-Holland Publishing: Amsterdam, The Netherlands, 1977; pp. 77–109. [Google Scholar]
- Thomas, T.H. Some reflections on the relationship between endogenous hormones and light-mediated seed dormancy. Plant Growth Regul. 1992, 11, 239–248. [Google Scholar] [CrossRef]
- Ochuodho, J.O.; Modi, A.T. Temperature and light requirements for the germination of Cleome gynandra seeds. S. Afr. J. Plant Soil 2005, 22, 49–54. [Google Scholar] [CrossRef]
- Thanos, C.A.; Georghiou, K.; Douma, D.J.; Marangaki, C.J. Photoinhibition of seed germination in Mediterranean maritime plants. Ann. Bot. 1991, 68, 469–475. [Google Scholar] [CrossRef]
- Thanos, C.A.; Mitrakos, K. Watermelon seed germination. 1. Effects of light, temperature and osmotic stress. Seed Sci. Res. 1992, 2, 155–162. [Google Scholar] [CrossRef]
- Benvenuti, S.; Macchia, M.; Miele, S. Quantitative analysis of emergence of seedlings from buried weed seeds with increasing soil depth. Weed Sci. 2001, 49, 528–535. [Google Scholar] [CrossRef]
- Williams, J.D.; Groves, R.H. The influence of temperature and photoperiod growth and development of “Parthenium lysterophorus” L. Weed Res. 1980, 20, 47. [Google Scholar] [CrossRef]
- Pandey, H.N.; Dubey, S.K. Achene germination of Parthenium hysterophorus L.: Effects of light, temperature, provenance and achene size. Weed Res. 1988, 28, 185–190. [Google Scholar] [CrossRef]
- Piggin, C.M.; Reeves, T.G.; Brooke, H.D.; Code, G.R. Germination of wild radish (Raphanus raphanistrum L.). In Proceedings of the First Conference of the Council of The Australian Weed Science Society, Melbourne, Australia; 1978; pp. 233–240. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: London, UK, 2014. [Google Scholar]
- Chauhan, B.S.; Johnson, D.E. Seed germination ecology of Portulaca oleracea L.: An important weed of rice and upland crops. Ann. Appl. Biol. 2009, 155, 61–69. [Google Scholar] [CrossRef]
- Manalil, S.; Hafiz, H.A.; Chauhan, B.S. Germination ecology of turnip weed (Rapistrum rugosum (L.) All.) in the northern regions of Australia. PLoS ONE 2018, 13, e0201023. [Google Scholar] [CrossRef]
- Pierce, G.L.; Warren, S.L.; Mikkelsen, R.L.; Linker, H.M. Effects of soil calcium and pH on seed germination and subsequent growth of large crabgrass (Digitaria sanguinalis). Weed Technol. 1999, 13, 421–424. [Google Scholar] [CrossRef]
- Teuton, T.C.; Brecke, B.J.; Bryan, U.J.; MacDonald, G.E.; Miller, G.L.; Ducar, J.T. Factors affecting seed germination of tropical signalgrass (Urochloa subquadripara). Weed Sci. 2004, 52, 376–381. [Google Scholar] [CrossRef]
- Ismail, B.S.; Chuah, T.S.; Salmijah, S.; Teng, Y.T.; Schumacher, R.W. Germination and seedling emergence of glyphosate-resistant and susceptible biotypes of goosegrass (Eleusine indica (L.) Gaertn.). Weed Biol. Manag. 2002, 2, 177–185. [Google Scholar] [CrossRef]
- Kulya, J.; Lontom, W.; Bunnag, S.; Theerakulpisut, P. Cleome gynandra L. (C4 plant) shows higher tolerance of salt stress than its C3 close relative, C. viscosa L. Adv. Agric. Bot. 2011, 3, 59–66. [Google Scholar]
- Sabila, M.H.; Grey, T.L.; Webster, T.M.; Vencill, W.K.; Shilling, D.G. Evaluation of factors that influence Benghal dayflower (Commelina benghalensis) seed germination and emergence. Weed Sci. 2012, 60, 75–80. [Google Scholar] [CrossRef]
- Sugimoto, K. Evaluation of salt tolerance in Japanese wild radishes (Raphanus sativus f. raphanistroides Makino). Bull. Minamikyushu Univ. A 2009, 39, 79–88. [Google Scholar]
- Woolley, J.T.; Stoller, E.W. Light penetration and light induced seed germination in soil. Plant Physiol. 1978, 61, 597–600. [Google Scholar] [CrossRef]
- Taylor, G.B.; Revell, C.K. Effect of pod burial, light, and temperature on seed softening in yellow serradella. Aust. J. Agric. Res. 1999, 50, 1203–1209. [Google Scholar] [CrossRef]
- Nutt, B.J. Incidence and Inheritance of Hard-Seededness and Early Maturity in Ornithopus sativus. Ph.D. Thesis, Murdoch University, Perth, Australia, 2012. [Google Scholar]
- Grundy, A.C.; Mead, A.; Burston, S. Modelling the emergence response of weed seeds to burial depth: Interactions with seed density, weight and shape. J. Appl. Ecol. 2003, 40, 757–770. [Google Scholar] [CrossRef]
- Husrev, M.; Bernard, Z.H. The effects of depth and duration of seed burial on viability, dormancy, germination, and emergence of ivyleaf speedwell (Veronica hederifolia). Weed Technol. 2017, 20, 438–444. [Google Scholar] [CrossRef]
- Mohler, C.L. A model of the effects of tillage on emergence of weed seedlings. Ecol. Appl. 1993, 3, 53–73. [Google Scholar] [CrossRef]
- Lapham, J.; Drennan, D.S.H. The fate of yellow nutsedge (Cyperus esculentus) seed and seedlings in soil. Weed Sci. 1990, 38, 125–128. [Google Scholar] [CrossRef]
- Peachey, R.E.; Mallory-Smith, C. Influence of winter seed position and recovery date on hairy nightshade (Solanum sarrachoides) recruitment and seed germination, dormancy, and mortality. Weed Sci. 2007, 55, 49–59. [Google Scholar] [CrossRef]
- Puricelli, E.; Faccini, D.; Orioli, G.; Sabbatini, M.R. Seed survival and predation of Anoda cristata in soyabean crops. Weed Res. 2005, 45, 477–482. [Google Scholar] [CrossRef]
- Robert, G. Secondary Seed Dormancy and the Seedbank Ecology of Brassica napus L. in Western Canada. Ph.D. Thesis, University of Manitoba, Winnipeg, MB, Canada, 2003. [Google Scholar]
- Taylor, I.N.; Walker, S.R.; Adkins, S.W. Burial depth and cultivation influence emergence and persistence of Phalaris paradoxa seed in an Australian sub-tropical environment. Weed Res. 2005, 45, 33–40. [Google Scholar] [CrossRef]
- Geneve, R.L. Seed dormancy in commercial vegetable and flower species. Seed Technol. 1998, 20, 236–250. [Google Scholar]
- Groot, S.P.C.; Karssen, C.M. Dormancy and germination of abscisic acid-deficient tomato seeds. Studies with the sitiens mutant. Plant Physiol. 1992, 99, 952–958. [Google Scholar] [CrossRef] [PubMed]
- Leubner-Metzger, G. Functions and regulation of β-1,3-glucanases during seed germination, dormancy release and after-ripening. Seed Sci. Res. 2003, 13, 17–34. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saifullah, K.; Williams, A.; Adkins, S. Spider Plant (Cleome gynandra L.): An Emerging Weed in the Sweet Corn–Brassica Cropping System. Agronomy 2023, 13, 1430. https://doi.org/10.3390/agronomy13051430
Saifullah K, Williams A, Adkins S. Spider Plant (Cleome gynandra L.): An Emerging Weed in the Sweet Corn–Brassica Cropping System. Agronomy. 2023; 13(5):1430. https://doi.org/10.3390/agronomy13051430
Chicago/Turabian StyleSaifullah, Khaled, Alwyn Williams, and Steve Adkins. 2023. "Spider Plant (Cleome gynandra L.): An Emerging Weed in the Sweet Corn–Brassica Cropping System" Agronomy 13, no. 5: 1430. https://doi.org/10.3390/agronomy13051430