

A Plastid RNA Polymerase-Associated Protein Is Involved in Early Chloroplast Development in Rice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
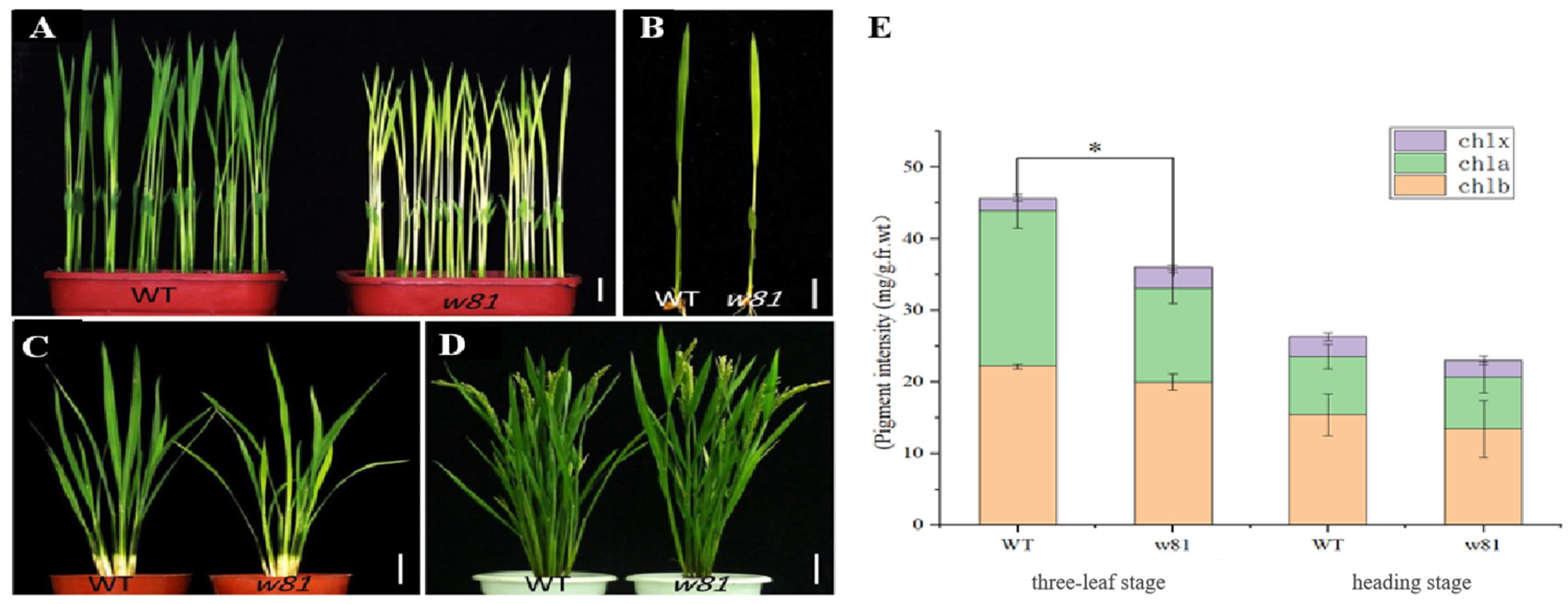
2.2. Measurement of Chlorophyll Contents
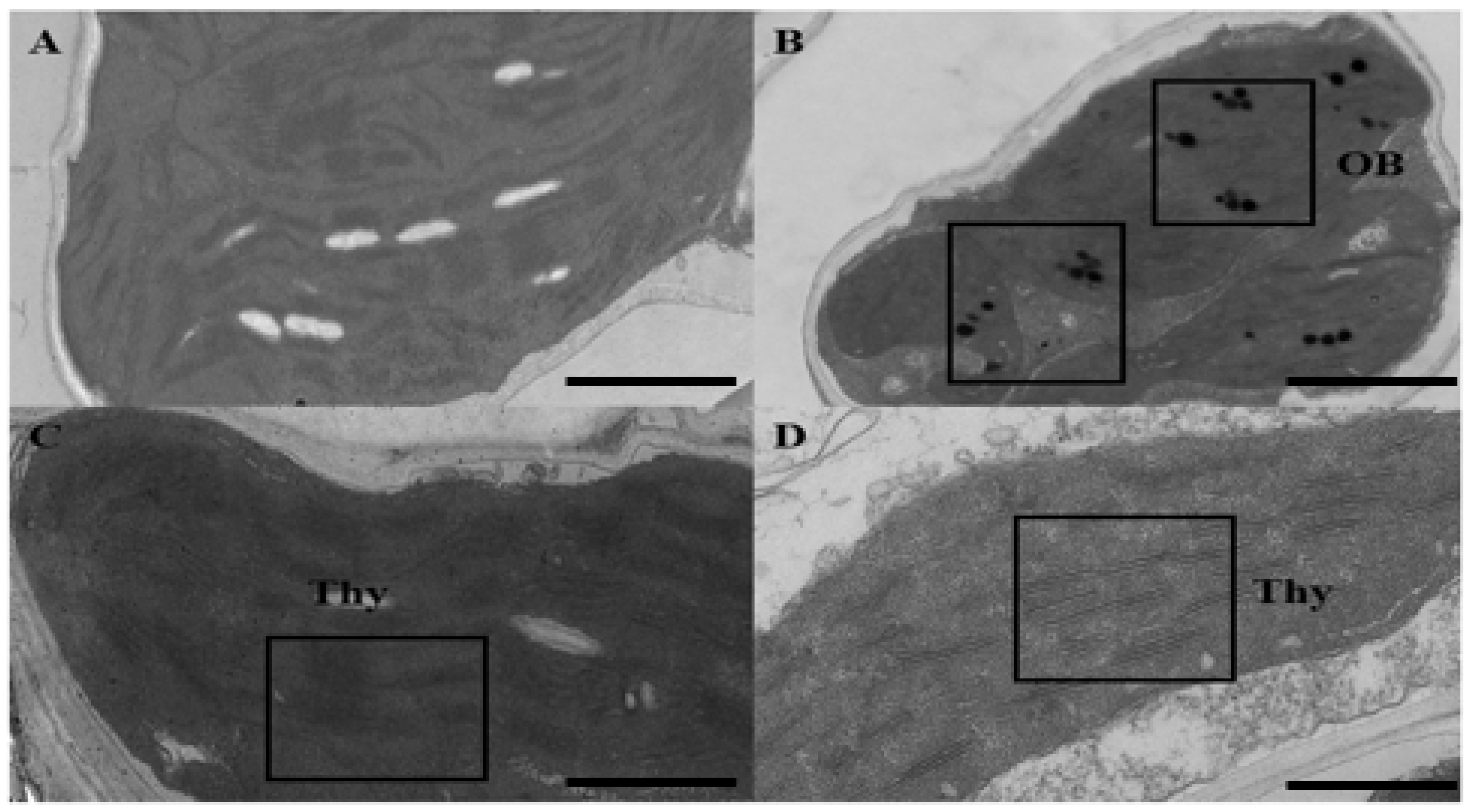
2.3. Ultrastructural Analysis of Chloroplast
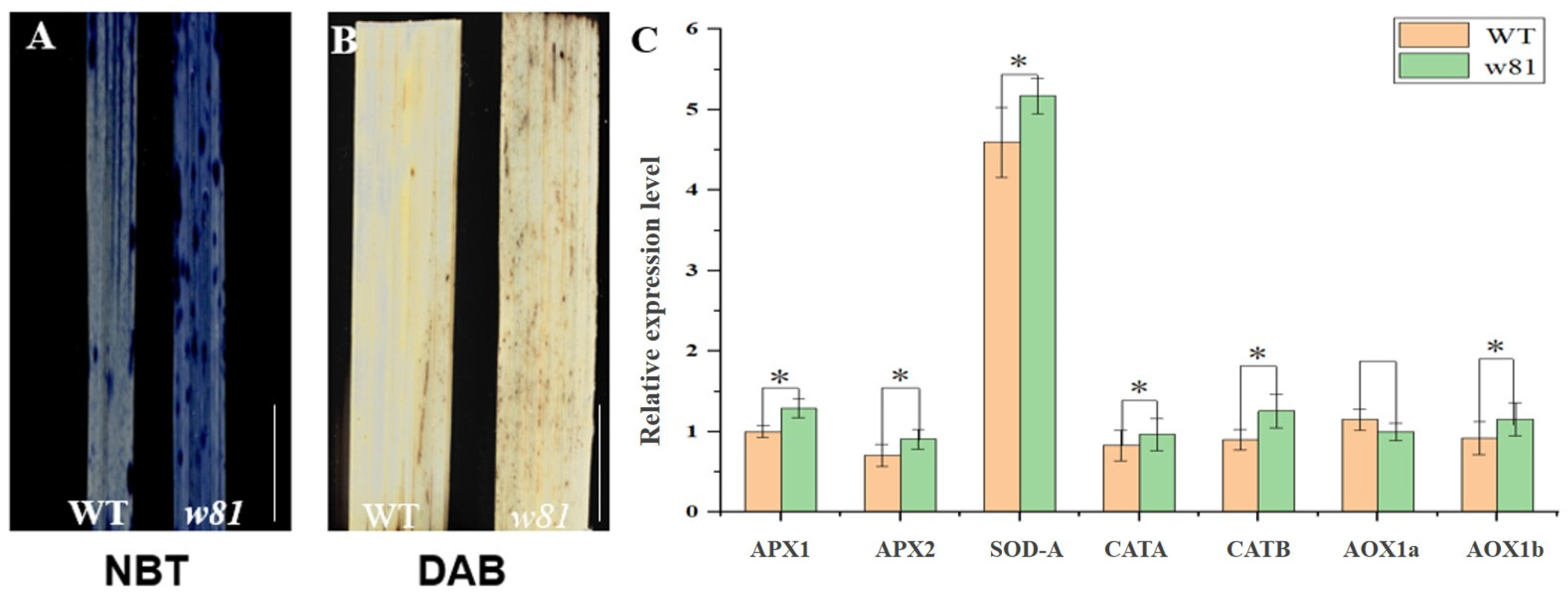
2.4. Reactive Oxygen Species (ROS) Detection
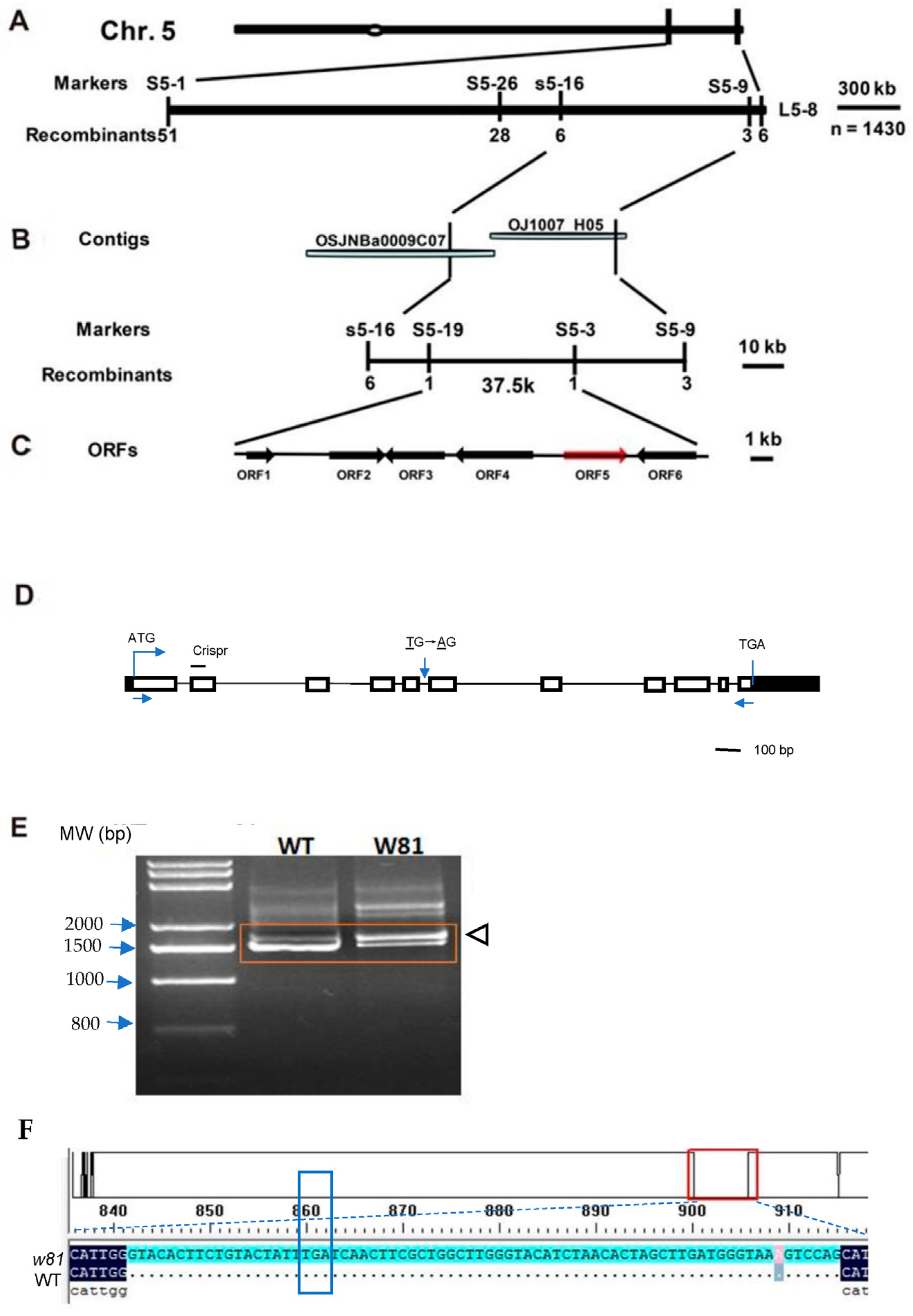
2.5. Cloning of the W81 Gene
2.6. Gene Complementation and Knocking out of W81
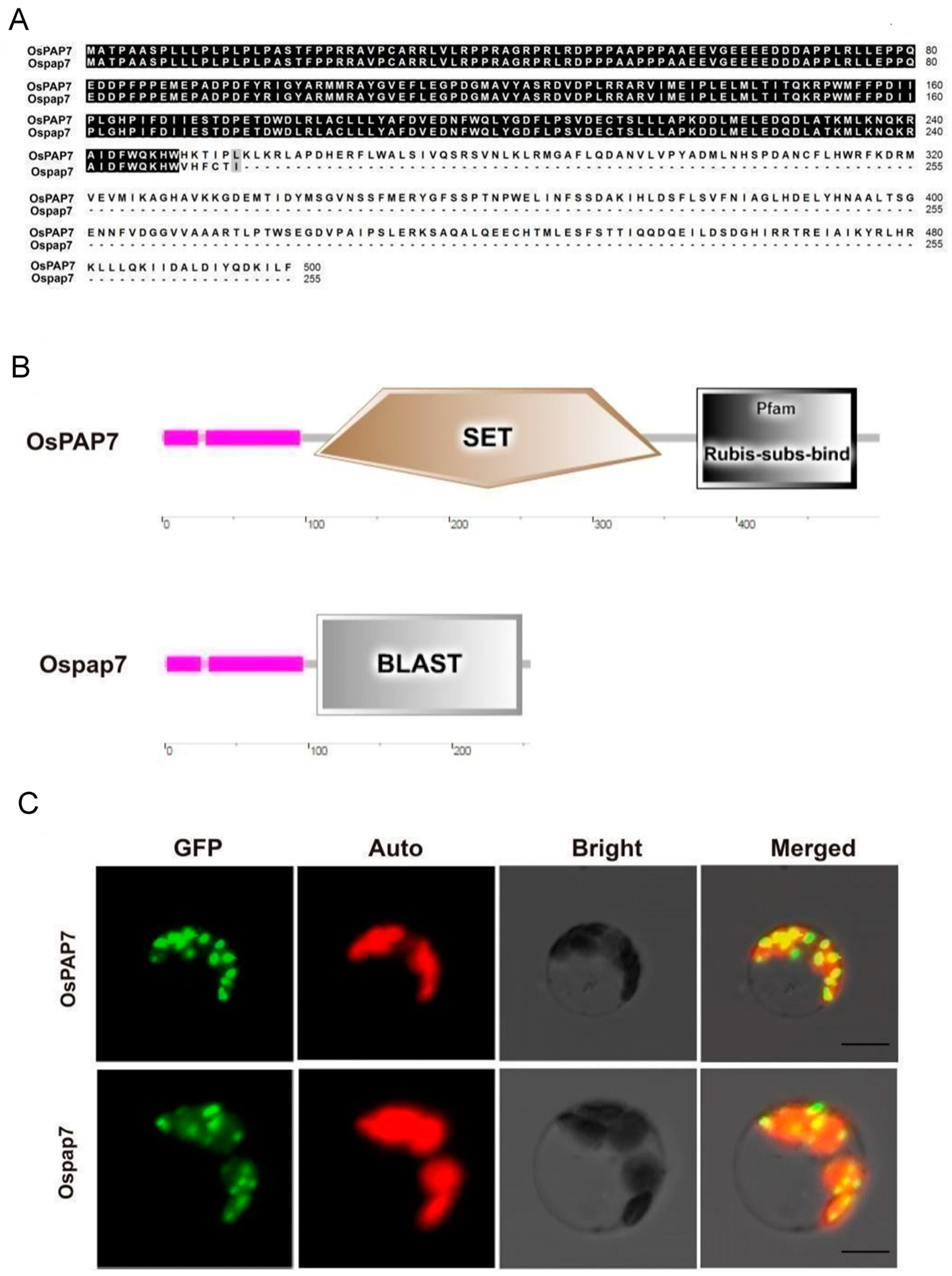
2.7. Subcellular Localization
2.8. Quantitative Real-Time PCR (qRT-PCR) Analysis
2.9. Yeast Two-Hybrid Assay
2.10. Bimolecular Fluorescence Complementation Assay
3. Results
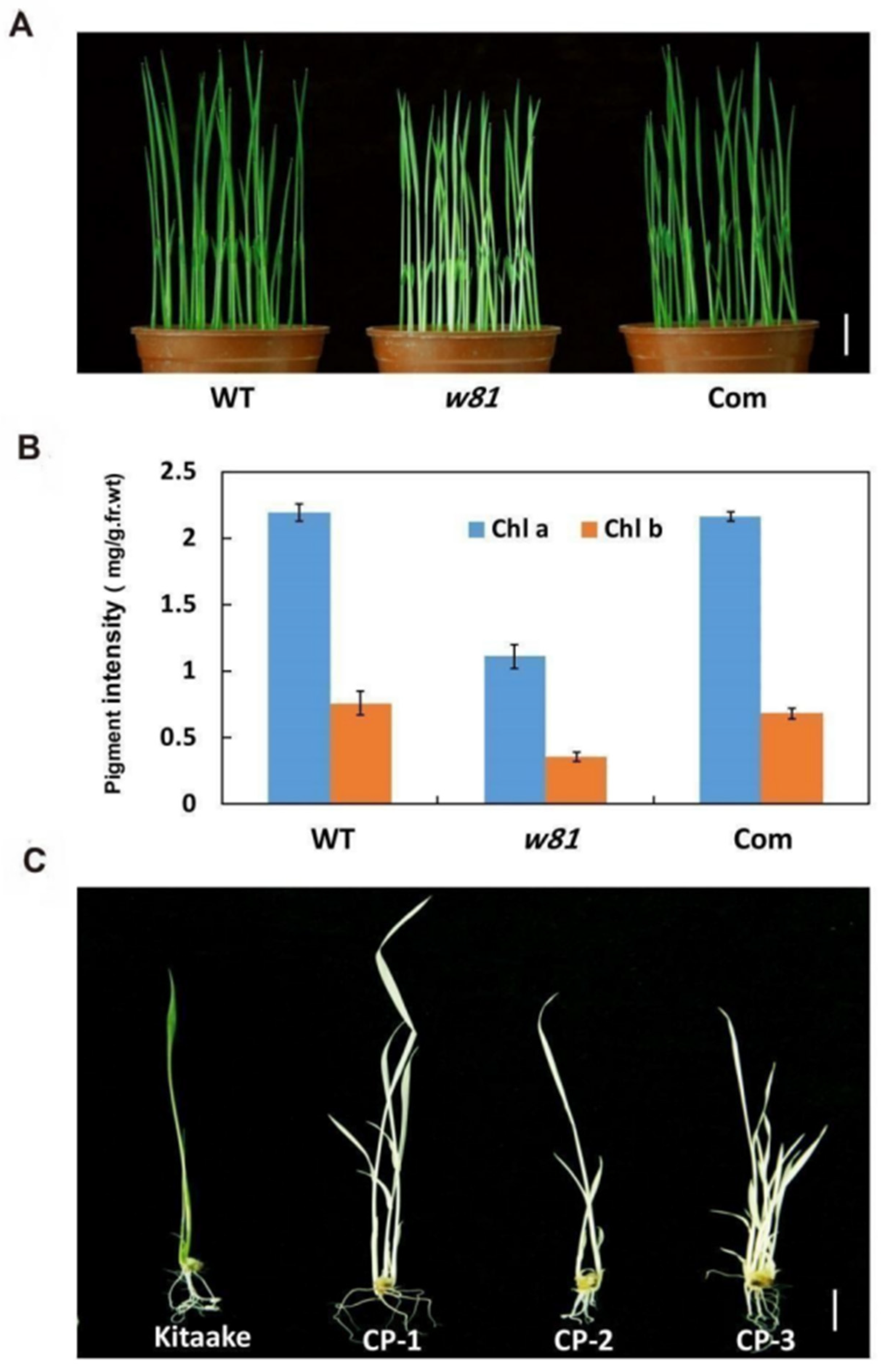
3.1. Phenotypic Analysis of the w81 Mutant
3.2. Analysis of ROS Accumulation in the w81 Mutant
3.3. Map-Based Cloning
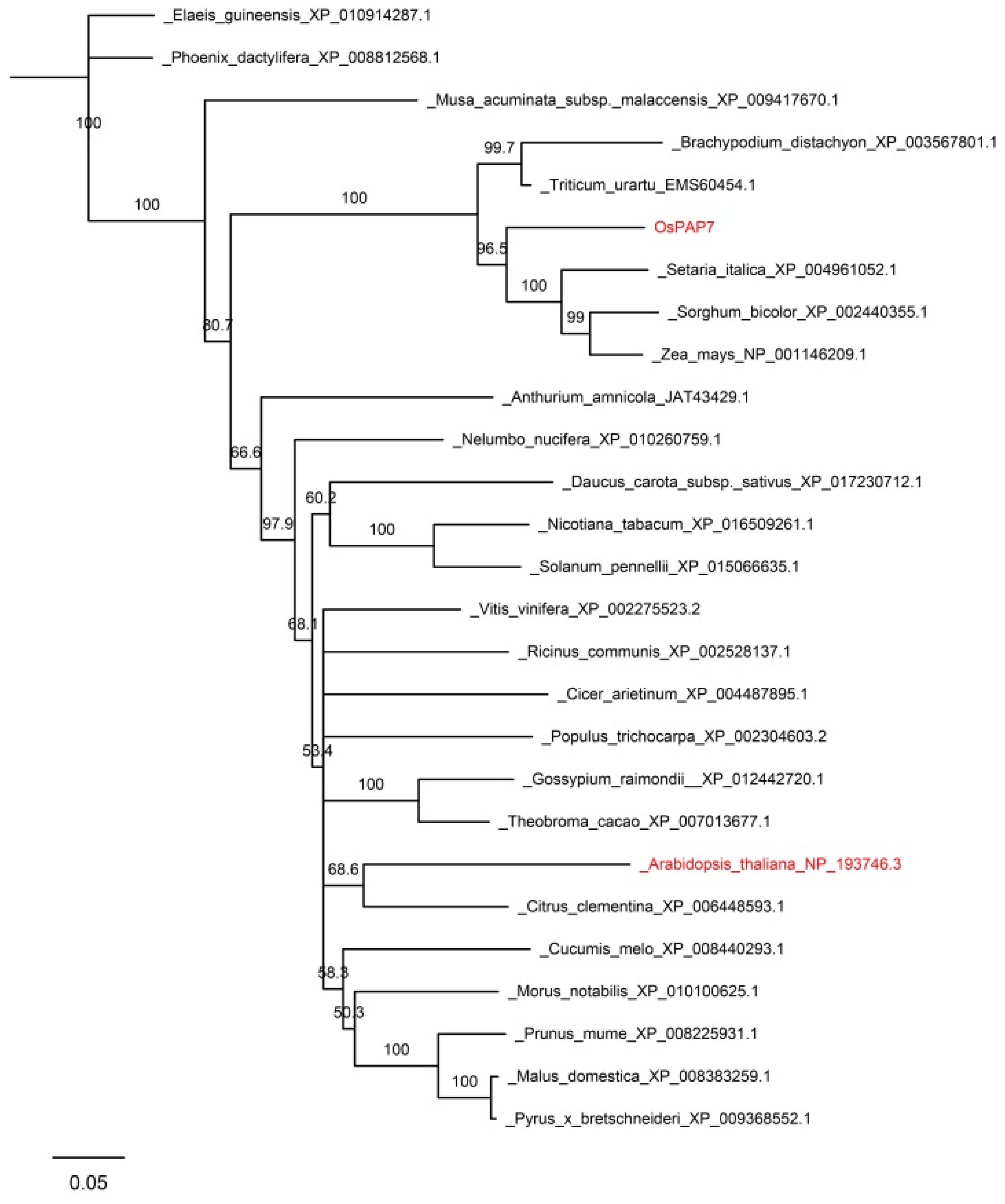
3.4. Bioinformatics Analysis and Subcellular Localization of OsPAP7
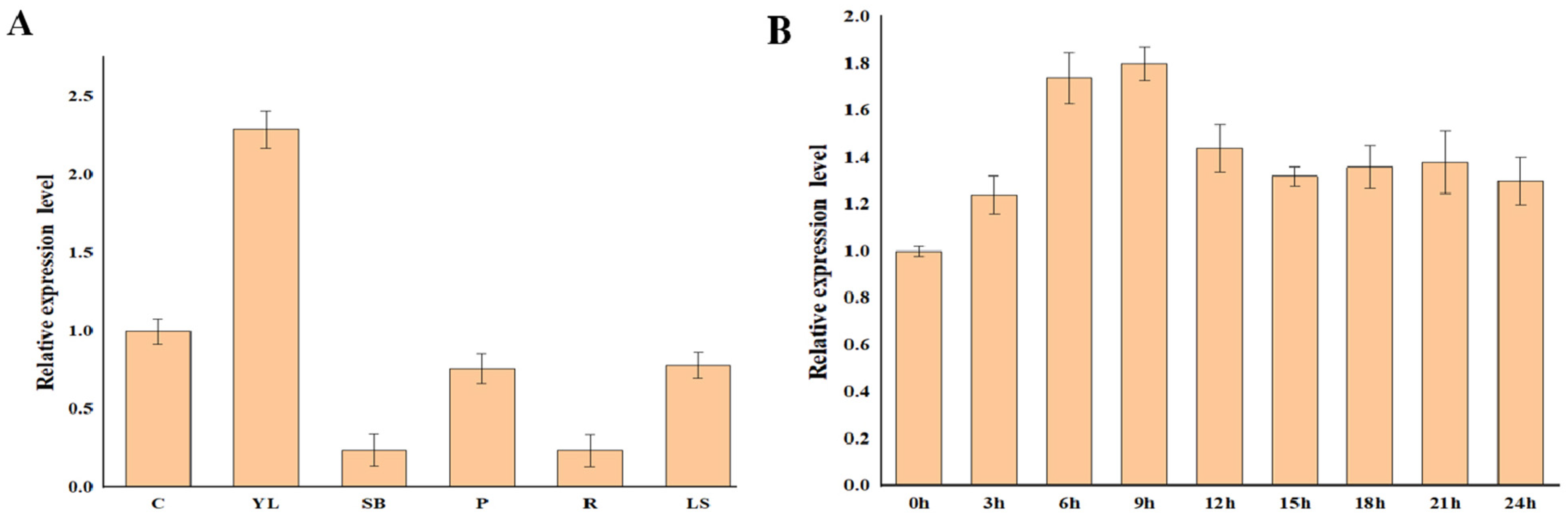
3.5. Expression Analysis of OsPAP7
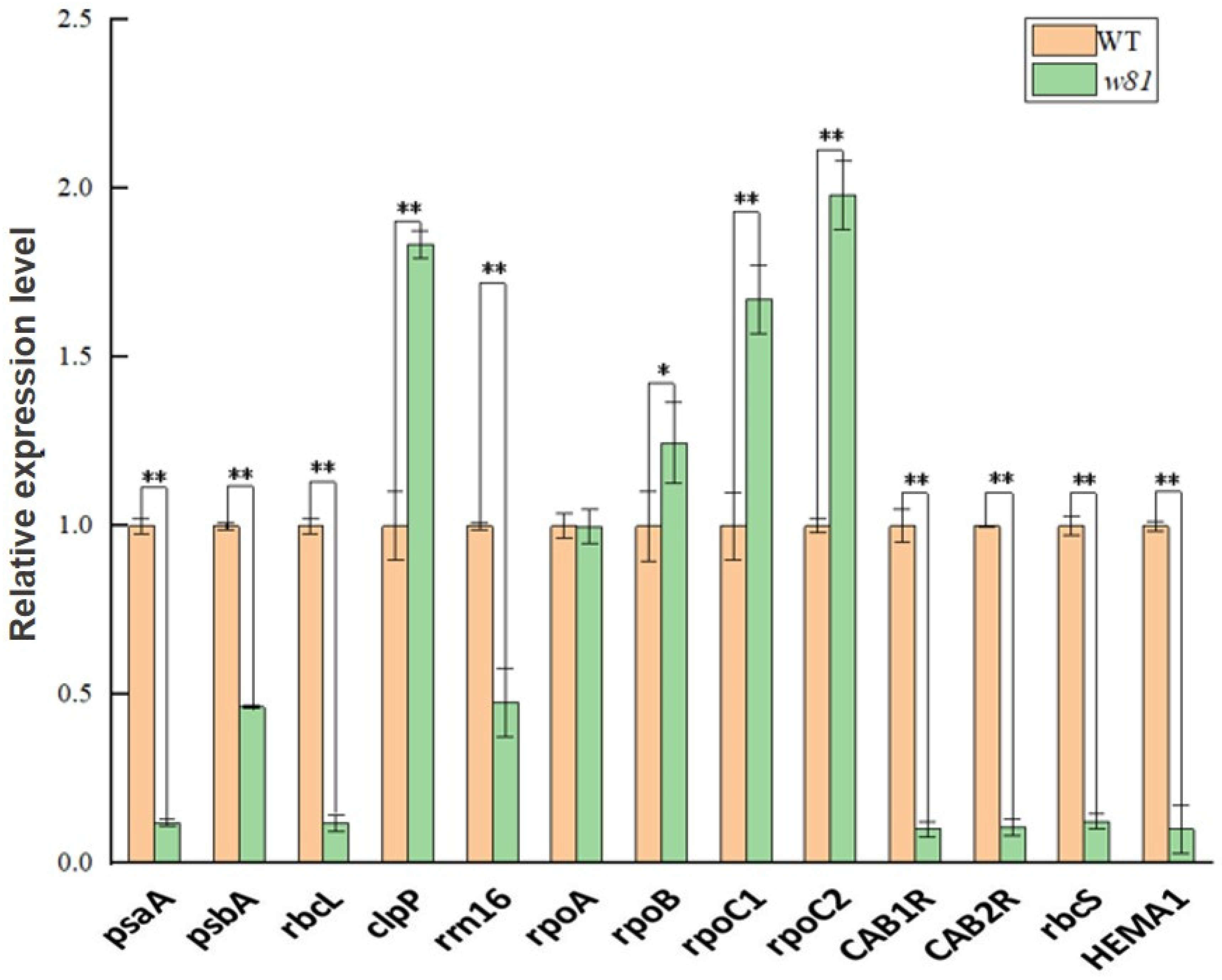
3.6. Expression of Plastid- and Nuclear-Encoded Genes
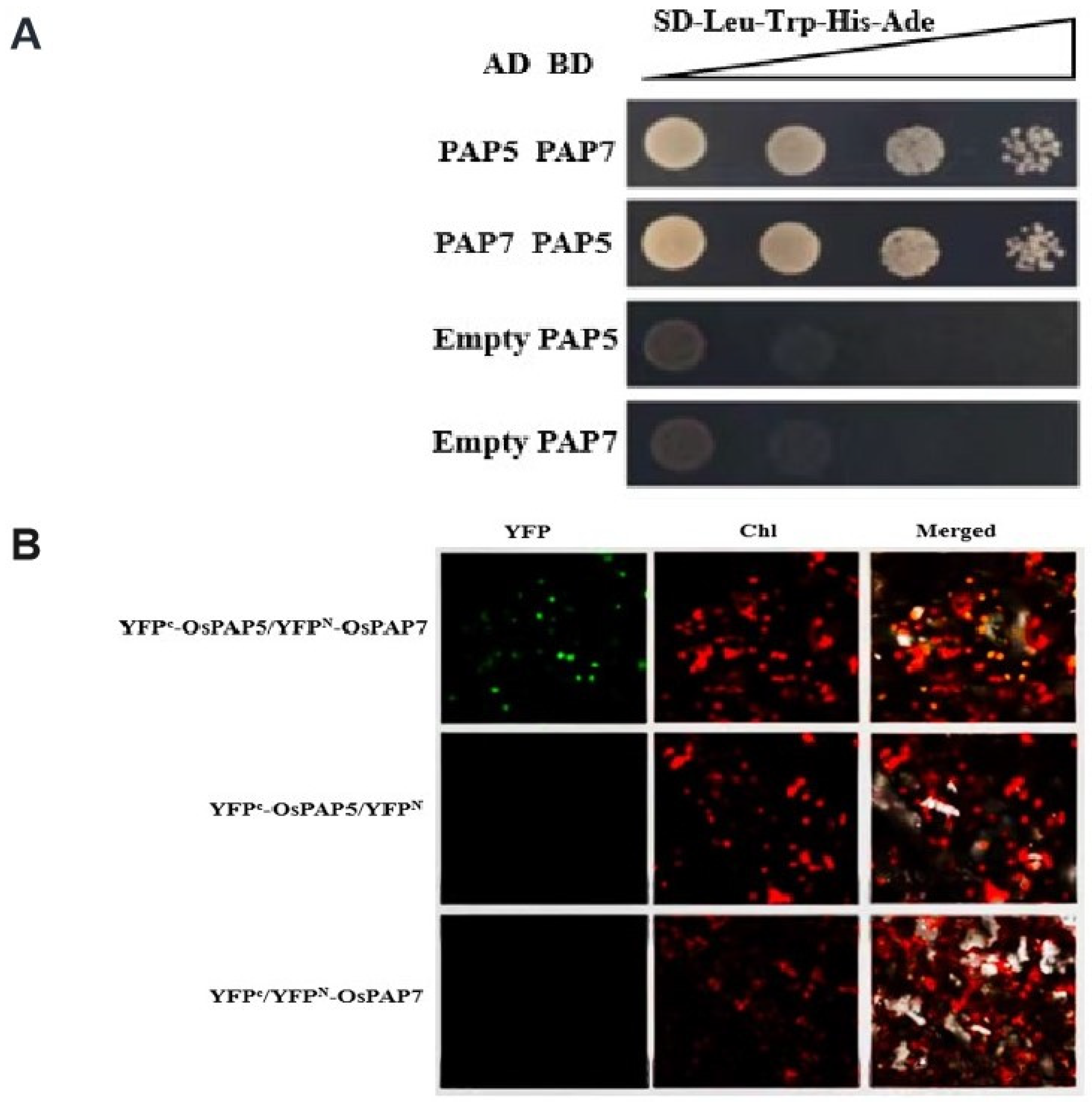
3.7. OsPAP7 Directly Interacts with OsPAP5
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Primer | Sequence (5′–3′) |
|---|---|
| Primers for Map-Based Cloning | |
| S5-1-F | TCACATTGCCTCCAACAAAG |
| S5-1-R | TTCCCAGGTCGTTTGTTTTC |
| S5-26-F | TCAATAGGCCCAACCAAAAG |
| S5-26-R | AGTGACGCGATCTCCAACTC |
| S5-16-F | ACATCGTACGTGTGCATGGT |
| S5-16-R | CGAATTCATCGTCCTCCATC |
| S5-9-F | TCCATTCCACGTACTGTTGC |
| S5-9-R | CACAAGGGGATTGAGGAGAA |
| L5-8-F | TCTCCTTGGCCATCGTATTC |
| L5-8-R | GGCCGCATTTGTATGAAATC |
| S5-19-F | ATCTCCACCTACGCCCTCTT |
| S5-19-R | CTGTGGATCTCCTGCCTCTC |
| S5-3-F | CTTTGGTTTTGGTCGGTTGT |
| S5-3-R | TCTTGCCCCTTGAGACATTT |
| 980-J-3-F | AACTTCGCTGGCTTGGGTAC |
| 980-J-3-R | ATCTCCTGGTCTTGTTGAATAGTG |
| Primers for detecting transcripts | |
| 980-CDS-CUT-1F | ATGGCGACTCCCGCCGCT |
| 980-CDS-CUT-1R | CTAAAACAAAATTTTATCCTGG |
| Primers for transgenic and CRISPR constructs | |
| w81-G-F | CCATGATTACGAATTCGTGGGGACAGGTTGAGTGG |
| w81-G-R | TACCGAGCTCGAATTCTTTCATCGCTGGGTTGTCG |
| CP-980-F | AGATGATCCGTGGCACCCCGACTTCTACCGGATGTTTTAGAGCTATGC |
| CP-980-R | GCATAGCTCTAAAACATCCGGTAGAAGTCGGGGTGCCACGGATCATCT |
| OsPAP7-GFP-InXbaI | CGGAGCTAGCTCTAGAATGGCGACTCCCGCCGCTTC |
| OsPAP7-GFP-InBamHI | TGCTCACCATGGATCCAAACAAAATTTTATCCTGGTAG |
| Ospap7-GFP-InXbaI | CGGAGCTAGCTCTAGAATGGCGACTCCCGCC |
| Ospap7-GFP-InBamHI | TGCTCACCATGGATCCAATAGTACAGAAGTGTACC |
| Primers for Y2H | |
| BD-OsPAP7-F | CATGGAGGCCGAATTCATGGCGACTCCCGCCGCT |
| BD-OsPAP7-R | CAGGTCGACGGATCCCCTAAAACAAAATTTTATCCT |
| BD-OsPAP5-F | CATGGAGGCCGAATTCATCCTGTCCCCTTCCGAA |
| BD-OsPAP5-R | CAGGTCGACGGATCCCTCAGCAACTTGATCCTTATAT |
| AD-OsPAP1-F | CAGATTACGCTCATATGATGGCCACCCCTACCCCCAC |
| AD-OsPAP1-R | CACCCGGGTGGAATTCTTACTCCTCTGCAGGTGGCGGTT |
| AD-OsPAP2-F | CAGATTACGCTCATATGATGAAGGCCTCCGGCGTCG |
| AD-OsPAP2-R | CACCCGGGTGGAATTCTCACAGTGCAAGGAGTTCT |
| AD-OsPAP3-F | CAGATTACGCTCATATGATGGCGGCCACTCCGGCC |
| AD-OsPAP3-R | CACCCGGGTGGAATTCTCATTTTGTGATGAAACAC |
| AD-OsPAP4-F | CAGATTACGCTCATATGATGGCGGCTTTCGCCTC |
| AD-OsPAP4-R | CACCCGGGTGGAATTCTCATGCAACTGGGATATTTGG |
| AD-OsPAP5-F | CAGATTACGCTCATATGATGGCGTCGTGCTCCCG |
| AD-OsPAP5-R | CACCCGGGTGGAATTCTTACTCATCTTCTTCGAAGTCCAT |
| AD-OsPAP6-F | CAGATTACGCTCATATGATGGCCATGGCGGCCTC |
| AD-OsPAP6-R | CACCCGGGTGGAATTCTTAGATGAAAACCATCCAGATAAT |
| AD-OsPAP7-F | CAGATTACGCTCATATGATGGCGACTCCCGCCG |
| AD-OsPAP7-R | CACCCGGGTGGAATTCCTAAAACAAAATTTTATCCTGGTAG |
| AD-OsPAP8-F | CAGATTACGCTCATATGATGGCGGCCACCGTGT |
| AD-OsPAP8-R | CACCCGGGTGGAATTCTCAGAACCAATTCGAGTAGTCAAAG |
| AD-OsPAP9-F | CAGATTACGCTCATATGATGGCGTTCGCCACACTG |
| AD-OsPAP9-R | CACCCGGGTGGAATTCCACACCCCTAGGGACTTCTCT |
| AD-OsPAP10-F | CAGATTACGCTCATATGATGGCCATGGCCGCGG |
| AD-OsPAP10-R | CACCCGGGTGGAATTCTCACAATTCATTATCAATGATATTT |
| Primers for BiFC | |
| BiFC-N-PAP5-BamH-F | AACTAGTGGAGGATCCATGGCGTCGTGCTCCCGCA |
| BiFC-N-PAP5-BamH-R | ACCCACCTCCGGATCCCTCATCTTCTTCGAAGTCC |
| BiFC-C-PAP7-PmlI-F | CCTACGTAGTCACGTGATGGCGACTCCCGCCGCT |
| BiFC-C-PAP7-PmlI-F | CTCCGGACGTCACGTGAAACAAAATTTTATCCTGG |
| Primers for qRT-PCR | |
| OsPAP7-RT-F | CTTCTCGAGTGATGCCAAGA |
| OsPAP7-RT-R | ATGCAGCATTGTGGTAGAGC |
| HEMA1-F | CACCAGTCTGAATCATAT |
| HEMA1-R | CTACCACTTCTCTAATCC |
| CAB1R-F | AGACGTTCGCCAAGAACC |
| CAB1R-R | GAGGAGCTCCGGGAAGAC |
| CAB2R-F | GTTCTCCATGTTCGGCTTCT |
| CAB2R-R | GACGAAGTTGGTGGCGTAG |
| psaA-F | GAGATACCACTTCCTCAT |
| psaA-R | ACTAAGAAATTCTGCGTATT |
| psbA-F | AAGTTTCTCTGATGGTATG |
| psbA-R | ATAGCACTGAATAGGGAA |
| rbcS-F | TCATCAGCTTCATCGCCTAC |
| rbcS-R | ACTGGGAACACACGAAACAA |
| rbcL-F | GTTGAAAGGGATAAGTTGA |
| rbcL-R | AATGGTTGTGAGTTTACG |
| clpP-F | GATACGATGCAAACGGTGAC |
| clpP-R | GGGAATGCTATACGCTTGGT |
| rrn16-F | GGAGAAGAAGCAATGACGGT |
| rrn16-R | GATCATTCCGGATAACGCTT |
| rpoA-F | AAATCGTTGATACGGCACAA |
| rpoA-R | ATTCACATTTCGAACAGGCA |
| rpoB-F | GCATTGTTGGAACTGGATTG |
| rpoB-R | GCCGATGGGTAACTAAAGGA |
| rpoC1-F | TTGTGGGTCCTTCACTTTCA |
| rpoC1-R | TTCCCATACAATGGGTTCCT |
| rpoC2-F | GAGGACGAACATGGGACTTT |
| rpoC2-R | GTTCCTCGATGCTCAATCAA |
| Ubq-F | GCTCCGTGGCGGTATCAT |
| Ubq-R | CGGCAGTTGACAGCCCTAG |
| Genotype | Plant Height (cm) | Number of Tillers per Plant | Branch Number per Panicle | Number of Spikelets per Panicle | 1000-Grain Weight (g) |
|---|---|---|---|---|---|
| Wild type | 83.0 ± 3.4 | 9.9 ± 1.7 | 9.3 ± 1.6 | 122.8 ± 22.2 | 24.5 ± 0.1 |
| w81 | 82.5 ± 2.9 | 8.9± 1.4 | 8.7 ± 1.4 | 123.2 ± 22.7 | 25.1 ± 0.1 |
| Cross | Normal | Yellow | Ratio 3:1 a |
|---|---|---|---|
| Wild type | 638 | 203 | 2.45 |
| w81 | 577 | 189 | 1.49 |
Appendix B



References
- Miyoshi, K.; Ito, Y.; Serizawa, A.; Kurata, N. OsHAP3 genes regulate chloroplast biogenesis in rice. Plant J. 2003, 36, 532–540. [Google Scholar] [CrossRef]
- Gothandam, K.M.; Kim, E.-S.; Cho, H.; Chung, Y.-Y. OsPPR1, a pentatricopeptide repeat protein of rice is essential for the chloroplast biogenesis. Plant Mol. Biol. 2005, 58, 421–433. [Google Scholar] [CrossRef]
- Lv, Y.S.; Shao, G.N.; Qiu, J.H.; Jiao, G.A.; Sheng, Z.H.; Xie, L.H.; Wu, Y.W.; Tang, S.Q.; Wei, X.J.; Hu, P.S. White Leaf and Panicle 2, encoding a PEP-associated protein, is required for chloroplast biogenesis under heat stress in rice. J. Exp. Bot. 2017, 68, 5147–5160. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ren, Y.L.; Zhou, K.N.; Liu, L.L.; Wang, J.L.; Xu, Y.; Zhang, H.; Zhang, L.; Feng, Z.M.; Wang, L.W.; et al. WHITE STRIPE LEAF4 encodes a novel p-type ppr protein required for chloroplast biogenesis during early leaf development. Front. Plant Sci. 2017, 8, 1116. [Google Scholar] [CrossRef]
- Liu, Z.W.; Wang, Z.Y.; Gu, H.; You, J.; Hu, M.M.; Zhang, Y.J.; Zhu, Z.; Wang, Y.H.; Liu, S.J.; Chen, L.M.; et al. Identification and phenotypic characterization of ZEBRA LEAF16 encoding a β-Hydroxyacyl-ACP dehydratase in rice. Front. Plant Sci. 2018, 9, 782. [Google Scholar] [CrossRef]
- Lee, S.; Joung, Y.H.; Kim, J.-K.; Choi, Y.D.; Jang, G. An isoform of the plastid RNA polymerase associated protein FSD3 neg-atively regulates chloroplast development. BMC Plant Biol. 2019, 19, 524. [Google Scholar] [CrossRef] [PubMed]
- Liebers, M.; Cozzi, C.; Uecker, F.; Chambon, L.; Blanvillain, R.; Pfannschmidt, T. Biogenic signals from plastids and their role in chloroplast development. J. Exp. Bot. 2022, 73, 7105–7125. [Google Scholar] [CrossRef] [PubMed]
- Hajdukiewicz, P.T.; Allison, L.A.; Maliga, P. The two RNA polymerases encoded by the nuclear and the plastid compartments transcribe distinct groups of genes in tobacco plastids. EMBO J. 1997, 16, 4041–4048. [Google Scholar] [CrossRef]
- Liere, K.; Weihe, A.; Börner, T. The transcription machineries of plant mitochondria and chloroplasts: Composition, function, and regulation. J. Plant Physiol. 2011, 168, 1345–1360. [Google Scholar] [CrossRef]
- Pfannschmidt, T.; Blanvillain, R.; Merendino, L.; Courtois, F.; Chevalier, F.; Liebers, M.; Grübler, B.; Hommel, E.; Lerbs-Mache, S. Plastid RNA polymerases: Orchestration of enzymes with different evolutionary origins controls chloroplast biogenesis during the plant life cycle. J. Exp. Bot. 2015, 66, 6957–6973. [Google Scholar] [CrossRef]
- Hedtke, B.; Borner, T.; Weihe, A. Mitochondrial and chloroplast phage-type RNA polymerases in Arabidopsis. Science 1997, 277, 809–811. [Google Scholar] [CrossRef]
- Yu, Q.-B.; Huang, C.; Yang, Z.-N. Nuclear-encoded factors associated with the chloroplast transcription machinery of higher plants. Front. Plant Sci. 2014, 5, 316. [Google Scholar] [CrossRef]
- Zhelyazkova, P.; Sharma, C.M.; Förstner, K.U.; Liere, K.; Vogel, J.; Börner, T. The primary transcriptome of barley chloro-plasts: Numerous noncoding RNAs and the dominating role of the plastid-encoded RNA polymerase. Plant Cell 2012, 24, 123–136. [Google Scholar] [CrossRef]
- Börner, T.; Aleynikova, A.Y.; Zubo, Y.O.; Kusnetsov, V.V. Chloroplast RNA polymerases: Role in chloroplast biogenesis. Biochim. Biophys. Acta 2015, 1847, 761–769. [Google Scholar] [CrossRef]
- Legen, J.; Kemp, S.; Krause, K.; Profanter, B.; Herrmann, R.G.; Maier, R.M. Comparative analysis of plastid transcription profiles of entire plastid chromosomes from tobacco attributed to wild-type and PEP-deficient transcription machineries. Plant J. 2002, 31, 171–188. [Google Scholar] [CrossRef]
- Pfalz, J.; Pfannschmidt, T. Essential nucleoid proteins in early chloroplast development. Trends Plant Sci. 2013, 18, 186–194. [Google Scholar] [CrossRef]
- Steiner, S.; Schröter, Y.; Pfalz, J.; Pfannschmidt, T. Identification of essential subunits in the plastid-encoded RNA polymerase complex reveals building blocks for proper plastid development. Plant Physiol. 2011, 157, 1043–1055. [Google Scholar] [CrossRef]
- Majeran, W.; Friso, G.; Asakura, Y.; Qu, X.; Huang, M.; Ponnala, L.; Watkins, K.P.; Barkan, A.; van Wijk, K.J. Nucleoid-enriched proteomes in developing plastids and chloroplasts from maize leaves: A new conceptual framework for nucleoid functions. Plant Physiol. 2012, 158, 156–189. [Google Scholar] [CrossRef]
- Melonek, J.; Oetke, S.; Krupinska, K. Multifunctionality of plastid nucleoids as revealed by proteome analyses. Biochim. Biophys. Acta 2016, 1864, 1016–1038. [Google Scholar] [CrossRef]
- Pfalz, J.; Liere, K.; Kandlbinder, A.; Dietz, K.-J.; Oelmüller, R. pTAC2, -6, and -12 Are Components of the Transcriptionally Active Plastid Chromosome That Are Required for Plastid Gene Expression. Plant Cell 2005, 18, 176–197. [Google Scholar] [CrossRef]
- Ji, Y.; Lehotai, N.; Zan, Y.J.; Dubreuil, C.; Díaz, M.G.; Strand, Å. A fully assembled plastid-encoded RNA polymerase complexdetected in etioplasts and proplastids in Arabidopsis. Physiol. Plant. 2021, 171, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Yagi, Y.; Ishizaki, Y.; Nakahira, Y.; Tozawa, Y.; Shiina, T. Eukaryotic-type plastid nucleoid protein pTAC3 is essential for transcription by the bacterial-type plastid RNA polymerase. Proc. Natl. Acad. Sci. USA 2012, 109, 7541–7546. [Google Scholar] [CrossRef] [PubMed]
- Myouga, F.; Hosoda, C.; Umezawa, T.; Iizumi, H.; Kuromori, T.; Motohashi, R.; Shono, Y.; Nagata, N.; Ikeuchi, M.; Shinozaki, K. A hetero complex of iron superoxide dismutases defends chloroplast nucleoids against oxidative stress and is essential for chloroplast development in Arabidopsis. Plant Cell 2008, 20, 3148–3162. [Google Scholar] [CrossRef]
- Gao, Z.P.; Yu, Q.B.; Zhao, T.T.; Ma, Q.; Chen, G.X.; Yang, Z.N. A functional component of the transcriptionally active chromosome complex, Arabidopsis pTAC14, interacts with pTAC12/HEMERA and regulates plastid gene expression. Plant Physiol. 2011, 157, 1733–1745. [Google Scholar] [CrossRef] [PubMed]
- Arsova, B.; Hoja, U.; Wimmelbacher, M.; Greiner, E.; Üstün, Ş.; Melzer, M.; Petersen, K.; Lein, W.; Börnke, F. Plastidial thioredoxin z interacts with two fructokinase-like proteins in a thiol-dependent mmanner: Evidence for an essential role in chloroplast development in Arabidopsis and Nicotiana benthamiana. Plant Cell 2010, 22, 1498–1515. [Google Scholar] [CrossRef]
- Huang, C.; Yu, Q.-B.; Lv, R.-H.; Yin, Q.-Q.; Chen, G.-Y.; Xu, L.; Yang, Z.-N. The reduced plastid-encoded polymerase-dependent plastid gene expression leads to the delayed greening of the Arabidopsis fln2 mutant. PLoS ONE 2013, 8, e73092. [Google Scholar] [CrossRef]
- Yu, Q.-B.; Lu, Y.; Ma, Q.; Zhao, T.-T.; Huang, C.; Zhao, H.-F.; Zhang, X.-L.; Lv, R.-H.; Yang, Z.-N. TAC7, an essential component of the plastid transcriptionally active chromosome complex, interacts with FLN1, TAC10, TAC12 and TAC14 to regulate chloroplast gene expression in Arabidopsis thaliana. Physiol. Plant. 2012, 148, 408–421. [Google Scholar] [CrossRef]
- Chang, S.H.; Lee, S.; Um, T.Y.; Kim, J.-K.; Choi, Y.D.; Jang, G. pTAC10, a Key Subunit of Plastid-Encoded RNA Polymerase, Promotes Chloroplast Development. Plant Physiol. 2017, 174, 435–449. [Google Scholar] [CrossRef]
- Pfalz, J.; Holtzegel, U.; Barkan, A.; Weisheit, W.; Mittag, M.; Pfannschmidt, T. ZmpTAC12 binds single-stranded nucleic acids and is essential for accumulation of the plastid-encoded polymerase complex in maize. New Phytol. 2015, 206, 1024–1037. [Google Scholar] [CrossRef]
- Liu, L.L.; You, J.; Zhu, Z.; Chen, K.Y.; Hu, M.M.; Gu, H.; Liu, Z.W.; Wang, Z.Y.; Wang, Y.H.; Liu, S.J.; et al. WHITE STRIPE LEAF8, encoding a deoxyribonucleoside kinase, is involved in chloroplast development in rice. Plant Cell Rep. 2020, 39, 19–33. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Ren, Y.; Lv, J.; Wang, Y.; Liu, F.; Zhou, F.; Zhao, S.; Chen, S.; Peng, C.; Zhang, X.; et al. Young Leaf Chlorosis 1, a chloroplast-localized gene required for chlorophyll and lutein accumulation during early leaf development in rice. Planta 2013, 237, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zhang, X.; He, B.; Diao, L.; Sheng, S.; Wang, J.; Guo, X.; Su, N.; Wang, L.; Jiang, L.; et al. A Chlorophyll-Deficient Rice Mutant with Impaired Chlorophyllide Esterification in Chlorophyll Biosynthesis. Plant Physiol. 2007, 145, 29–40. [Google Scholar] [CrossRef]
- Hiei, Y.; Ohta, S.; Komari, T.; Kumashiro, T. Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J. 1994, 6, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Waadt, R.; Kudla, J. In Planta Visualization of Protein Interactions Using Bimolecular Fluorescence Complementation (BiFC). CSH Protoc. 2008, 2008, pdb.prot4995. [Google Scholar] [CrossRef]
- Yoo, S.-D.; Cho, Y.-H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef]
- Livak, K.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kang, S.G.; Lee, K.E.; Singh, M.; Kumar, P.; Matin, M.N. Rice lesion mimic mutants (LMM): The current understanding of genetic mu tations in the failure of ROS scavenging during lesion formation. Plants 2021, 10, 1598. [Google Scholar] [CrossRef]
- Garcia, M.; Myouga, F.; Takechi, K.; Sato, H.; Nabeshima, K.; Nagata, N.; Takio, S.; Shinozaki, K.; Takano, H. An Arabidopsis homolog of the bacterial peptidoglycan synthesis enzyme MurE has an essential role in chloroplast development. Plant J. 2008, 53, 924–934. [Google Scholar] [CrossRef]
- Liebers, M.; Gillet, F.X.; Israel, A.; Pounot, K.; Chambon, L.; Chieb, M.; Chevalier, F.; Ruedas, R.; Favier, A.; Gans, P.; et al. Nucleo-plastidic PAP8/pTAC6 couples chloroplast formation with photomorphogenesis. EMBO J. 2020, 39, e104941. [Google Scholar] [CrossRef]
- Chambon, L.; Gillet, F.-X.; Chieb, M.; Cobessi, D.; Pfannschmidt, T.; Blanvillain, R. PAP8/pTAC6 is part of a nuclear protein complex and displays rna recognition motifs of viral origin. Int. J. Mol. Sci. 2022, 23, 3059. [Google Scholar] [CrossRef]
- Grübler, B.; Merendino, L.; Twardziok, S.O.; Mininno, M.; Allorent, G.; Chevalier, F.; Liebers, M.; Blanvillain, R.; Mayer, K.F.X.; Lerbs-Mache, S.; et al. Light and plastid signals regulate different sets of genes in the albino mutant Pap7-1. Plant Physiol. 2017, 175, 1203–1219. [Google Scholar] [CrossRef] [PubMed]
- Leister, D. Chloroplast research in the genomic age. Trends Genet. 2003, 19, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Lin, Q.; Zhou, C.; Liu, X.; Miao, R.; Ma, T.; Chen, Y.; Mou, C.; Jing, R.; Feng, M.; et al. Young Leaf White Stripe encodes a P-type PPR protein required for chloroplast development. J. Integr. Plant Biol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Liu, Z.W.; Gu, H. Phenotypic identification and candidate gene analysis of rice white stripe leaf mutant wsl7. J. Nanjing Agric. Univ. 2019, 42, 21–29. [Google Scholar]
- Jiang, H.; Zhang, A.; Ruan, B.; Hu, H.; Guo, R.; Chen, J.; Qian, Q.; Gao, Z. Identification of Green-Revertible Yellow 3 (GRY3), encoding a 4-hydroxy-3-methylbut-2-enyl diphosphate reductase involved in chlorophyll synthesis under high temperature and high light in rice. Crops J. 2023; in press. [Google Scholar] [CrossRef]
- Foyer, C.H.; Lopez-Delgado, H.; Dat, J.F.; Scott, I.M. Hydrogen peroxide- and glutathione-associated mechanisms of acclimatory stress tolerance and signalling. Physiol. Plant. 1997, 100, 241–254. [Google Scholar] [CrossRef]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.; Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ. 2012, 35, 259–270. [Google Scholar] [CrossRef]
- Saed-Moucheshi, A.; Shekoofa, A.; Pessarakli, M. Reactive Oxygen Species (ROS) Generation and Detoxifying in Plants. J. Plant Nutr. 2014, 37, 1573–1585. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K.; Gollery, M.; Shulaev, V.; Breusegem, V.F. ROS signaling: The new wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef]
- Lu, T.; Meng, Z.; Zhang, G.; Qi, M.; Sun, Z.; Liu, Y.; Li, T. Sub-high Temperature and High Light Intensity Induced Irreversible Inhibition on Photosynthesis System of Tomato Plant (Solanum lycopersicum L.). Front. Plant Sci. 2017, 8, 365. [Google Scholar] [CrossRef]
- Kremnev, D.; Strand, A. Plastid encoded RNA polymerase activity and expression of photosynthesis genes required for embryo and seed development in Arabidopsis. Front. Plant Sci. 2014, 5, 385. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, C.; Wang, Y.; Niu, M.; Ren, Y.; Zhou, K.; Zhang, H.; Lin, Q.; Wu, F.; Cheng, Z.; et al. WSL3, a component of the plastid-encoded plastid RNA polymerase, is essential for early chloroplast development in rice. Plant Mol. Biol. 2016, 92, 581–595. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Shen, L.; Wang, Z.; Hu, G.; Ren, D.; Hu, J.; Zhu, L.; Gao, Z.; Zhang, G.; Guo, L.; et al. OsCAF1, a CRM Domain Containing Protein, Influences Chloroplast Development. Int. J. Mol. Sci. 2019, 20, 4386. [Google Scholar] [CrossRef] [PubMed]
- Demarsy, E.; Courtois, F.; Azevedo, J.; Buhot, L.; Lerbs-Mache, S. Building Up of the Plastid Transcriptional Machinery during Germination and Early Plant Development. Plant Physiol. 2006, 142, 993–1003. [Google Scholar] [CrossRef] [PubMed]
- Kusumi, K.; Iba, K. Establishment of the chloroplast genetic system in rice during early leaf development and at low tempera tures. Front. Plant Sci. 2014, 5, 386. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, S.; Wang, Y.; Ding, X.; Tian, Y.; Wu, Z.; Li, H.; Li, Q.; Wang, Y.; Zhou, S.; Dong, X.; et al. A Plastid RNA Polymerase-Associated Protein Is Involved in Early Chloroplast Development in Rice. Agronomy 2023, 13, 1424. https://doi.org/10.3390/agronomy13051424
Song S, Wang Y, Ding X, Tian Y, Wu Z, Li H, Li Q, Wang Y, Zhou S, Dong X, et al. A Plastid RNA Polymerase-Associated Protein Is Involved in Early Chloroplast Development in Rice. Agronomy. 2023; 13(5):1424. https://doi.org/10.3390/agronomy13051424
Chicago/Turabian StyleSong, Shuang, Ying Wang, Xin Ding, Yunlu Tian, Zewan Wu, Hang Li, Qing Li, Yunpeng Wang, Shirong Zhou, Xiaoou Dong, and et al. 2023. "A Plastid RNA Polymerase-Associated Protein Is Involved in Early Chloroplast Development in Rice" Agronomy 13, no. 5: 1424. https://doi.org/10.3390/agronomy13051424