
The Inhibitory Mechanism of Eugenol on Lasiodiplodia theobromae and Its Induced Disease Resistance of Passion Fruit

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of Pathogenic Fungi
2.1.1. Isolation and Identification of Pathogenic Fungi and Detection of Pathogenicity
2.1.2. Morphological Identification of Pathogenic Fungi
2.1.3. ITS Sequence Analysis of Pathogenic Fungi
2.2. Effect of Eugenol on the Mycelial Growth of the Passion Fruit Rot Pathogen
2.3. Effect of Different Concentrations of Eugenol on Spore Germination of the Passion Fruit Rot Pathogen
2.4. Effect of Eugenol on Cell Membrane Integrity of the Passion Fruit Rot Pathogen
2.4.1. Determination of Relative Conductivity, Nucleic Acid Release, Intracellular Protein Release, and Soluble Sugar Release by Fruit Rot Pathogens
2.4.2. Propidium Iodide (PI) Staining to Observe the Effect of Eugenol on the DNA of Passion-Fruit-Rot-Causing Fungi
2.5. Inoculation Treatment for Induction of Eugenol-Dependent Passion Fruit Rot Resistance
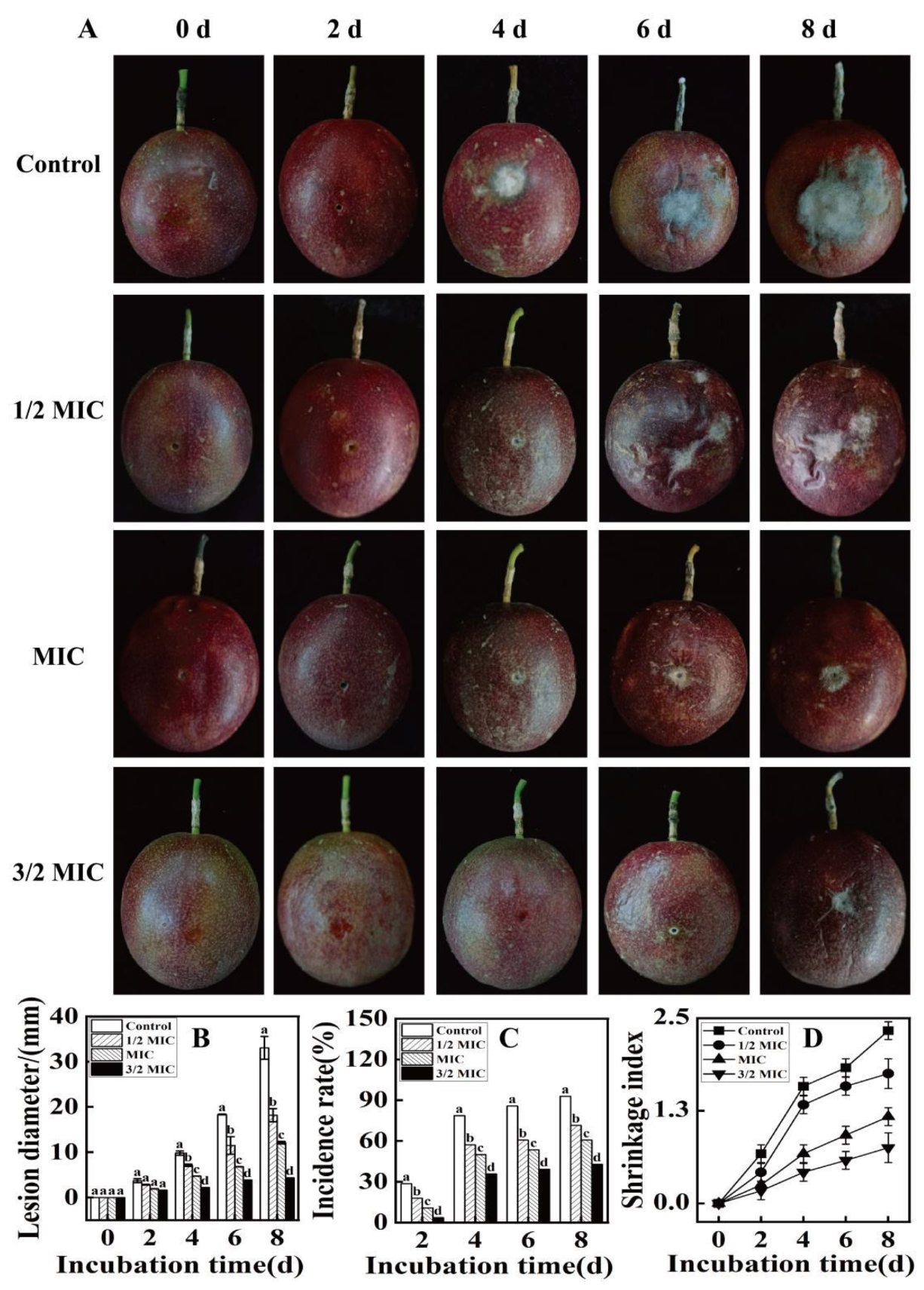
Effect of Eugenol on the Incidence of Fruit Rot and Spot Diameter in Passion Fruit
2.6. Effect of Eugenol Treatment on the Shrivelling of Passion Fruit
2.7. Effect of Eugenol Treatment on Defence Enzyme Activity in Passion Fruit
2.8. Effect of Eugenol Treatment on the Total Phenolic and Flavonoid Content of Passion Fruit
2.9. Statistical Analysis of Data
3. Results
3.1. Isolation, Identification, and Pathogenicity Detection of Pathogenic Fungi
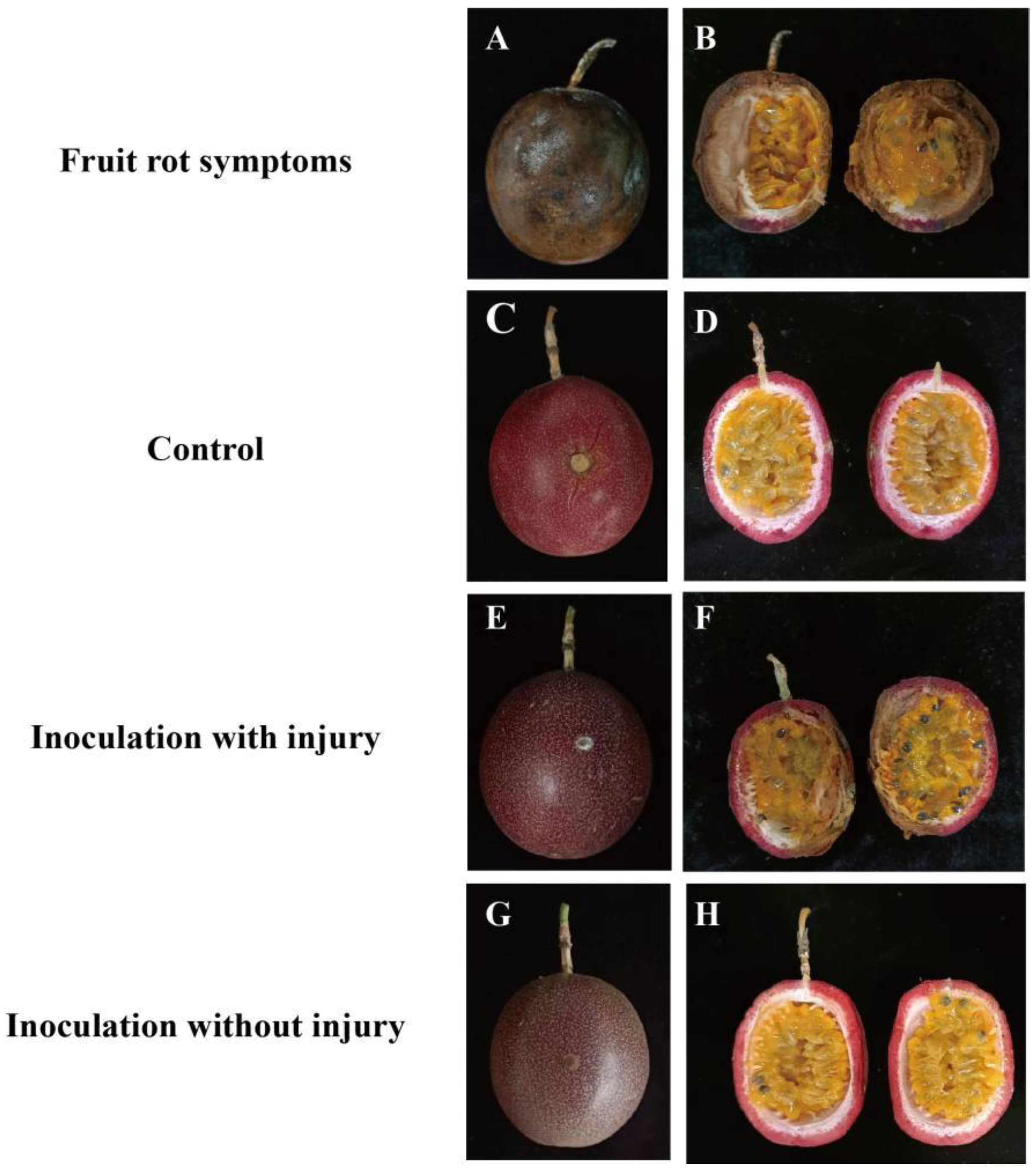
3.1.1. Symptoms of Naturally Occurring Passion Fruit Rot Disease
3.1.2. Pathogenicity of the Passion Fruit Rot Pathogen
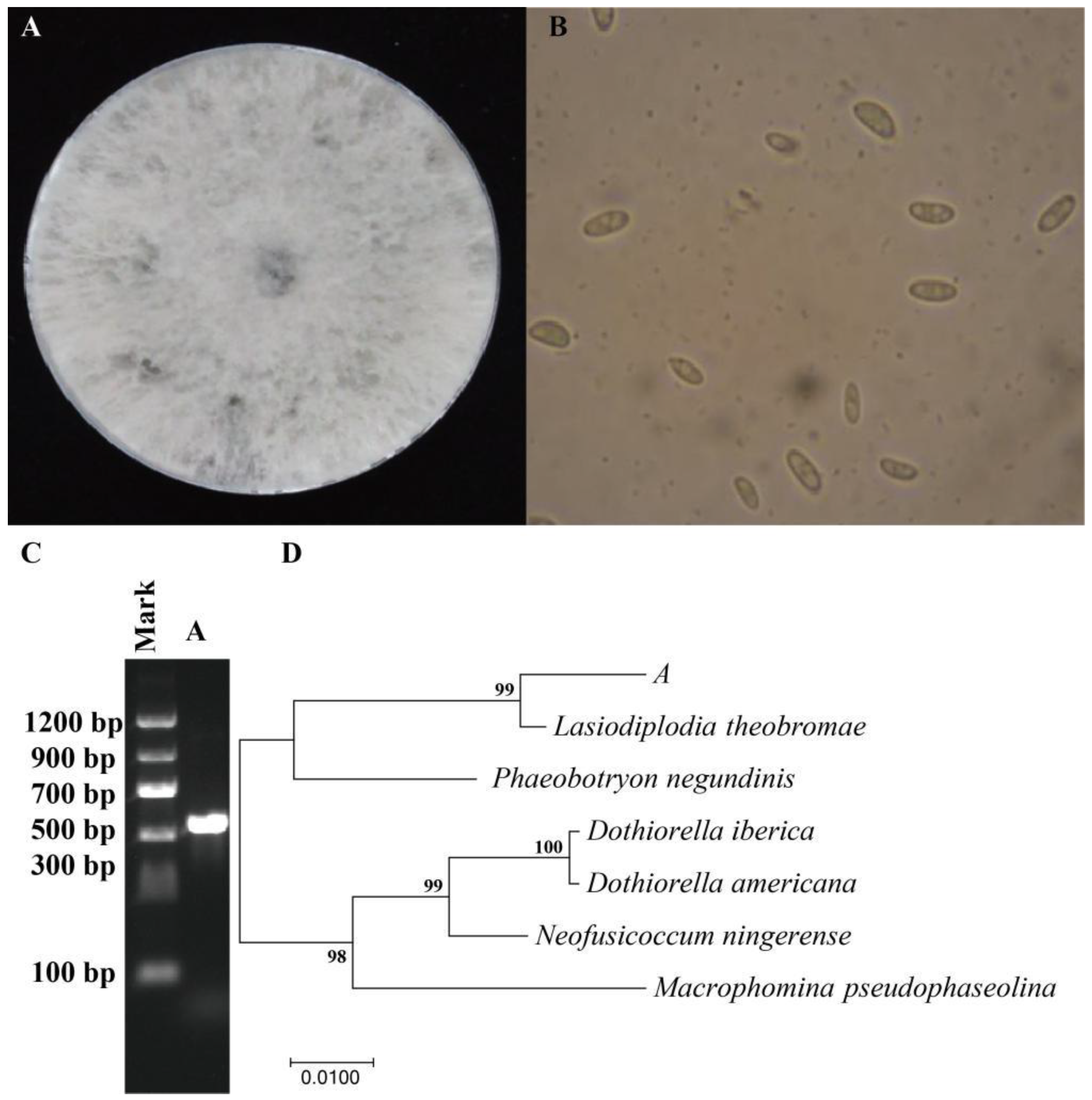
3.1.3. Identification of Pathogenic Fungi of Passion Fruit Rot Disease
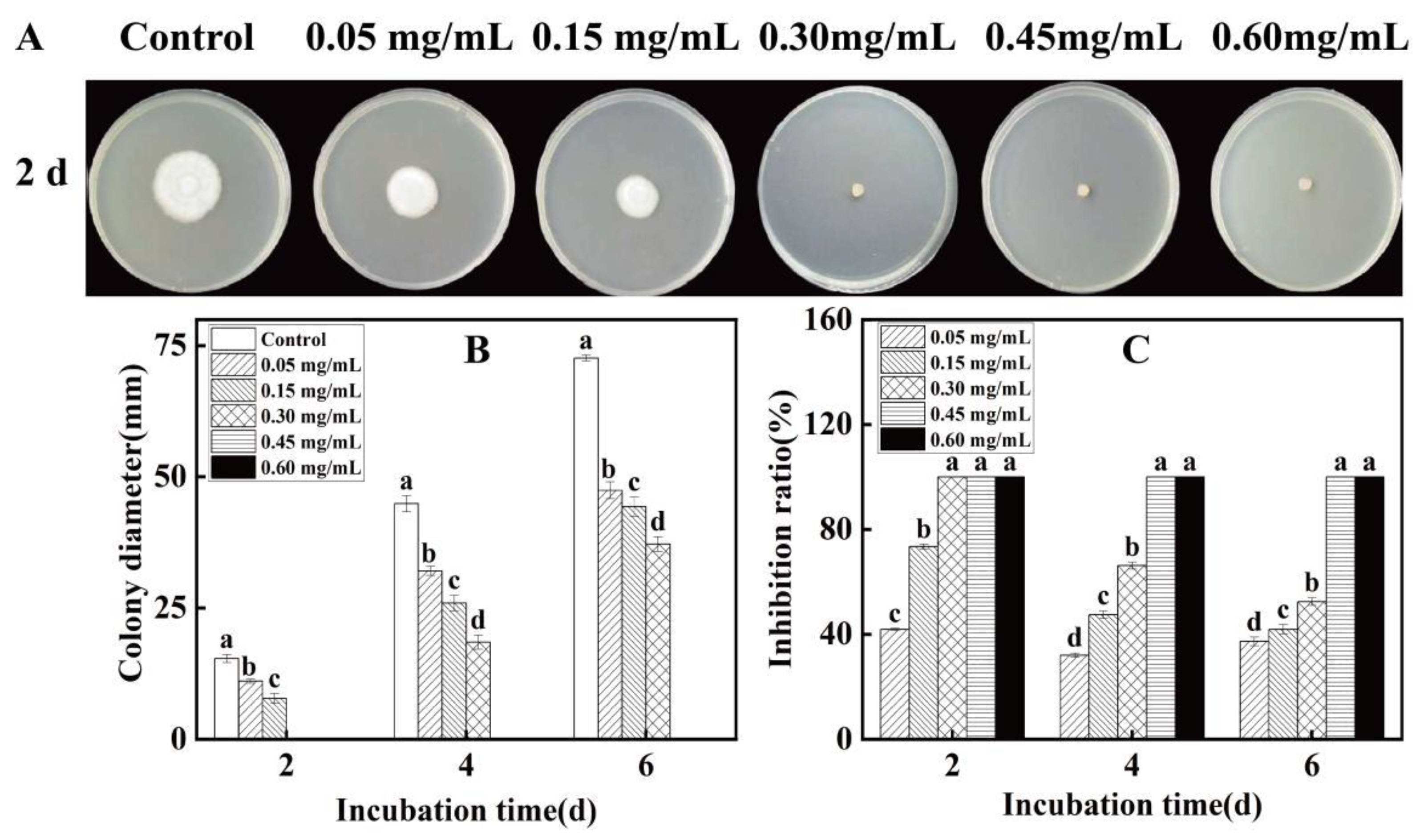
3.2. Inhibitory Effect of Eugenol on the Passion Fruit Rot Pathogen
3.3. Effect of Eugenol on the Cell Membrane of the Passion Fruit Rot Pathogen
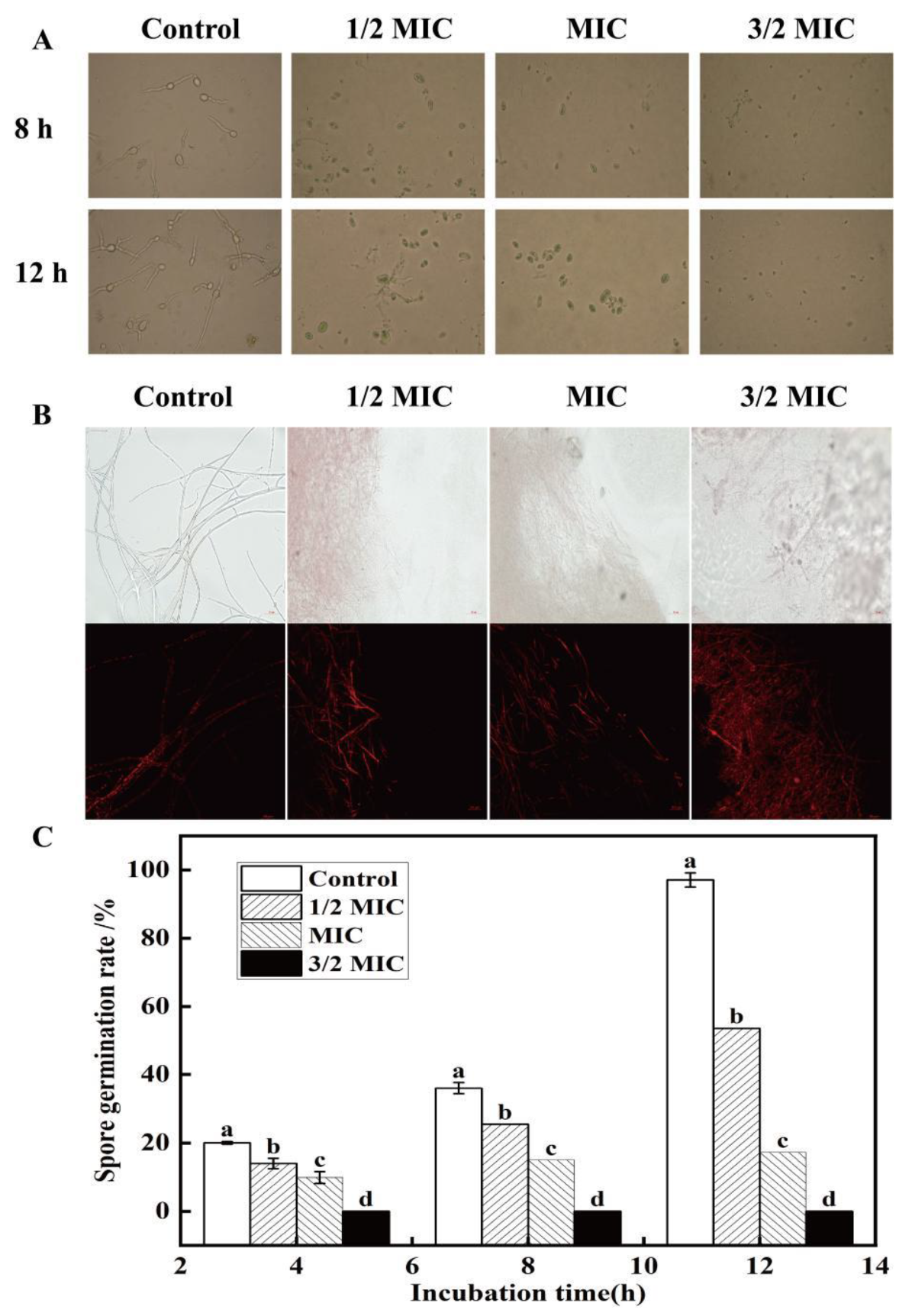
3.3.1. Effect of Eugenol on Spore Germination and Cell Membrane of the Passion Fruit Rot Pathogen
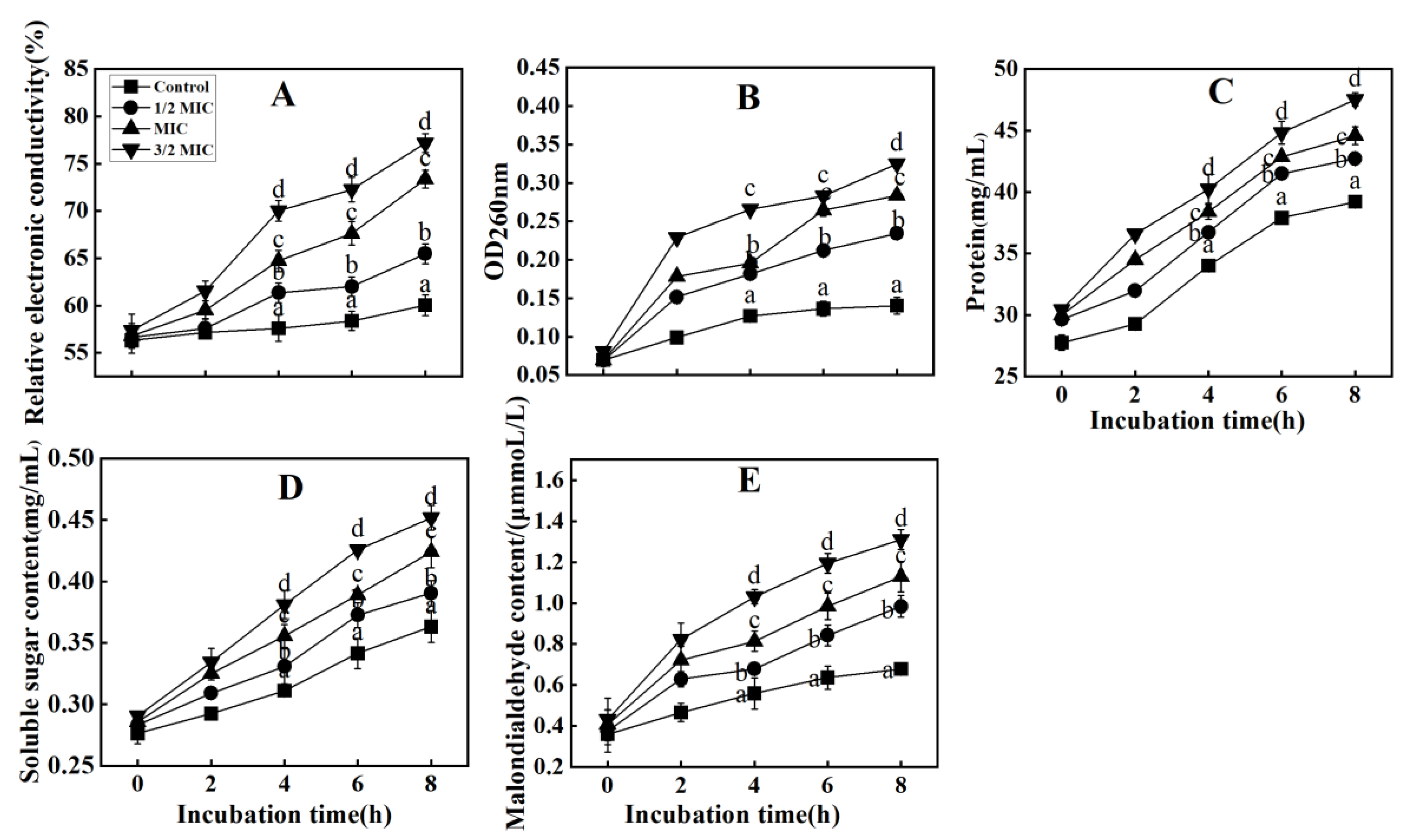
3.3.2. Effect of Eugenol on Mycelial Nucleic Acid Release and Electrical Conductivity of the Mycorrhizal Fungi That Causes Fruit Rot in Passion Fruit
3.3.3. Effects of Eugenol on Intracellular Protein Release, Soluble Sugar Release, and Malondialdehyde (MDA) Content in the Pathogenic Passion Fruit Rot Fungi
3.4. Mechanism of Eugenol-Dependent Resistance to Passion Fruit Rot after Harvest
3.4.1. Effect of Different Eugenol Concentrations on the Resistance of Passion Fruit to Fruit Rot
3.4.2. Effect of Eugenol on Disease-Resistant Substances and Defence-Related Enzyme Activities in Passion Fruit
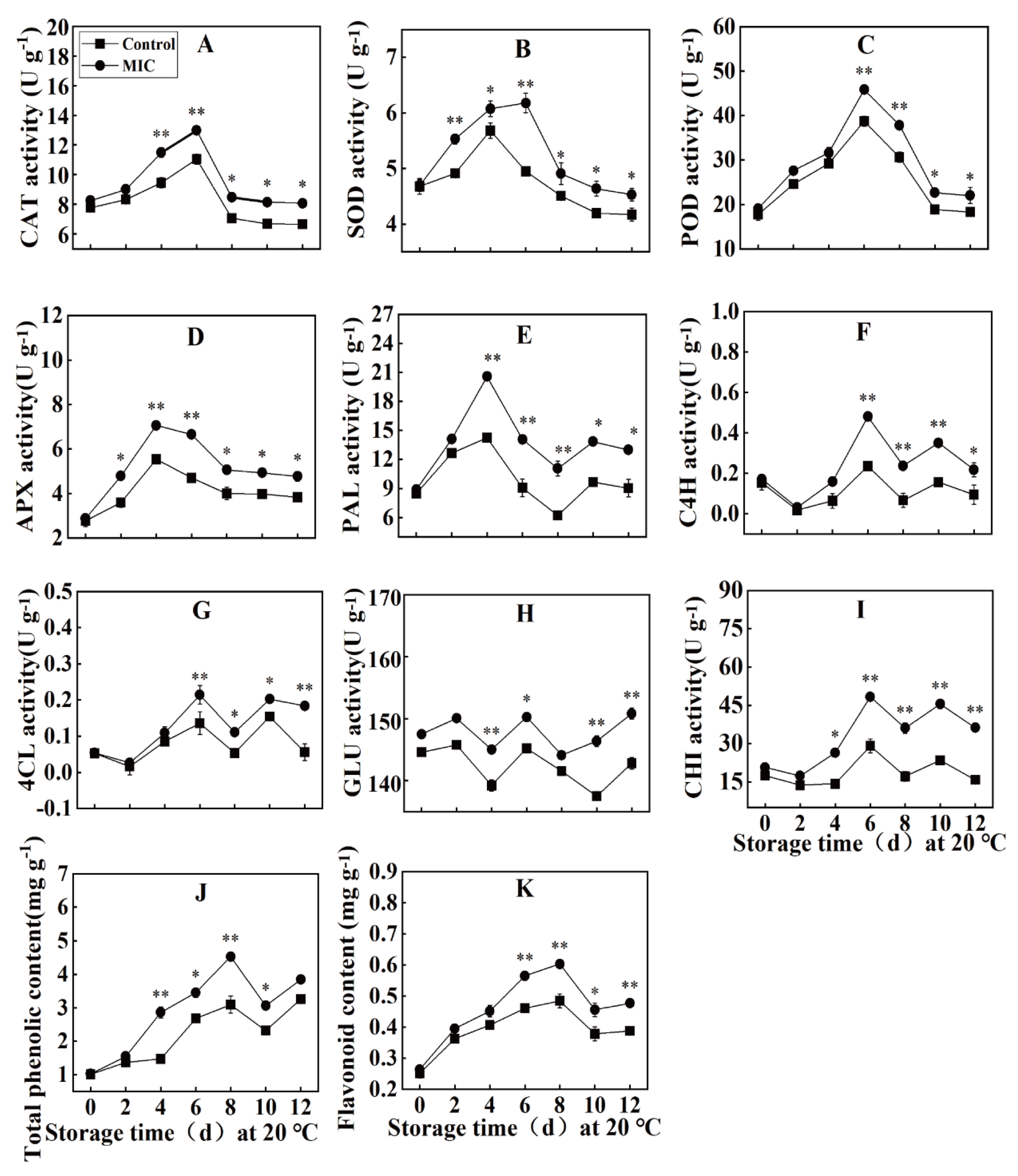
Effect of Eugenol on CAT, SOD, POD, and APX Activities in Passion Fruit
Effect of Eugenol on the Activity of PAL, C4H, and 4CL in Passion Fruit
Effect of Eugenol on the GLU and CHI Activity in Passion Fruit
Effect of Eugenol on the Total Phenolic and Flavonoid Content of Passion Fruit
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Correia, A.D.O.; Alexandre, R.S.; Pfenning, L.H.; Cabanez, P.A.; Ferreira, A.; Ferreira, M.; de Lima, P.A.M.; de Mello, M.T.; Otoni, W.C.; Lopes, J.C. Passiflora mucronata, a passion fruit wild species resistant to fusariosis and a potential rootstock for commercial varieties. Sci. Hortic. 2022, 302, 111174. [Google Scholar] [CrossRef]
- De Oliveira, A.B.; de Almeida, L.; Mônica Maria, D.A.L.; Carlos, F.H.M.; Luciana, D.S.O.; Kellina, O.D.S.; Enéas, G.F.; Laurent, U.; Maria, R.A.D.M. Effects of organic and conventional farming systems on quality and antioxidant metabolism of passion fruit during maturation. Sci. Hortic. 2017, 222, 84–89. [Google Scholar] [CrossRef]
- Jordán, M.J.; Goodner, K.L.; Shaw, P.E. Characterization of the aromatic profile in aqueous essence and fruit juice of yellow passion fruit (Passiflora edulis sims F. Flavicarpa degner) by GC-MS and GC/O. J. Agric. Food Chem. 2002, 50, 1523–1528. [Google Scholar] [CrossRef] [PubMed]
- Porto-Figueira, P.; Freitas, A.; Cruz, C.J.; Figueira, J.; Câmara, J.S. Profiling of passion fruit volatiles: An effective tool to discriminate between species and varieties. Food Res. Int. 2015, 77, 408–418. [Google Scholar]
- Alemwati, P.; Sagar, V.; Ram, K.P.; Ram, A.; Ram, R.S.; Sanjay, K.S. Physiological and quality changes during postharvest ripeningof purple passion fruit (Passiflora edulis Sims). Fruits 2014, 69, 19–30. [Google Scholar]
- Mcevoy, C.T.; Wallace, I.R.; Hamill, L.L.; Hunter, S.J.; Neville, C.E.; Patterson, C.; Woodside, J.V.; Young, I.S.; McKinley, M.C. Increasing fruit and vegetable intake has no dose-response effect on conventional cardiovascular risk factors in overweight adults at high risk of developing cardiovascular disease. J. Nutr. 2015, 145, 1464–1471. [Google Scholar] [CrossRef]
- Sun, Y.B.; Jiang, C.Q.; Kar, K.C.; Wei, S.Z.; Gabriel, M.; Tai, H.L.; Schooling, C.M. Fruit and vegetable consumption and cardiovascular risk factors in older Chinese: The guangzhou biobank cohort study. PLoS ONE 2015, 10, e0135380. [Google Scholar] [CrossRef]
- Araujo, L.S.; Rodrigues, C.E.M.; Soares, T.L.; Idália, S.D.; Jesus, O.N.D. Effect of time and storage conditions on the physical and physico-chemical characteristics of the pulp of yellow and purple passion fruit. Food Sci. Technol. 2017, 37, 500–506. [Google Scholar] [CrossRef]
- Shiomi, S.; Kubo, Y.; Wamocho, L.S.; Koaze, H.; Nakamura, R.; Inaba, A. Postharvest ripening and ethylene biosynthesis in purple passion fruit. Postharvest Biol. Technol. 1996, 8, 188–207. [Google Scholar] [CrossRef]
- Du, Y.X.; Shi, N.N.; Chen, W.L.; Ruan, H.C.; Yang, X.J.; Gan, L. Identification of colletotrichum brevisporum causing anthracnose on passion fruit. Can. J. Plant Pathol. 2017, 39, 1367725. [Google Scholar] [CrossRef]
- Dai, Y.L.; Wang, C.C.; Lin, H.L. First report of Septoria Blotch of passion fruit caused by Septoria passifloricola in Taiwan. Plant Dis. 2021, 105, 700. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Niu, X.L.; Yang, J.Y. First report of postharvest fruit rot on passion fruit (Passiflora edulis) caused by Lasiodiplodia theobromae in China’s mainland. Plant Dis. 2020, 105, 1198. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Zhu, X.J.; Cao, P.; Shaomin, W.; Lu, Y.H. Antibacterial and antibiofilm activities of eugenol from essential oil of Syzygium aromaticum (L.) Merr. & L.M. Perry (clove) leaf against periodontal pathogen Porphyromonas gingivalis. Microb. Pathog. 2017, 113, 396–402. [Google Scholar] [PubMed]
- Jafri, H.; Mohd, S.A.K.; Iqbal, A. In vitro efficacy of eugenol in inhibiting single and mixed-biofilms of drug-resistant strains of Candida albicans and Streptococcus mutans. Phytomedicine 2018, 54, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Devi, K.P.; Nisha, S.A.; Sakthivel, R.; Pandian, S.K. Eugenol (an essential oil of clove) acts as an antibacterial agent against Salmonella typhi by disrupting the cellular membrane. J. Ethnopharmacol. 2010, 1301, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.Y.; Zhang, C.H.; Li, C.Z.; Lin, L. Antimicrobial mechanism of clove oil on Listeria monocytogenes. Food Control 2018, 94, 140–146. [Google Scholar] [CrossRef]
- Edwards, K.; Johnstone, C.; Thompson, C. A simple and rapid method for the preparation of plant genomic DNA for PCR analysis. Nucleic Acids Res. 1991, 19, 1349. [Google Scholar] [CrossRef]
- Yahyazadeh, M.; Omidbaigi, R.; Zare, R.; Taheri, H. Effect of some essential oils on mycelial growth of Penicillium digitatum Sacc. World J. Microbiol. Biotechnol. 2008, 24, 1445–1450. [Google Scholar] [CrossRef]
- Ge, Y.H.; Chen, Y.R.; Li, C.Y.; Wei, M.L.; Lv, J.Y. Inhibitory effects of sodium silicate on the fungal growth and secretion of cell wall-degrading enzymes by Trichothecium roseum. J. Phytopathol. 2017, 165, 620–625. [Google Scholar] [CrossRef]
- Liu, K.; Zhou, X.; Fu, M. Inhibiting effects of ε-poly-lysine (ε-PL) on Pencillium digitatum and its involved mechanism. Postharvest Biol. Technol. 2017, 123, 94–101. [Google Scholar] [CrossRef]
- Tao, N.G.; Yang, Q.L.; Jia, L. Citral inhibits mycelial growth of Penicillium italicum by a membrane damage mechanism. Food Control 2014, 41, 116–121. [Google Scholar] [CrossRef]
- Chen, C.Y.; Qi, W.W.; Peng, X.; Chen, J.Y.; Wan, C.P. Inhibitory Effect of 7-Demethoxytylophorine on Penicillium italicum and its Possible Mechanism. Microorganisms 2019, 7, 36. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Ge, Y.H.; Li, C.Y.; Chen, Y.R.; Wang, W.H.; Duan, B.; Li, X. Antifungal activity of ε-poly-L-lysine on Trichothecium roseum in vitro and its mechanisms. Physiol. Mol. Plant Pathol. 2018, 103, 23–27. [Google Scholar] [CrossRef]
- Qin, G.Z.; Zong, Y.Y.; Chen, Q.L.; Hua, D.L.; Tian, S.P. Inhibitory effect of boron against Botrytis cinerea on table grapes and its possible mechanisms of action. Int. J. Food Microbiol. 2010, 138, 45–150. [Google Scholar] [CrossRef]
- Chen, H.Z.; Cheng, Z.; Wisniewski, M.; Liu, Y.S.; Liu, J. Ecofriendly hot water treatment reduces postharvest decay and elicits defense response in kiwifruit. Environ. Sci. Pollut. Res. 2015, 22, 15037–15045. [Google Scholar] [CrossRef]
- Xia, Y.X.; Chen, T.; Qin, G.Z.; Li, B.Q.; Tian, S.P. Synergistic action of antioxidative systems contributes to the alleviation of senescence in kiwifruit. Postharvest Biol. Technol. 2016, 111, 15–24. [Google Scholar] [CrossRef]
- Zhang, M.; Xu, L.Y.; Zhang, L.Y.; Zhang, L.; Guo, Y.H.; Qi, X.; He, L. Effects of quercetin on postharvest blue mold control in kiwifruit. Sci. Hortic. 2018, 228, 18–25. [Google Scholar] [CrossRef]
- Zheng, F.L.; Zheng, W.W.; Li, L.M.; Pan, S.M.; Liu, M.C.; Zhang, W.W.; Liu, H.S.; Zhu, C.Y. Chitosan Controls Postharvest Decay and Elicits Defense Response in Kiwifruit. Food Bioprocess Technol. 2017, 10, 1937–1945. [Google Scholar] [CrossRef]
- Zhang, W.L.; Jiang, H.T.; Cao, J.K.; Jiang, W.B. UV-C treatment controls brown rot in postharvest nectarine by regulating ROS metabolism and anthocyanin synthesis. Postharvest Biol. Technol. 2021, 180, 111613. [Google Scholar] [CrossRef]
- Li, S.G.; Xu, Y.H.; Bi, Y.; Zhang, B.; Shen, S.L.; Jiang, T.J.; Zheng, X.L. Melatonin treatment inhibits gray mold and induces disease resistance in cherry tomato fruit during postharvest. Postharvest Biol. Technol. 2019, 157, 110962. [Google Scholar] [CrossRef]
- Li, G.J.; Zhu, S.H.; Wu, W.X.; Zhang, C.; Peng, Y.; Wang, Q.G.; Shi, J.Y. Exogenous nitric oxide induces disease resistance against Monilinia fructicola through activating the phenylpropanoid pathway in peach fruit. J. Sci. Food Agric. 2017, 97, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Bautista-Rosales, P.U.; Calderon-Santoyo, M.; Servín-Villegas, R.; Ochoa-Alvarez, N.A.; Vazquez-Juarez, R.; Ragazzo-Sanchez, J.A. Biocontrol action mechanisms of Cryptococcus laurentii on Colletotrichum gloeosporioides of mango. J. Crop Protection 2014, 65, 194–201. [Google Scholar] [CrossRef]
- Boller, T.; Gehri, A.; Mauch, F.; Vögeli, U. Chitinase in bean leaves: Induction by ethylene, purification, properties, and possible function. Planta 1983, 157, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Berger, L.R.; Reynolds, D.M. The chitinase system of a strain of Streptomyces griseus. Biochim. Biophys. Acta (BBA)—Lipids Lipid Metab. 1958, 29, 522–534. [Google Scholar]
- Ramful, D.; Tarnus, E.; Aruoma, O.I.; Bourdon, E.; Bahorun, T. Polyphenol composition, vitamin C content and antioxidant capacity of Mauritian citrus fruit pulps. Food Res. Int. 2011, 44, 2088–2099. [Google Scholar] [CrossRef]
- Wang, H.; Chen, Y.H.; Lin, H.T.; Lin, M.S.; Chen, Y.H.; Lin, Y.F. 1-Methylcyclopropene containing-papers suppress the disassembly of cell wall polysaccharides in anxi persimmon fruit during storage. Int. J. Biol. Macromol. 2020, 151, 723–729. [Google Scholar] [CrossRef]
- Mirdehghan, S.H.; Valero, D. Bioactive compounds in tomato fruit and its antioxidant activity as affected by incorporation of Aloe, eugenol, and thymol in fruit package during storage. Int. J. Food Prop. 2017, 20, 1798–1806. [Google Scholar]
- Yang, R.P.; Miao, J.Y.; Shen, Y.T.; Cai, N.; Wan, C.; Zou, L.Q.; Chen, C.Y.; Chen, J.Y. Antifungal effect of cinnamaldehyde, eugenol and carvacrol nanoemulsion against Penicillium digitatum and application in postharvest preservation of citrus fruit. LWT 2021, 141, 110924. [Google Scholar] [CrossRef]
- Ju, J.; Lei, Y.; Guo, Y.H.; Yu, H.; Cheng, Y.L.; Yao, W.R. Eugenol and citral kills Aspergillus niger through the tricarboxylic acid cycle and its application in food preservation. LWT 2023, 173, 114226. [Google Scholar] [CrossRef]
- Hu, Q.; Zhou, M.F.; Wei, S.Y. Progress on the antimicrobial activity research of clove oil and eugenol in the food antisepsis field. J. Food Sci. 2018, 83, 1476–1483. [Google Scholar] [CrossRef]
- Ju, J.; Xie, Y.F.; Yu, H.; Guo, Y.H.; Cheng, Y.L.; Zhang, R.R.; Yao, W.R. Synergistic inhibition effect of citral and eugenol against Aspergillus niger and their application in bread preservation. Food Chem. 2020, 310, 125974. [Google Scholar] [CrossRef]
- Li, W.S.; Yuan, S.Z.; Sun, J.; Li, Q.; Jiang, W.; Cao, J. Ethyl p-coumarate exerts antifungal activity in vitroand in vivo against fruit Alternaria alternata via membrane-targeted mechanism. Int. J. Food Microbiol. 2018, 278, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.X.; Fan, X.J.; Cai, X.Q.; Hu, F.P. The inhibitory mechanisms by mixtures of two endophytic bacterial strains isolated from Ginkgo biloba against pepper phytophthora blight. Biol. Control 2015, 85, 59–67. [Google Scholar] [CrossRef]
- Shu, C.; Zhao, H.D.; Jiao, W.X.; Liu, B.D.; Cao, J.K.; Jiang, W.B. Antifungal efficacy of ursolic acid in control of Alternaria alternata causing black spot rot on apple fruit and possible mechanisms involved. Sci. Hortic. 2019, 256, 108636. [Google Scholar] [CrossRef]
- Sun, W.J.; Lv, W.J.; Li, L.N.; Yin, G.; Hang, X.F.; Xue, Y.F.; Chen, J.; Shi, Z.Q. Eugenol confers resistance to Tomato yellow leaf curl virus (TYLCV) by regulating the expression of SlPer1 in tomato plants. New Biotechenology 2016, 33, 345–354. [Google Scholar] [CrossRef]
- Kumar, P.; Dashyal, M.S.; Doddaraju, P.; Meti, B.S.; Girigowda, M. Differential gene responses in different varieties of pomegranate during the pathogenesis of Xanthomonas axonopodis pv. punicae. 3 Biotech 2021, 11, 180. [Google Scholar] [CrossRef]
- Yang, R.P.; Miao, J.Y.; Chen, X.; Chen, C.Y.; Simal-Gandara, J.; Chen, J.Y.; Wan, C.P. Essential oils nano-emulsion confers resistance against Penicillium digitatum in ‘Newhall’ navel orange by promoting phenylpropanoid metabolism. Ind. Crops Prod. 2022, 187, 115297. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Shuai, L.; Luo, D.; Ba, L. The Inhibitory Mechanism of Eugenol on Lasiodiplodia theobromae and Its Induced Disease Resistance of Passion Fruit. Agronomy 2023, 13, 1408. https://doi.org/10.3390/agronomy13051408
Sun Y, Shuai L, Luo D, Ba L. The Inhibitory Mechanism of Eugenol on Lasiodiplodia theobromae and Its Induced Disease Resistance of Passion Fruit. Agronomy. 2023; 13(5):1408. https://doi.org/10.3390/agronomy13051408
Chicago/Turabian StyleSun, Yanzheng, Liang Shuai, Donglan Luo, and Liangjie Ba. 2023. "The Inhibitory Mechanism of Eugenol on Lasiodiplodia theobromae and Its Induced Disease Resistance of Passion Fruit" Agronomy 13, no. 5: 1408. https://doi.org/10.3390/agronomy13051408