
Evaluation of Vegetative Development of Quinoa under Water Stress by Applying Different Organic Amendments


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Layout and Characteristics of Soil and Organic Amendments
2.2. Experimental Set-up and Conditions
2.3. Plant Measurements
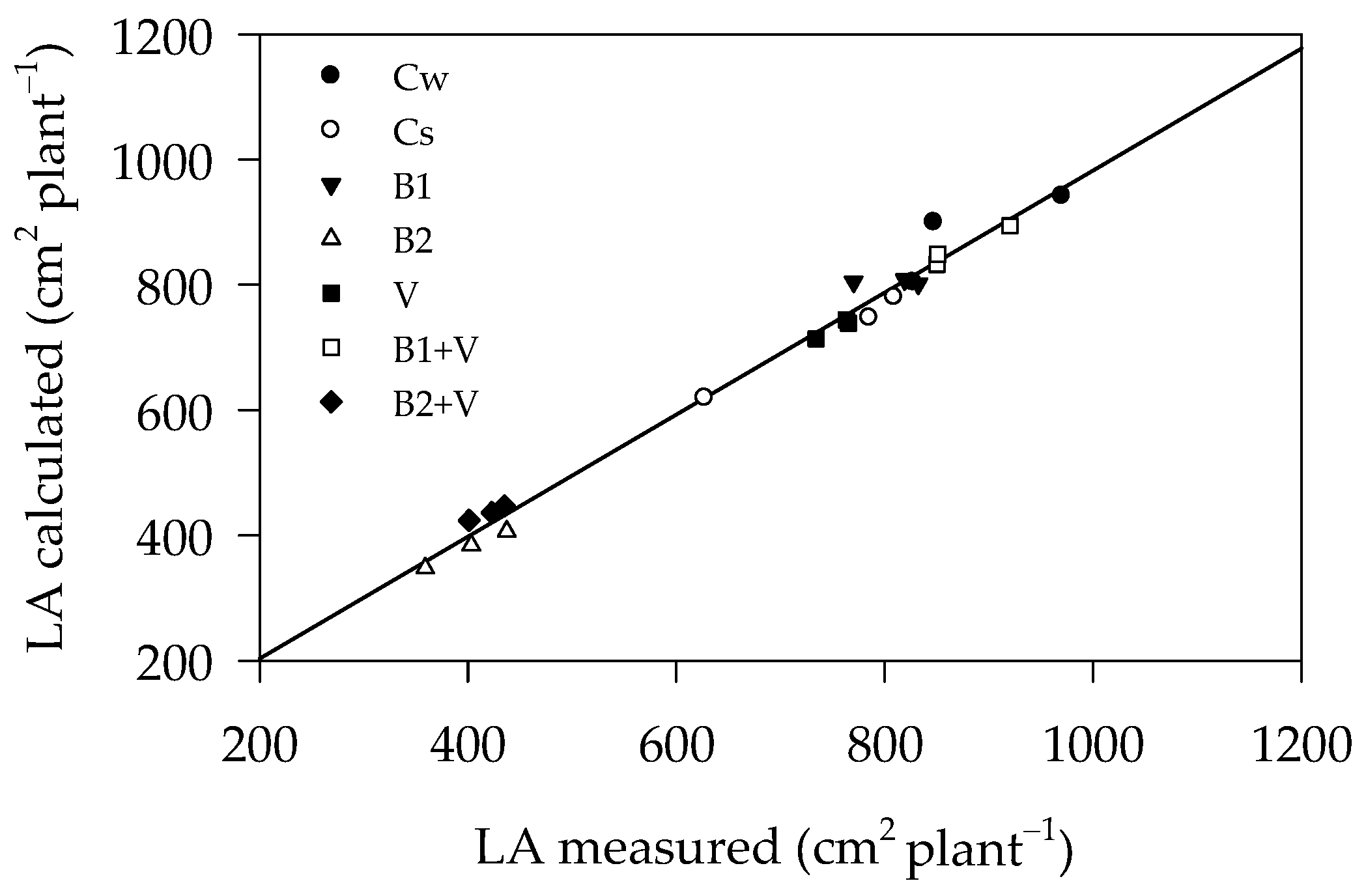
2.3.1. Growth-Related Parameters
2.3.2. Water-Related Parameters
2.4. Statistical Analysis
3. Results
3.1. Phenological Stages
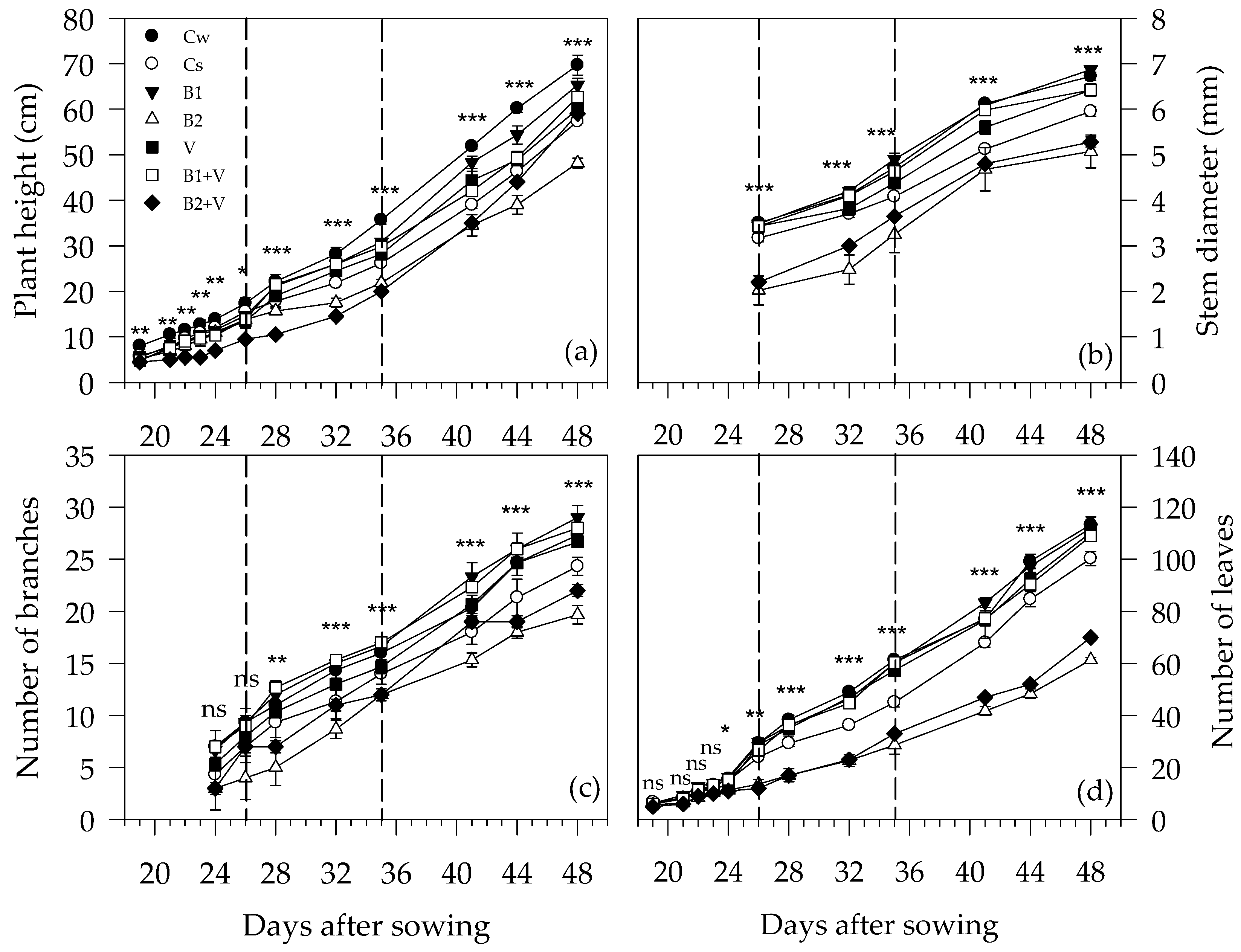
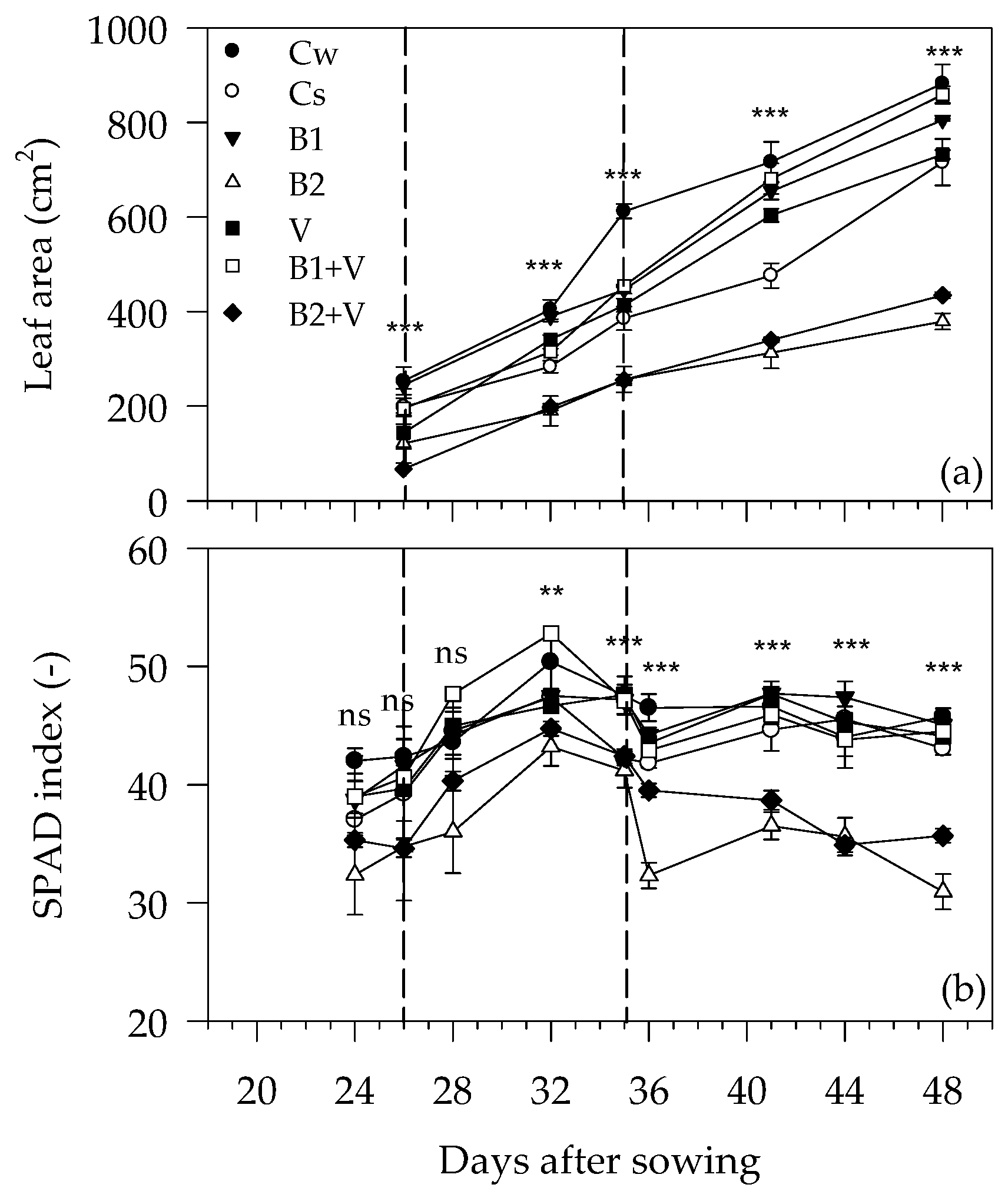
3.2. Growth-Related Parameters during the Experiment
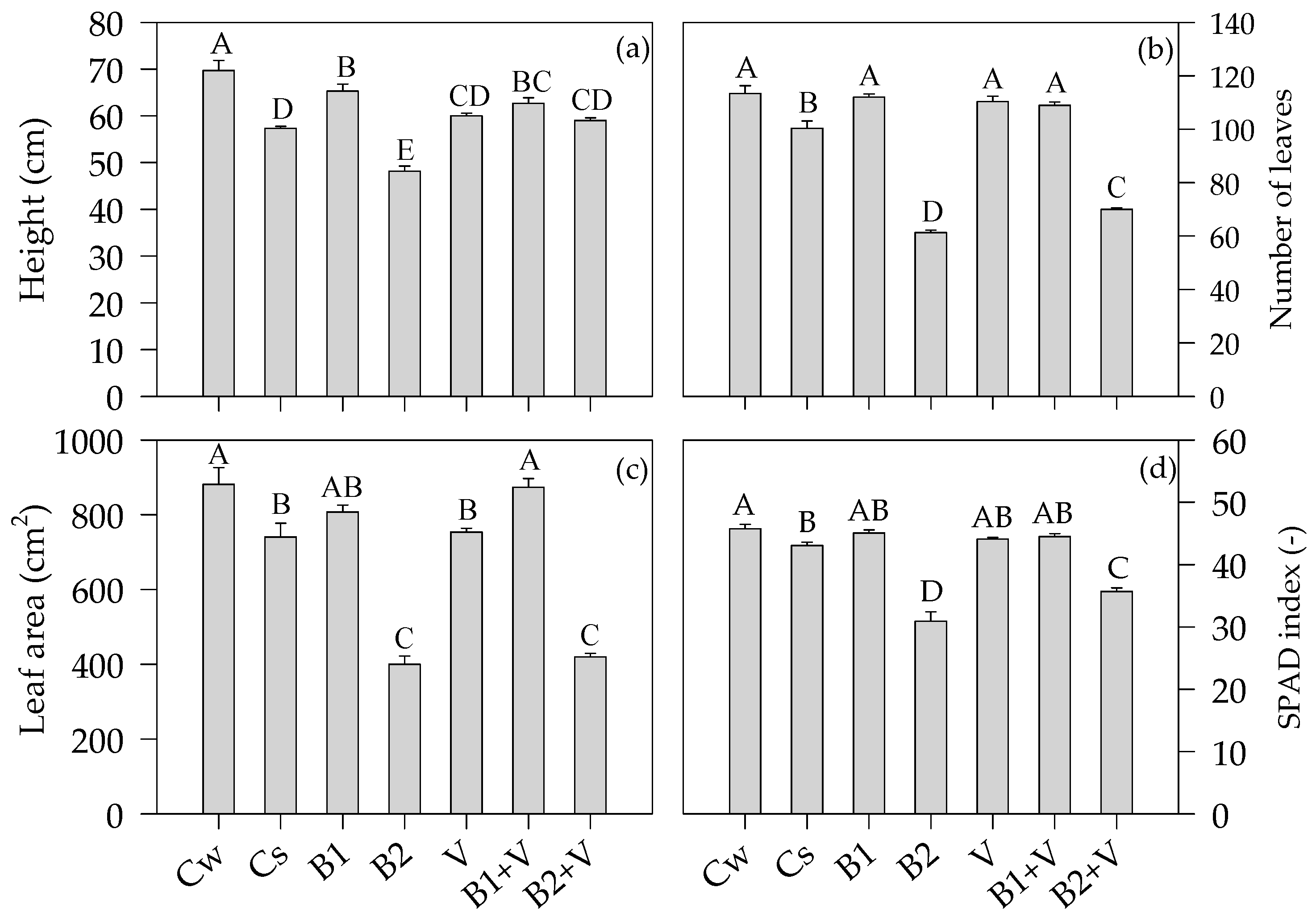
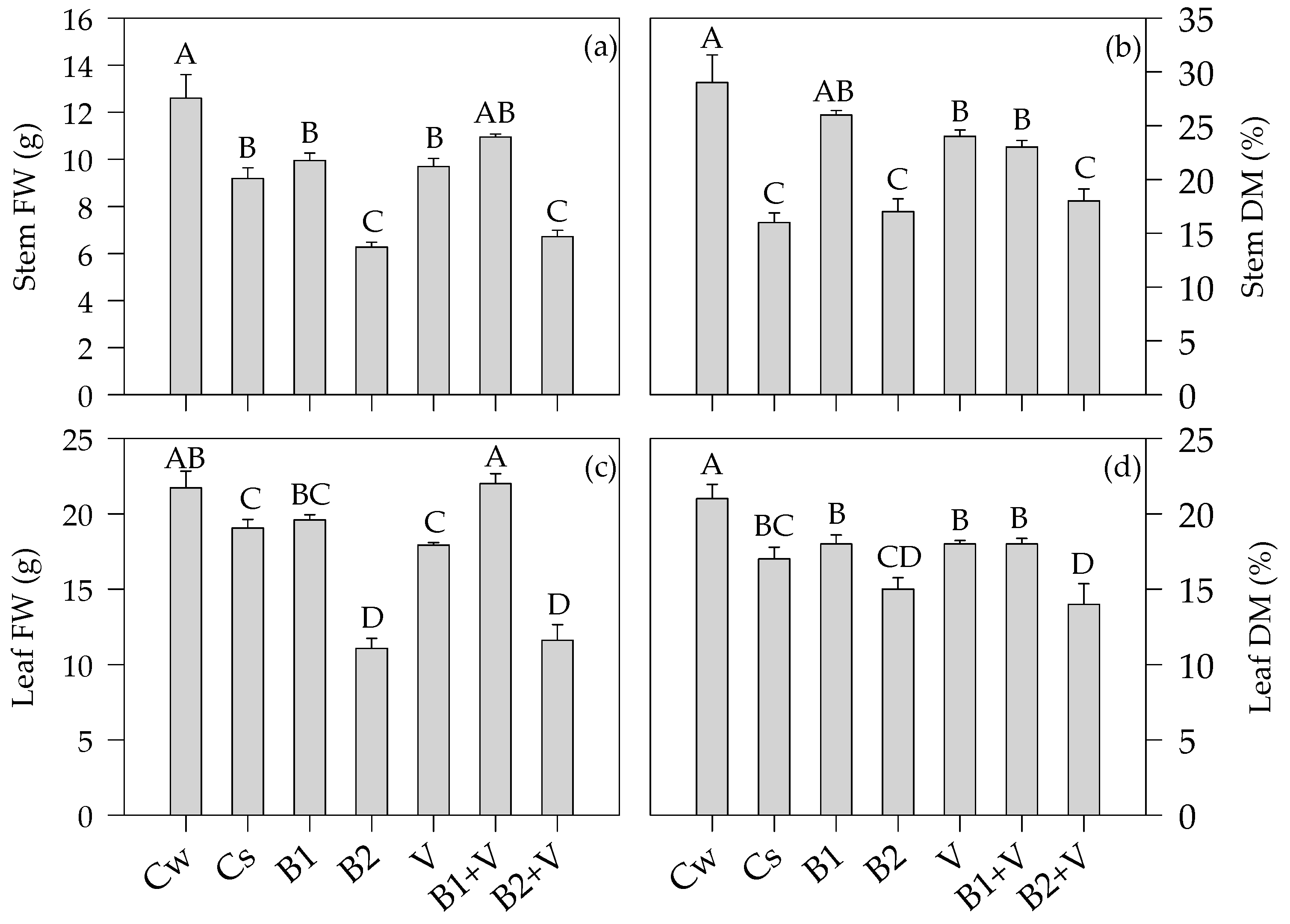
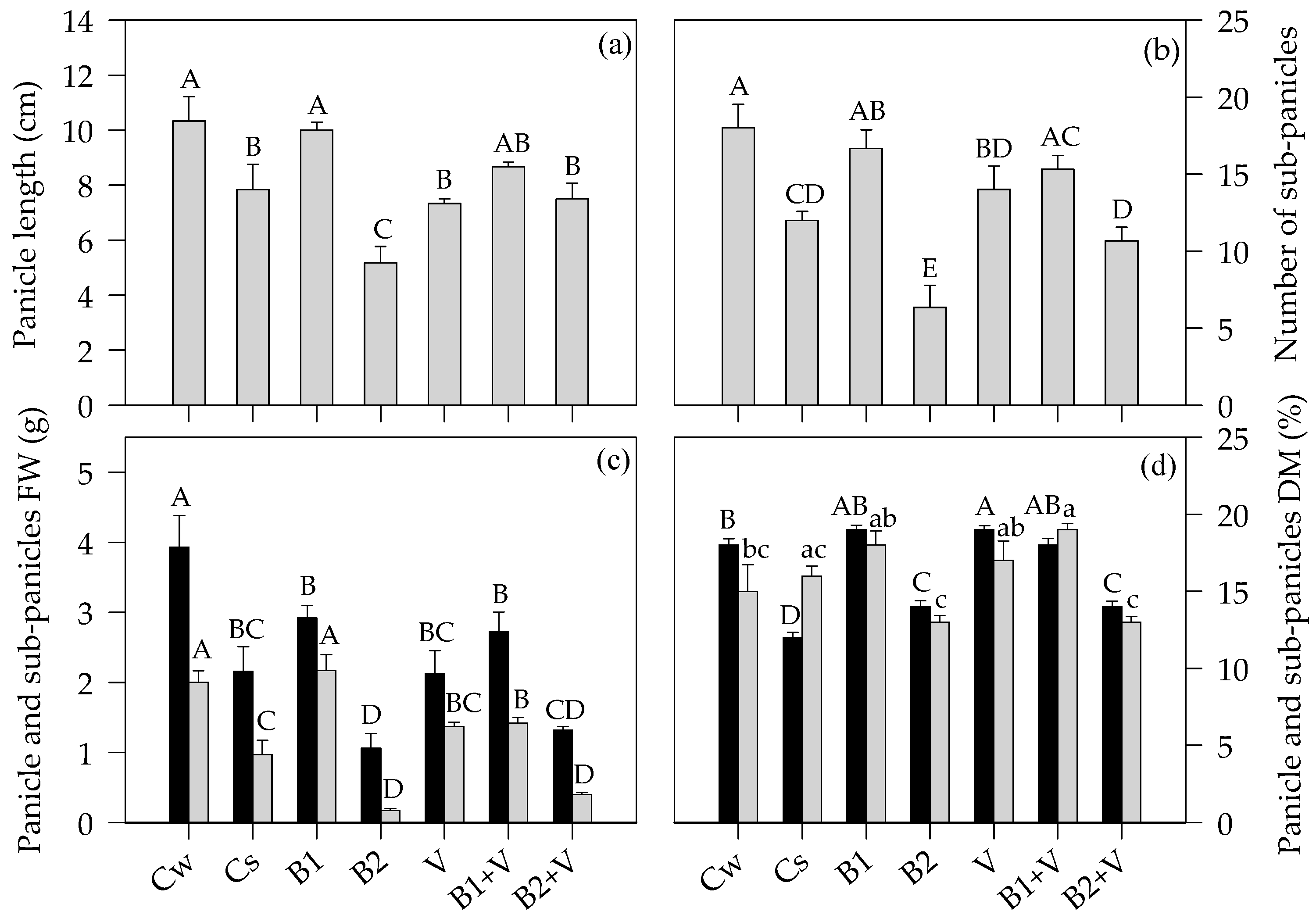
3.3. Growth-Related Parameters at the End of the Experiment
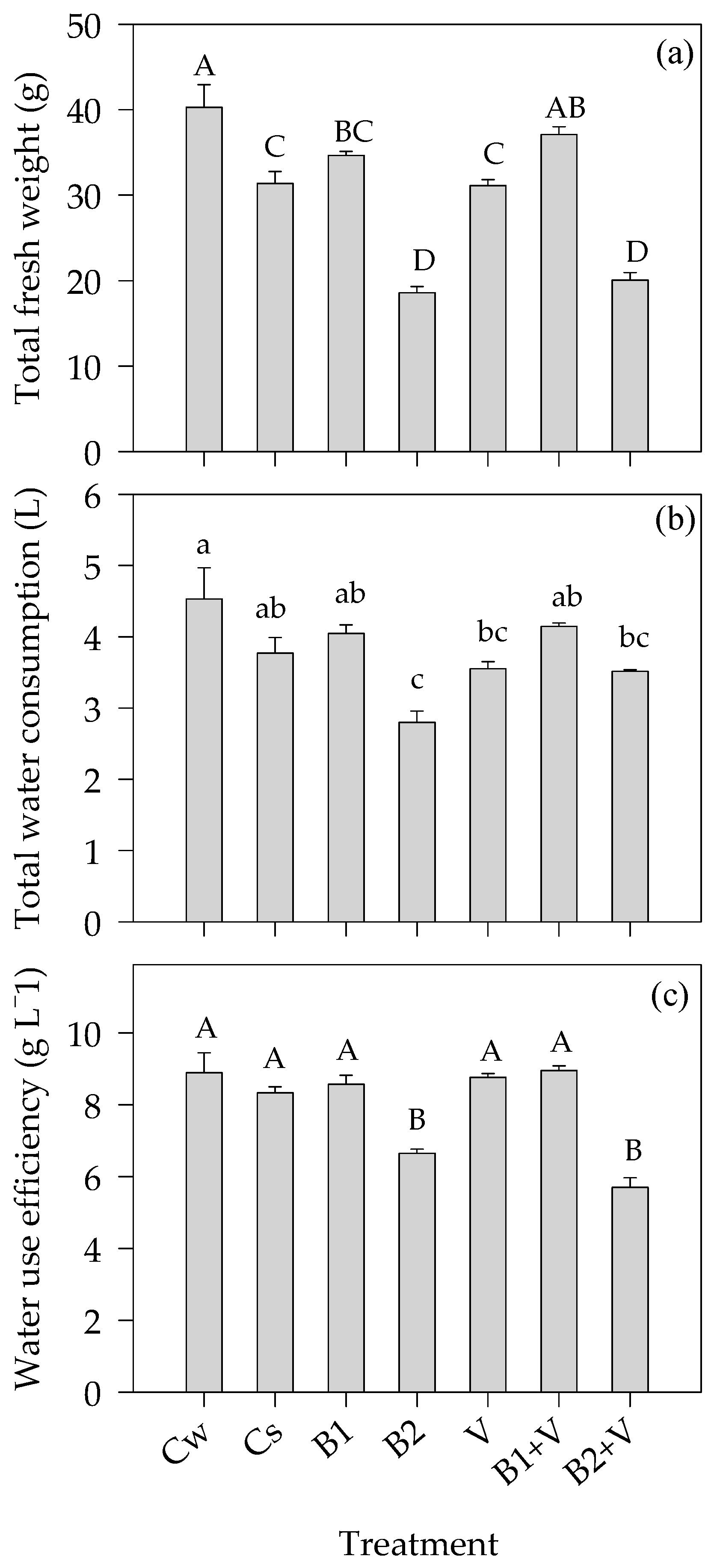
3.4. Water-Related Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ain, Q.T.; Siddique, K.; Bawazeer, S.; Ali, I.; Mazhar, M.; Rasool, R.; Mubeen, B.; Ullah, F.; Unar, A.; Jafar, T.H. Adaptive mechanisms in quinoa for coping in stressful environments: An update. PeerJ 2023, 11, e14832. [Google Scholar] [CrossRef] [PubMed]
- Akram, M.Z.; Basra, S.M.A.; Hafeez, M.B.; Khan, S.; Nazeer, S.; Iqbal, S.; Saddiq, M.S.; Zahra, N. Adaptability and yield potential of new quinoa lines under agro-ecological conditions of Faisalabad-Pakistan. Asian J. Agric. Biol. 2021, 2, 202005301. [Google Scholar] [CrossRef]
- Hafeez, M.B.; Iqbal, S.; Li, Y.; Saddiq, M.S.; Basra, S.M.; Zhang, H.; Zahra, N.; Akram, M.Z.; Bertero, D.; Curti, R.N. Assessment of phenotypic diversity in the USDA collection of quinoa links genotypic adaptation to germplasm origin. Plants 2022, 11, 738. [Google Scholar] [CrossRef] [PubMed]
- Bazile, D.; Bertero, H.D.; Nieto, C. State of the Art Report on Quinoa around the World in 2013; FAO Headquarter: Rome, Italy, 2015. [Google Scholar]
- FAO. Home-International Year of Quinoa 2013; FAO Headquarters: Rome, Italy, 2013; Available online: http://www.fao.org/quinoa-2013/en (accessed on 22 April 2023).
- Wu, Q.; Bai, X.; Zhao, W.; Shi, X.; Xiang, D.; Wan, Y.; Wu, X.; Sun, Y.; Zhao, J.; Peng, L. Investigation into the underlying regulatory mechanisms shaping inflorescence architecture in Chenopodium quinoa. BMC Genom. 2019, 20, 658. [Google Scholar] [CrossRef] [PubMed]
- Voronov, S.; Pleskachiov, Y.; Shitikova, A.; Zargar, M.; Abdelkader, M. Diversity of the Biological and Proteinogenic Characteristics of Quinoa Genotypes as a Multi-Purpose Crop. Agronomy 2023, 13, 279. [Google Scholar] [CrossRef]
- Coţovanu, I.; Mironeasa, C.; Mironeasa, S. Nutritionally Improved Wheat Bread Supplemented with Quinoa Flour of Large, Medium and Small Particle Sizes at Typical Doses. Plants 2023, 12, 698. [Google Scholar] [CrossRef]
- Bastidas, E.; Roura, R.; Rizzolo, D.; Massanés, T.; Gomis, R. Quinoa (Chenopodium quinoa Willd), from nutritional value to potential health benefits: An integrative review. J. Nutr. Food Sci. 2016, 6, 497. [Google Scholar]
- Zurita Silva, A.; Jacobsen, S.-E.; Razzaghi, F.; Álvarez Flores, R.; Ruiz, K.B.; Morales, A.; Silva Ascencio, H. Quinoa Drought Responses and Adaptation. In State of the Art Report of Quinoa in the World in 2013 by FAO and CIRAD; FAO Headquarter: Roma, Italy, 2015; pp. 157–171. Available online: https://repositorio.uchile.cl/handle/2250/130689 (accessed on 22 April 2023).
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Skirycz, A.; Inzé, D. More from less: Plant growth under limited water. Curr. Opin. Biotechnol. 2010, 21, 197–203. [Google Scholar] [CrossRef]
- Jacobsen, S.-E.; Liu, F.; Jensen, C.R. Does root-sourced ABA play a role for regulation of stomata under drought in quinoa (Chenopodium quinoa Willd.). Sci. Hortic. 2009, 122, 281–287. [Google Scholar] [CrossRef]
- Nicotra, A.; Babicka, N.; Westoby, M. Seedling root anatomy and morphology: An examination of ecological differentiation with rainfall using phylogenetically independent contrasts. Oecologia 2002, 130, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Al-Naggar, A.; Abd El-Salam, R.; Badran, A.; El-Moghazi, M. Genotype and drought effects on morphological, physiological and yield traits of quinoa (Chenopodium quinoa Willd.). Asian J. Adv. Agric. Res. 2017, 3, 1–15. [Google Scholar] [CrossRef]
- Otterbach, S.L.; Khoury, H.; Rupasinghe, T.; Mendis, H.; Kwan, K.H.; Lui, V.; Natera, S.H.; Klaiber, I.; Allen, N.M.; Jarvis, D.E. Characterization of epidermal bladder cells in Chenopodium quinoa. Plant Cell Environ. 2021, 44, 3836–3852. [Google Scholar] [CrossRef] [PubMed]
- Shabala, L.; Mackay, A.; Tian, Y.; Jacobsen, S.E.; Zhou, D.; Shabala, S. Oxidative stress protection and stomatal patterning as components of salinity tolerance mechanism in quinoa (Chenopodium quinoa). Physiol. Plant. 2012, 146, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Garrido, M.; Silva, P.; Silva, H.; Muñoz, R.; Baginsky, C.; Acevedo, E. Evaluación del rendimiento de nueve genotipos de quinua (Chenopodium quinoa Willd. j bajo diferentes disponibilidades hídricas en ambiente mediterráneo. Idesia 2013, 31, 69–76. [Google Scholar] [CrossRef]
- Geerts, S.; Raes, D.; Garcia, M.; Mendoza, J.; Huanca, R. Crop water use indicators to quantify the flexible phenology of quinoa (Chenopodium quinoa Willd.) in response to drought stress. Field Crops Res. 2008, 108, 150–156. [Google Scholar] [CrossRef]
- Stikić, R.; Jovanović, Z.; Marjanović, M.; Djordjević, S. The effect of drought on water regime and growth of quinoa (Chenopodium quinoa Willd.). Ratar. I Povrt. 2015, 52, 80–84. [Google Scholar] [CrossRef]
- Beccari, G.; Quaglia, M.; Tini, F.; Pannacci, E.; Covarelli, L. Phytopathological threats associated with quinoa (Chenopodium quinoa Willd.) cultivation and seed production in an area of Central Italy. Plants 2021, 10, 1933. [Google Scholar] [CrossRef]
- Bilalis, D.; Roussis, I.; Kakabouki, I.; Folina, A. Quinoa (Chenopodium quinoa Willd.) crop under Mediterranean conditions: A review. Cienc. E Investig. Agrar. Rev. Latinoam. Cienc. Agric. 2019, 46, 51–68. [Google Scholar] [CrossRef]
- Das, S.K.; Ghosh, G.K.; Avasthe, R. Application of biochar in agriculture and environment, and its safety issues. Biomass Convers. Biorefin. 2020, 13, 1359–1369. [Google Scholar] [CrossRef]
- Sayara, T.; Basheer-Salimia, R.; Hawamde, F.; Sánchez, A. Recycling of organic wastes through composting: Process performance and compost application in agriculture. Agronomy 2020, 10, 1838. [Google Scholar] [CrossRef]
- Lee, J.; Sarmah, A.K.; Kwon, E.E. Production and formation of biochar. In Biochar from Biomass and Waste; Elsevier: Amsterdam, The Netherlands, 2019; pp. 3–18. [Google Scholar]
- Laird, D.A. The charcoal vision: A win–win–win scenario for simultaneously producing bioenergy, permanently sequestering carbon, while improving soil and water quality. Agron. J. 2008, 100, 178–181. [Google Scholar] [CrossRef]
- Lei, O.; Zhang, R. Effects of biochars derived from different feedstocks and pyrolysis temperatures on soil physical and hydraulic properties. J. Soils Sediments 2013, 13, 1561–1572. [Google Scholar] [CrossRef]
- Manolikaki, I.; Diamadopoulos, E. Agronomic potential of biochar prepared from brewery byproducts. J. Environ. Manag. 2020, 255, 109856. [Google Scholar] [CrossRef] [PubMed]
- Murtaza, G.; Ahmed, Z.; Usman, M.; Tariq, W.; Ullah, Z.; Shareef, M.; Iqbal, H.; Waqas, M.; Tariq, A.; Wu, Y. Biochar induced modifications in soil properties and its impacts on crop growth and production. J. Plant Nutr. 2021, 44, 1677–1691. [Google Scholar] [CrossRef]
- Subedi, R.; Bertora, C.; Zavattaro, L.; Grignani, C. Crop response to soils amended with biochar: Expected benefits and unintended risks. Ital. J. Agron. 2017, 12, 161–173. [Google Scholar] [CrossRef]
- Libutti, A.; Francavilla, M.; Monteleone, M. Hydrological properties of a clay loam soil as affected by biochar application in a pot experiment. Agronomy 2021, 11, 489. [Google Scholar] [CrossRef]
- Libutti, A.; Cammerino, A.R.B.; Francavilla, M.; Monteleone, M. Soil amendment with biochar affects water drainage and nutrient losses by leaching: Experimental evidence under field-grown conditions. Agronomy 2019, 9, 758. [Google Scholar] [CrossRef]
- Wang, D.; Li, C.; Parikh, S.J.; Scow, K.M. Impact of biochar on water retention of two agricultural soils—A multi-scale analysis. Geoderma 2019, 340, 185–191. [Google Scholar] [CrossRef]
- Abd El-Mageed, T.A.; Rady, M.M.; Taha, R.S.; Abd El Azeam, S.; Simpson, C.R.; Semida, W.M. Effects of integrated use of residual sulfur-enhanced biochar with effective microorganisms on soil properties, plant growth and short-term productivity of Capsicum annuum under salt stress. Sci. Hortic. 2020, 261, 108930. [Google Scholar] [CrossRef]
- Zhao, S.; Schmidt, S.; Gao, H.; Li, T.; Chen, X.; Hou, Y.; Chadwick, D.; Tian, J.; Dou, Z.; Zhang, W. A precision compost strategy aligning composts and application methods with target crops and growth environments can increase global food production. Nat. Food 2022, 3, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Adugna, G. A review on impact of compost on soil properties, water use and crop productivity. Acad. Res. J. Agric. Sci. Res. 2016, 4, 93–104. [Google Scholar]
- Azim, K.; Soudi, B.; Boukhari, S.; Perissol, C.; Roussos, S.; Thami Alami, I. Composting parameters and compost quality: A literature review. Org. Agric. 2018, 8, 141–158. [Google Scholar] [CrossRef]
- Rivier, P.-A.; Jamniczky, D.; Nemes, A.; Makó, A.; Barna, G.; Uzinger, N.; Rékási, M.; Farkas, C. Short-term effects of compost amendments to soil on soil structure, hydraulic properties, and water regime. J. Hydrol. Hydromech. 2022, 70, 74–88. [Google Scholar] [CrossRef]
- Abdelfattah, M.A.; Rady, M.M.; Belal, H.E.; Belal, E.E.; Al-Qthanin, R.; Al-Yasi, H.M.; Ali, E.F. Revitalizing fertility of nutrient-deficient virgin sandy soil using leguminous biocompost boosts Phaseolus vulgaris performance. Plants 2021, 10, 1637. [Google Scholar] [CrossRef]
- Libutti, A.; Rivelli, A.R. Quanti-qualitative response of Swiss chard (Beta vulgaris L. var. cycla) to soil amendment with biochar-compost mixtures. Agronomy 2021, 11, 307. [Google Scholar] [CrossRef]
- Libutti, A.; Trotta, V.; Rivelli, A.R. Biochar, vermicompost, and compost as soil organic amendments: Influence on Growth Parameters, Nitrate and Chlorophyll Content of Swiss Chard (Beta vulgaris L. var. cycla). Agronomy 2020, 10, 346. [Google Scholar] [CrossRef]
- Libutti, A.; Russo, D.; Lela, L.; Ponticelli, M.; Milella, L.; Rivelli, A.R. Enhancement of Yield, Phytochemical Content and Biological Activity of a Leafy Vegetable (Beta vulgaris L. var. cycla) by Using Organic Amendments as an Alternative to Chemical Fertilizer. Plants 2023, 12, 569. [Google Scholar] [CrossRef]
- Rivelli, A.R.; Libutti, A. Effect of biochar and inorganic or organic fertilizer co-application on soil properties, plant growth and nutrient content in Swiss chard. Agronomy 2022, 12, 2089. [Google Scholar] [CrossRef]
- Cheng, Y.; Bu, X.; Li, J.; Ji, Z.; Wang, C.; Xiao, X.; Li, F.; Wu, Z.-h.; Wu, G.; Jia, P. Application of biochar and compost improved soil properties and enhanced plant growth in a pb–zn mine tailings soil. Environ. Sci. Pollut. Res. 2023, 30, 32337–32347. [Google Scholar] [CrossRef]
- Huang, X.; Wang, H.; Zou, Y.; Qiao, C.; Hao, B.; Shao, Q.; Wu, W.; Wu, H.; Zhao, J.; Ren, L. Rice Straw Composting Improves the Microbial Diversity of Paddy Soils to Stimulate the Growth, Yield, and Grain Quality of Rice. Sustainability 2023, 15, 932. [Google Scholar] [CrossRef]
- Lim, S.L.; Lee, L.H.; Wu, T.Y. Sustainability of using composting and vermicomposting technologies for organic solid waste biotransformation: Recent overview, greenhouse gases emissions and economic analysis. J. Clean. Prod. 2016, 111, 262–278. [Google Scholar] [CrossRef]
- Ullah, N.; Ditta, A.; Imtiaz, M.; Li, X.; Jan, A.U.; Mehmood, S.; Rizwan, M.S.; Rizwan, M. Appraisal for organic amendments and plant growth-promoting rhizobacteria to enhance crop productivity under drought stress: A review. J. Agron. Crop Sci. 2021, 207, 783–802. [Google Scholar] [CrossRef]
- Monlau, F.; Francavilla, M.; Sambusiti, C.; Antoniou, N.; Solhy, A.; Libutti, A.; Zabaniotou, A.; Barakat, A.; Monteleone, M. Toward a functional integration of anaerobic digestion and pyrolysis for a sustainable resource management. Comparison between solid-digestate and its derived pyrochar as soil amendment. Appl. Energy 2016, 169, 652–662. [Google Scholar] [CrossRef]
- Zabaniotou, A.; Rovas, D.; Delivand, M.; Francavilla, M.; Libutti, A.; Cammerino, A.; Monteleone, M. Conceptual vision of bioenergy sector development in Mediterranean regions based on decentralized thermochemical systems. Sustain. Energy Technol. Assess. 2017, 23, 33–47. [Google Scholar] [CrossRef]
- Zabaniotou, A.; Rovas, D.; Libutti, A.; Monteleone, M. Boosting circular economy and closing the loop in agriculture: Case study of a small-scale pyrolysis–biochar based system integrated in an olive farm in symbiosis with an olive mill. Environ. Dev. 2015, 14, 22–36. [Google Scholar] [CrossRef]
- Kammann, C.I.; Linsel, S.; Gößling, J.W.; Koyro, H.-W. Influence of biochar on drought tolerance of Chenopodium quinoa Willd and on soil–plant relations. Plant Soil 2011, 345, 195–210. [Google Scholar] [CrossRef]
- Ramzani, P.M.A.; Shan, L.; Anjum, S.; Ronggui, H.; Iqbal, M.; Virk, Z.A.; Kausar, S. Improved quinoa growth, physiological response, and seed nutritional quality in three soils having different stresses by the application of acidified biochar and compost. Plant Physiol. Biochem. 2017, 116, 127–138. [Google Scholar] [CrossRef]
- Jensen, C.; Jacobsen, S.-E.; Andersen, M.; Nunez, N.; Andersen, S.; Rasmussen, L.; Mogensen, V. Leaf gas exchange and water relation characteristics of field quinoa (Chenopodium quinoa Willd.) during soil drying. Eur. J. Agron. 2000, 13, 11–25. [Google Scholar] [CrossRef]
- Razzaghi, F.; Jacobsen, S.-E.; Jensen, C.R.; Andersen, M.N. Ionic and photosynthetic homeostasis in quinoa challenged by salinity and drought–mechanisms of tolerance. Funct. Plant Biol. 2014, 42, 136–148. [Google Scholar] [CrossRef]
- Riccardi, M.; Pulvento, C.; Lavini, A.; d’Andria, R.; Jacobsen, S.E. Growth and ionic content of quinoa under saline irrigation. J. Agron. Crop Sci. 2014, 200, 246–260. [Google Scholar] [CrossRef]
- Geerts, S.; Mamani, R.S.; Garcia, M.; Raes, D. Response of quinoa (Chenopodium quinoa Willd.) to differential drought stress in the Bolivian Altiplano: Towards a deficit irrigation strategy within a water scarce region. In Proceedings of the 1st International Symposium on Land and Water Management for Sustainable Irrigated Agriculture, Adana, Turkey, 4–8 April 2006; pp. 4–8. [Google Scholar]
- Roy, R.; Núñez-Delgado, A.; Wang, J.; Kader, M.A.; Sarker, T.; Hasan, A.K.; Dindaroglu, T. Cattle manure compost and biochar supplementation improve growth of Onobrychis viciifolia in coal-mined spoils under water stress conditions. Environ. Res. 2022, 205, 112440. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Amonette, J.E.; Flury, M. Effect of biochar and biochar particle size on plant-available water of sand, silt loam, and clay soil. Soil Tillage Res. 2021, 212, 104992. [Google Scholar] [CrossRef]
- Italian Official Gazette Ministerial Decree. Approval of “Official Methods of Chemical Soil Analysis”. Ordinary Supplement 185 to the Official Journal of the Italian Republic 248, 21 October 1999, Roma, Italy. 1999. Available online: https://www.gazzettaufficiale.it/atto/serie_generale/caricaDettaglioAtto/originario?atto.dataPubblicazioneGazzetta=1992-05-25&atto.codiceRedazionale=092A2322&elenco30giorni=false (accessed on 22 April 2023).
- European Biochar Certificate (EBC). Comparison of European Biochar Certificate Version 4. 8 and IBI Biochar Standards Version 2.0 European Biochar Certificate First Publication March 2012b. Available online: https://www.european-biochar.org/en/home (accessed on 22 April 2023).
- The International Biochar Initiative. Standardized Product Definition and Product Testing Guidelines for Biochar That Is Used in Soil. Int. Biochar Initiat. 2015. Available online: https://biochar-international.org/standard-certification-training/biochar-standards/ (accessed on 22 April 2023).
- Jacobsen, S.-E.; Stølen, O. Quinoa-morphology, phenology and prospects for its production as a new crop in Europe. Eur. J. Agron. 1993, 2, 19–29. [Google Scholar] [CrossRef]
- Talebnejad, R.; Sepaskhah, A. Effect of different saline groundwater depths and irrigation water salinities on yield and water use of quinoa in lysimeter. Agric. Water Manag. 2015, 148, 177–188. [Google Scholar] [CrossRef]
- Smith, J.; Lüttge, U. Day-night changes in leaf water relations associated with the rhythm of crassulacean acid metabolism in Kalanchoë daigremontiana. Planta 1985, 163, 272–282. [Google Scholar] [CrossRef]
- Ghorbani, M.; Neugschwandtner, R.W.; Konvalina, P.; Asadi, H.; Kopecký, M.; Amirahmadi, E. Comparative effects of biochar and compost applications on water holding capacity and crop yield of rice under evaporation stress: A two-years field study. Paddy Water Environ. 2023, 21, 47–58. [Google Scholar] [CrossRef]
- Busch, D.; Kammann, C.; Grünhage, L.; Müller, C. Simple biotoxicity tests for evaluation of carbonaceous soil additives: Establishment and reproducibility of four test procedures. J. Environ. Qual. 2012, 41, 1023–1032. [Google Scholar] [CrossRef]
- Koger, C.H.; Reddy, K.N.; Poston, D.H. Factors affecting seed germination, seedling emergence, and survival of texasweed (Caperonia palustris). Weed Sci. 2004, 52, 989–995. [Google Scholar] [CrossRef]
- Videgain-Marco, M.; Marco-Montori, P.; Martí-Dalmau, C.; Jaizme-Vega, M.d.C.; Manyà-Cervelló, J.J.; García-Ramos, F.J. Effects of biochar application in a sorghum crop under greenhouse conditions: Growth parameters and physicochemical fertility. Agronomy 2020, 10, 104. [Google Scholar] [CrossRef]
- Bu, X.; Xue, J.; Wu, Y.; Ma, W. Effect of biochar on seed germination and seedling growth of Robinia pseudoacacia L. in karst calcareous soils. Commun. Soil Sci. Plant Anal. 2020, 51, 352–363. [Google Scholar] [CrossRef]
- Azuara, M.; Sáiz, E.; Manso, J.A.; García-Ramos, F.J.; Manyà, J.J. Study on the effects of using a carbon dioxide atmosphere on the properties of vine shoots-derived biochar. J. Anal. Appl. Pyrolysis 2017, 124, 719–725. [Google Scholar] [CrossRef]
- Pituello, C.; Francioso, O.; Simonetti, G.; Pisi, A.; Torreggiani, A.; Berti, A.; Morari, F. Characterization of chemical–physical, structural and morphological properties of biochars from biowastes produced at different temperatures. J. Soils Sediments 2015, 15, 792–804. [Google Scholar] [CrossRef]
- Demiraj, E.; Libutti, A.; Malltezi, J.; Rroço, E.; Brahushi, F.; Monteleone, M.; Sulçe, S. Effect of organic amendments on nitrate leaching mitigation in a sandy loam soil of Shkodra district, Albania. Ital. J. Agron. 2018, 13, 93–102. [Google Scholar] [CrossRef]
- Libutti, A.; Mucci, M.; Francavilla, M.; Monteleone, M. Effect of biochar amendment on nitrate retention in a silty clay loam soil. Ital. J. Agron. 2016, 11, 273–276. [Google Scholar] [CrossRef]
- Ventura, M.; Sorrenti, G.; Panzacchi, P.; George, E.; Tonon, G. Biochar reduces short-term nitrate leaching from a horizon in an apple orchard. J. Environ. Qual. 2013, 42, 76–82. [Google Scholar] [CrossRef]
- Wang, Y.; Pan, F.; Wang, G.; Zhang, G.; Wang, Y.; Chen, X.; Mao, Z. Effects of biochar on photosynthesis and antioxidative system of Malus hupehensis Rehd. seedlings under replant conditions. Sci. Hortic. 2014, 175, 9–15. [Google Scholar] [CrossRef]
- Akhtar, S.S.; Li, G.; Andersen, M.N.; Liu, F. Biochar enhances yield and quality of tomato under reduced irrigation. Agric. Water Manag. 2014, 138, 37–44. [Google Scholar] [CrossRef]
- Haider, I.; Raza, M.A.S.; Iqbal, R.; Aslam, M.U.; Habib-ur-Rahman, M.; Raja, S.; Khan, M.T.; Aslam, M.M.; Waqas, M.; Ahmad, S. Potential effects of biochar application on mitigating the drought stress implications on wheat (Triticum aestivum L.) under various growth stages. J. Saudi Chem. Soc. 2020, 24, 974–981. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, F.; Bendevis, M.; Shabala, S.; Jacobsen, S.E. Sensitivity of two quinoa (Chenopodium quinoa Willd.) varieties to progressive drought stress. J. Agron. Crop Sci. 2014, 200, 12–23. [Google Scholar] [CrossRef]
- Andersen, S.; Rasmussen, L.; Jensen, C.; Mogensen, V.; Andersen, M.; Jacobsen, S. Leaf water relations and gas exchange of field grown Chenopodium quinoa Willd. during drought. In Proceedings of the Small Grain Cereals and Pseudocereals; Workshop at KVL: Copenhagen, Denmark, 1996. [Google Scholar]
- Kammann, C.I.; Schmidt, H.-P.; Messerschmidt, N.; Linsel, S.; Steffens, D.; Müller, C.; Koyro, H.-W.; Conte, P.; Joseph, S. Plant growth improvement mediated by nitrate capture in co-composted biochar. Sci. Rep. 2015, 5, 11080. [Google Scholar]
- Khan, Z.; Khan, M.N.; Zhang, K.; Luo, T.; Zhu, K.; Hu, L. The application of biochar alleviated the adverse effects of drought on the growth, physiology, yield and quality of rapeseed through regulation of soil status and nutrients availability. Ind. Crops Prod. 2021, 171, 113878. [Google Scholar] [CrossRef]
- Zhang, A.; Bian, R.; Pan, G.; Cui, L.; Hussain, Q.; Li, L.; Zheng, J.; Zheng, J.; Zhang, X.; Han, X. Effects of biochar amendment on soil quality, crop yield and greenhouse gas emission in a Chinese rice paddy: A field study of 2 consecutive rice growing cycles. Field Crops Res. 2012, 127, 153–160. [Google Scholar] [CrossRef]
- Foster, E.J.; Hansen, N.; Wallenstein, M.; Cotrufo, M.F. Biochar and manure amendments impact soil nutrients and microbial enzymatic activities in a semi-arid irrigated maize cropping system. Agric. Ecosyst. Environ. 2016, 233, 404–414. [Google Scholar] [CrossRef]
- Mannan, M.; Halder, E.; Karim, M.; Ahmed, J. Alleviation of adverse effect of drought stress on soybean (Glycine max. L.) by using poultry litter biochar. Bangladesh Agron. J. 2016, 19, 61–69. [Google Scholar] [CrossRef]
- Agbna, G.H.; Dongli, S.; Zhipeng, L.; Elshaikh, N.A.; Guangcheng, S.; Timm, L.C. Effects of deficit irrigation and biochar addition on the growth, yield, and quality of tomato. Sci. Hortic. 2017, 222, 90–101. [Google Scholar] [CrossRef]
- Telahigue, D.C.; Yahia, L.B.; Aljane, F.; Belhouchett, K.; Toumi, L. Grain yield, biomass productivity and water use efficiency in quinoa (Chenopodium quinoa Willd.) under drought stress. J. Sci. Agric. 2017, 1, 222–232. [Google Scholar] [CrossRef]
- Aslam, M.U.; Raza, M.A.S.; Saleem, M.F.; Waqas, M.; Iqbal, R.; Ahmad, S.; Haider, I. Improving strategic growth stage-based drought tolerance in quinoa by rhizobacterial inoculation. Commun. Soil Sci. Plant Anal. 2020, 51, 853–868. [Google Scholar] [CrossRef]
- Tayyab, M.; Islam, W.; Khalil, F.; Ziqin, P.; Caifang, Z.; Arafat, Y.; Hui, L.; Rizwan, M.; Ahmad, K.; Waheed, S. Biochar: An efficient way to manage low water availability in plants. Appl. Ecol. Environ. Res. 2018, 16, 2565–2583. [Google Scholar] [CrossRef]
- de Melo Carvalho, M.; de Holanda Nunes Maia, A.; Madari, B.; Bastiaans, L.; Van Oort, P.; Heinemann, A.; Soler da Silva, M.; Petter, F.; Marimon, B., Jr.; Meinke, H. Biochar increases plant-available water in a sandy loam soil under an aerobic rice crop system. Solid Earth 2014, 5, 939–952. [Google Scholar] [CrossRef]
- Nawaz, M.F.; Bourrie, G.; Trolard, F. Soil compaction impact and modelling. A review. Agron. Sustain. Dev. 2013, 33, 291–309. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Property | Unit | Value |
|---|---|---|
| Sand | (%) | 66.1 |
| Silt | (%) | 11.5 |
| Clay | (%) | 22.4 |
| Field Capacity (FC) at −0.03 MPa | (% dw) | 22.8 |
| Permanent Wilting Point (PWP) at −1.5 MPa | (% dw) | 11.4 |
| pH | (-) | 7.6 |
| EC | (dS m−1) | 0.6 |
| Corg | (g kg−1) | 5.9 |
| Organic Matter | (%) | 1.0 |
| Ntotal | (‰) | 1.5 |
| C/N | (-) | 3.9 |
| Property | Unit | Biochar | Vermicompost | |
|---|---|---|---|---|
| Woodchips | Vineyard Prunings | |||
| pH | (-) | 8.9 ± 0.1 | 10.6 ± 0.1 | 7.6 ± 0.1 |
| EC | (mS m−1) | 52.0 ± 0.0 | 249.0 ± 0.0 | 265.0 ± 0.0 |
| Moisture | (% dw) | 5.6 ± 0.1 | 15.3 ± 0.3 | 4.0 ± 0.2 |
| Volatile solids | (% dw) | 42.3 ± 0.4 | 15.3 ± 0.3 | 27.5 ± 0.6 |
| Ash | (% dw) | 4.4 ± 0.2 | 9.9 ± 0.0 | 72.2 ± 0.6 |
| Fixed carbon | (% dw) | 53.3 ± 0.2 | 74.8 ± 0.3 | 0.2 ± 0.0 |
| C | (% dw) | 68.3 ± 0.1 | 67.7 ± 0.9 | 11.3 ± 0.0 |
| H | (% dw) | 4.0 ± 0.0 | 2.1 ± 0.0 | 1.5 ± 0.1 |
| N | (% dw) | 1.0 ± 0.0 | 1.0 ± 0.0 | 1.5 ± 0.0 |
| S | (% dw) | 0.03 ± 0.0 | 0.2 ± 0.0 | 0.3 ± 0.0 |
| Corg | (% dw) | 66.3 ± 0.1 | 67.0 ± 0.9 | 7.8 ± 0.1 |
| O | (% dw) | 22.3 ± 0.3 | 17.9 ± 1.5 | 5.2 ± 0.2 |
| H/Corg | (-) | 0.7 ± 0.0 | 0.4 ± 0.0 | - |
| O/Corg | (-) | 0.4 ± 0.0 | 0.2 ± 0.0 | - |
| C/N | (-) | 67.2 ± 2.0 | 66.2 ± 0.1 | 5.0 ± 0.2 |
| Treatment | Phenological Stage (Days) | |||||
|---|---|---|---|---|---|---|
| Seedling Emergence | Leaf Development Stages | Bud Initiation | Flowering Initiation | |||
| Two-Leaf | From Six-Leaf | To Twelve-Leaf | ||||
| Cw | 4.3 C | 8.7 C | 19.7 | 26.3 | 28.7 | 41.3 |
| Cs | 4.3 C | 10.7 A | 21.0 | 25.3 | 28.6 | 42.7 |
| B1 | 4.7 BC | 11.0 A | 19.7 | 25.0 | 28.3 | 40.7 |
| B2 | 5.7 B | 9.3 BC | 21.3 | 26.3 | 29.0 | 43.0 |
| V | 4.7 BC | 8.3 C | 21.3 | 25.3 | 28.3 | 42.3 |
| B1+V | 3.7 C | 9.3 BC | 22.3 | 26.3 | 29.3 | 44.3 |
| B2+V | 7.0 A | 10.0 AB | 21.0 | 26.0 | 28.0 | 41.0 |
| Significance | *** | ** | ns | ns | ns | ns |
| Treatment | Absolute Growth Rate | ||
|---|---|---|---|
| AGR1 | AGR2 | AGR3 | |
| Cw | 1.33 a | 2.04 A | 2.62 B |
| Cs | 1.38 a | 1.19 C | 2.40 B |
| B1 | 1.41 a | 1.80 AB | 2.65 B |
| B2 | 1.24 a | 0.89 C | 2.03 C |
| V | 1.14 a | 1.63 B | 2.45 B |
| B1+V | 1.29 a | 1.78 AB | 2.53 B |
| B2+V | 0.72 b | 1.17 C | 2.98 A |
| Significance | * | *** | *** |
| Treatment | End of Water-Stress Period | After Soil Re-Watering | |||||||
|---|---|---|---|---|---|---|---|---|---|
| RWC | Ψ | Ψπ | TW/DW | RWC | Ψ | Ψπ | Ψp | TW/DW | |
| Cw | 75.4 A | −1.70 A | −1.81 A | 11.3 | 77.5 A | −1.37 A | −1.53 A | 0.17 | 10.5 C |
| Cs | 56.1 D | −2.50 B | −2.49 BC | 11.6 | 69.1 BC | −1.68 BC | −1.94 BC | 0.26 | 11.2 BC |
| B1 | 58.5 CD | −2.58 B | −2.51 BC | 10.2 | 70.7 AC | −1.71 BC | −1.88 BC | 0.17 | 10.8 C |
| B2 | 64.7 BC | −2.07 AB | −2.05 AB | 11.1 | 73.2 AB | −1.50 AB | −1.72 AB | 0.22 | 12.9 AB |
| V | 59.5 CD | −2.53 B | −2.54 C | 10.1 | 70.0 BC | −1.82 C | −1.91 BC | 0.09 | 11.1 BC |
| B1+V | 55.5 D | −2.56 B | −2.58 C | 8.1 | 66.5 C | −1.77 BC | −2.01 C | 0.25 | 10.2 C |
| B2+V | 69.6 AB | −1.73 A | −1.76 A | 10.3 | 72.2 AB | −1.52 AB | −1.68 AB | 0.17 | 13.1 A |
| Significance | ** | ** | ** | ns | * | * | * | ns | * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akram, M.Z.; Libutti, A.; Rivelli, A.R. Evaluation of Vegetative Development of Quinoa under Water Stress by Applying Different Organic Amendments. Agronomy 2023, 13, 1412. https://doi.org/10.3390/agronomy13051412
Akram MZ, Libutti A, Rivelli AR. Evaluation of Vegetative Development of Quinoa under Water Stress by Applying Different Organic Amendments. Agronomy. 2023; 13(5):1412. https://doi.org/10.3390/agronomy13051412
Chicago/Turabian StyleAkram, Muhammad Zubair, Angela Libutti, and Anna Rita Rivelli. 2023. "Evaluation of Vegetative Development of Quinoa under Water Stress by Applying Different Organic Amendments" Agronomy 13, no. 5: 1412. https://doi.org/10.3390/agronomy13051412