
Compensatory Structural Growth Responses of Early-Succession Native Warm-Season Grass Stands to Defoliation Management

Abstract
:1. Introduction
2. Materials and Methods
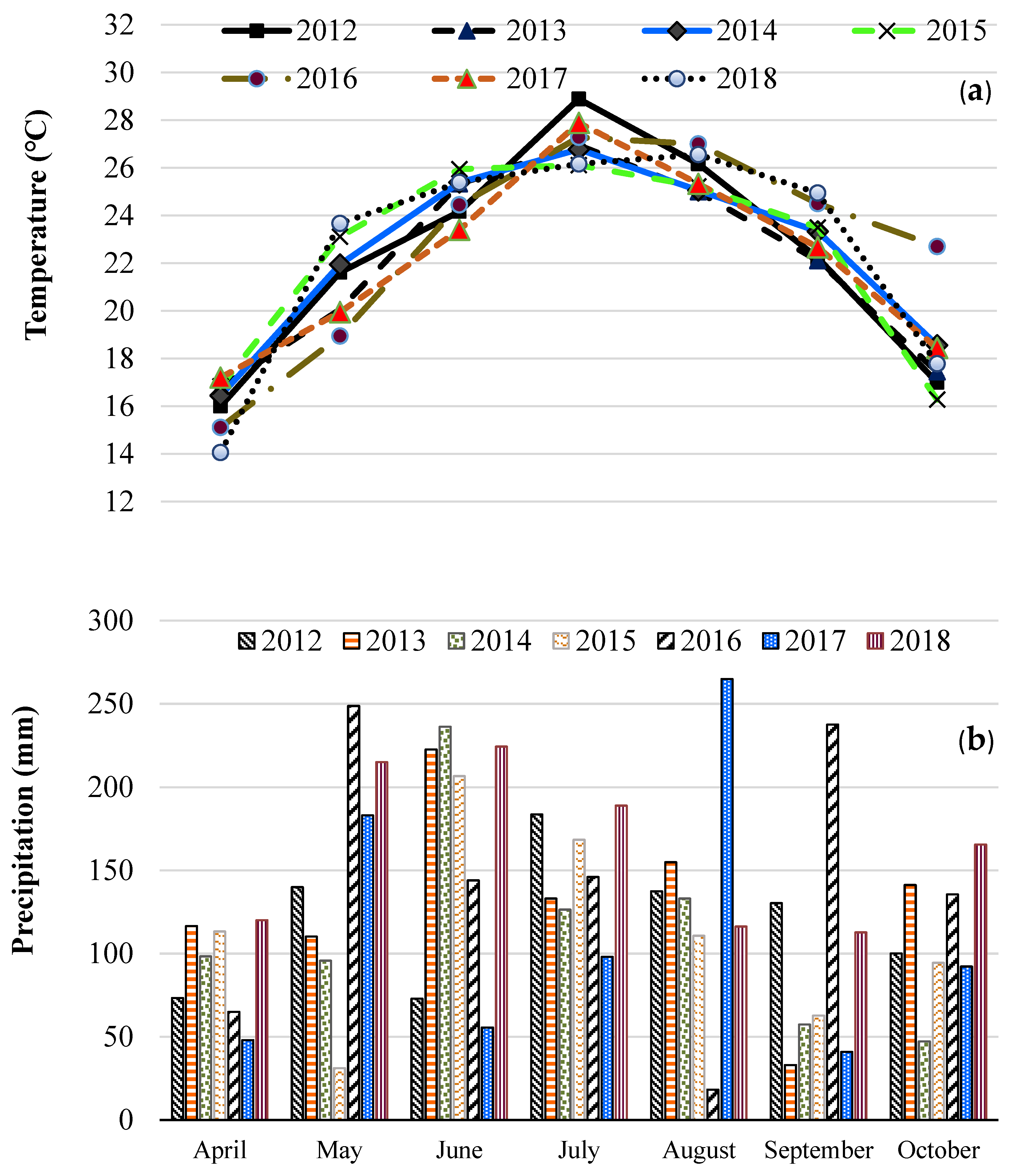
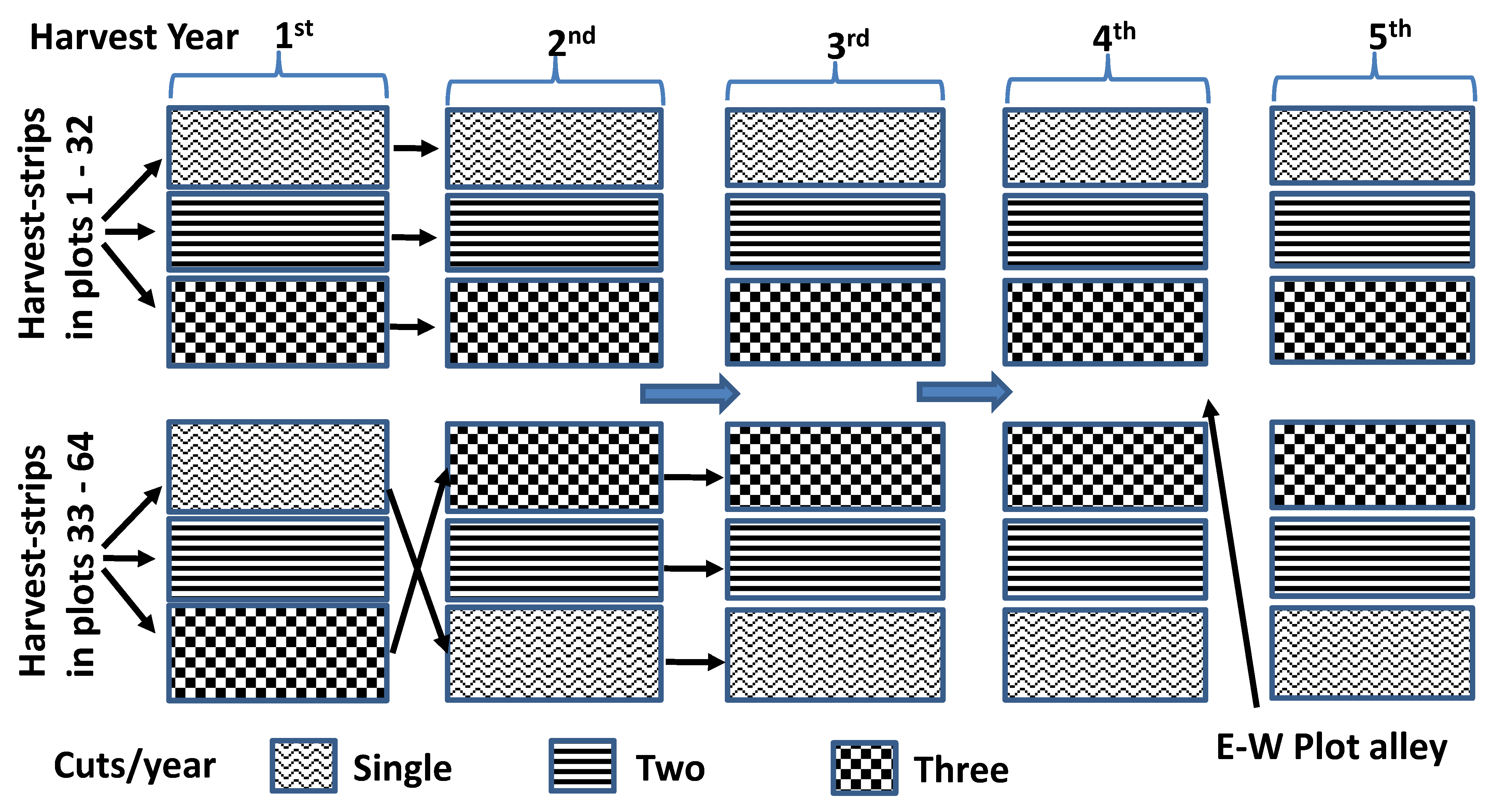
2.1. Location and Field Preparations
2.2. Vegetation Measurements
2.3. Data Analysis
3. Results and Discussion
3.1. Early-Summer Sward-Heights
3.2. Late-Fall Sward-Heights
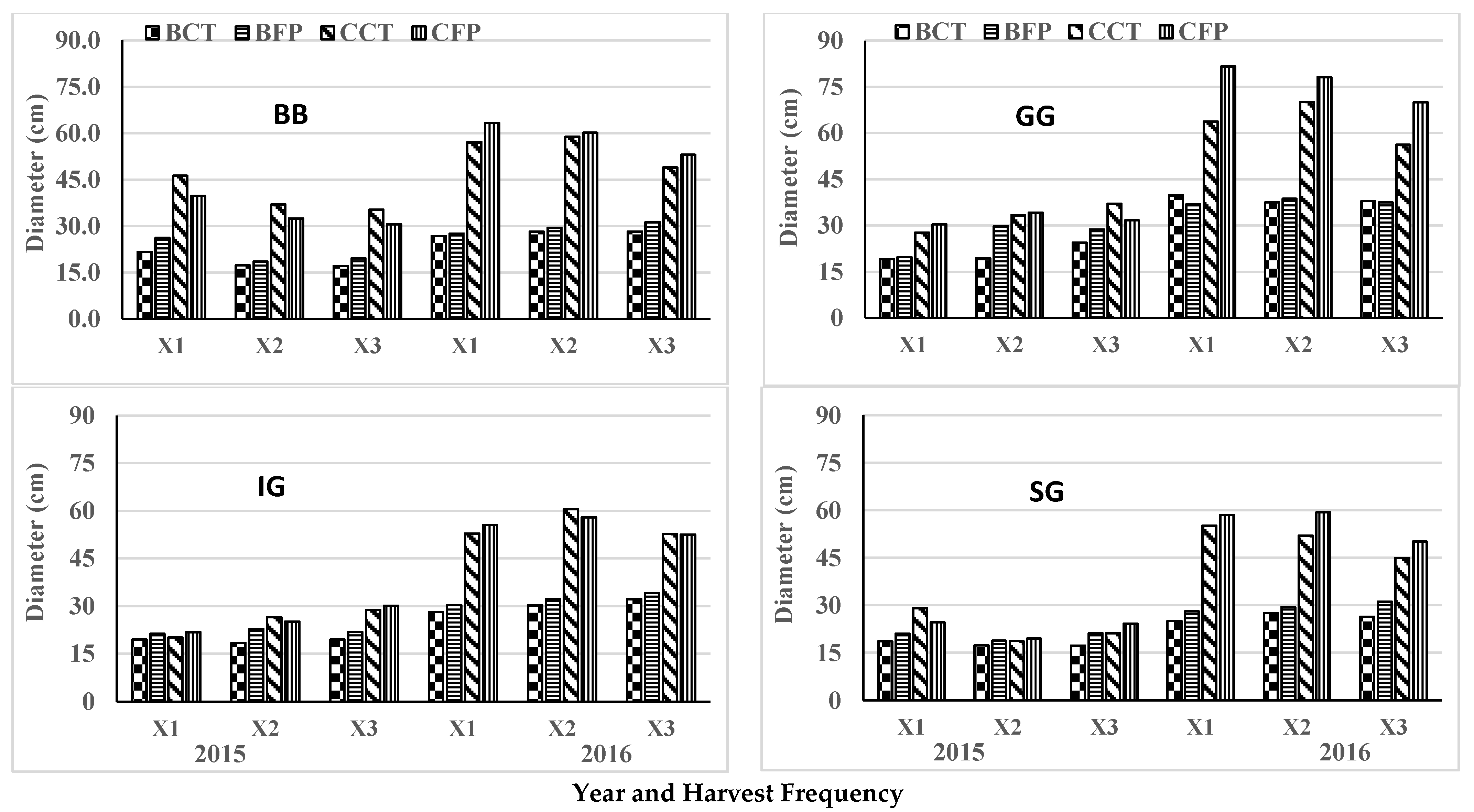
3.3. Basal Diameter
3.4. Canopy Diameter
3.5. Canopy-to-Basal Diameter Ratios
3.6. Sward Structure
3.6.1. Canopy Light Interception
3.6.2. Stand Density
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Temu, V.W.; Rude, B.J.; Baldwin, B.S. Yield response of native warm-season forage grasses to harvest intervals and durations in mixed stands. Agronomy 2014, 4, 90–107. [Google Scholar] [CrossRef]
- Caswell, H.; Reed, F.; Stephenson, S.N.; Werner, P.A. Photosynthetic pathways and selective herbivory: A hypothesis. Am. Nat. 1973, 107, 465–480. [Google Scholar] [CrossRef]
- Ferraro, D.O.; Oesterheld, M. Effect of defoliation on grass growth. a quantitative review. Oikos 2002, 98, 125–133. [Google Scholar] [CrossRef]
- Manske, L.L.; Ske, L.L. General Description of Grass Growth and Development and Defoliation Resistance Mechanisms; Range Management Report DREC 98-1022; NDSU Dickinson Research Extension Center: Dickinson, ND, USA, 1998. [Google Scholar]
- Garay, A.H. Tiller size/density compensation in perennial ryegrass miniature swards subject to differing defoliation heights and a proposed productivity index. Grass Forage Sci. 1999, 54, 347–356. [Google Scholar] [CrossRef]
- Slepetys, J.; Šterne, D. The productivity and persistency of pure and mixed forage legume swards. Latv. J. Agron. 2008, 11, 276–281. [Google Scholar]
- Harris, W. Pasture as an Ecosystem. In Pastures, Their Ecology and Management; Langer, R.H.M., Ed.; Oxford University Press: Oakdale, CA, USA, 1990; pp. 75–131. [Google Scholar]
- Lee, W.G.; Fenner, M.; Loughnan, A.; Lloyd, K.M. Long-term effects of defoliation: Incomplete recovery of a New Zealand alpine tussock grass, Chionochloa pallens, after 20 years. J. Appl. Ecol. 2000, 37, 348–355. [Google Scholar] [CrossRef]
- Briske, D.D.; Richards, J.H. Plant Responses to Defoliation: A Physiological, Morphological and Demographic Evaluation. Wildland Plants: Physiological Ecology and Developmental Morphology; Society for Range Manage: Denver, CO, USA, 1995; pp. 635–710. [Google Scholar]
- Dahl, B.E. Developmental Morphology of Plants. In Wildland Plants: Physiological Ecology and Developmental Morphology, 1st ed.; Bedunah, D.J., Sosebee, R.E., Eds.; Society for Range Manage: Denver, CO, USA, 1995; pp. 22–58. [Google Scholar]
- Noy-Meir, I. Compensating growth of grazed plants and its relevance to the use of rangelands. Ecol. Appl. 1993, 3, 32–34. [Google Scholar] [CrossRef]
- Anderson, V.J.; Briske, D.D. Herbivore-induced species replacement in grasslands: Is it driven by herbivory tolerance or avoidance? Ecol. Appl. 1995, 5, 1014–1024. [Google Scholar] [CrossRef]
- Cullen, B.R.; Chapman, D.F.; Quigley, P.E. Comparative defoliation tolerance of temperate perennial grasses. Grass Forage Sci. 2006, 61, 405–412. [Google Scholar] [CrossRef]
- Loeser, M.R.; Crews, T.E.; Sisk, T.D. Defoliation increased above-ground productivity in a semi-arid grassland. J. Range Manag. 2004, 57, 442–447. [Google Scholar] [CrossRef]
- Smith, S.E. Variation in response to defoliation between populations of Bouteloua curtipendula var. caespitosa (Poaceae) with different livestock grazing histories. Am.J. Bot. 1998, 85, 1266–1272. [Google Scholar] [CrossRef]
- Mullahey, J.J.; Waller, S.S.; Moser, L.E. Defoliation effects on production and morphological development of little bluestem. J. Range Manag. 1990, 43, 497–500. [Google Scholar] [CrossRef]
- Turner, C.L.; Seastedt, T.R.; Dyer, M.I. Maximization of aboveground grassland production: The role of defoliation frequency, intensity, and history. Ecol. Applic. 1993, 3, 175–186. [Google Scholar] [CrossRef]
- Woodis, J.E.; Jackson, R.D. The effects of clipping height and frequency on net primary production of Andropogon gerardii (C4 grass) and Bromus inermis (C3 grass) in greenhouse experiments. Grass Forage Sci. 2008, 63, 458–466. [Google Scholar] [CrossRef]
- Nelson, C.J.; Moser, L.E. Plant Factors Affecting Forage Quality. In Forage Quality, Evaluation, and Utilization; ASA-CSSA-SSSA: Madison, WI, USA, 1994; pp. 115–154. [Google Scholar]
- Campbell, G.S.; Norman, J.M. The Description and Measurement of Plant Canopy Structure. In Plant Canopies: Their Growth, Form and Function; Russell, G., Marshall, B., Jarvis, P.G., Eds.; Cambridge University Press: Cambridge, UK, 1989; pp. 1–20. [Google Scholar]
- D’angelo, G.H.; Postulka, E.B.; Ferrari, L. Infrequent and intense defoliation benefits dry-matter accumulation and persistence of clipped Arrhenatherum elatius. Grass Forage Sci. 2005, 60, 17–24. [Google Scholar] [CrossRef]
- Hodgson, J. Grazing Management. Science Into Practice; Longman Group UK Ltd.: Harlow, UK, 1990; p. 203. [Google Scholar]
- Difante, G.D.S.; Nascimento Júnior, D.D.; Euclides, V.P.B.; Silva, S.C.D.; Barbosa, R.A.; Gonçalves, W.V. Sward structure and nutritive value of tanzania guineagrass subjected to rotational stocking managements. Rev. Bras. De Zootec. 2009, 38, 9–19. [Google Scholar] [CrossRef]
- Norman, J.M.; Campbell, G.S. Canopy Structure. In Plant Physiological Ecology; Springer: Dordrecht, The Netherlands, 1989; pp. 301–325. [Google Scholar]
- Vermeire, L.T.; Gillen, R.L. Estimating herbage standing crop with visual obstruction in tallgrass prairie. J. Range Manag. 2001, 54, 57–60. [Google Scholar] [CrossRef]
- Benkobi, L.; Uresk, D.W.; Schenbeck, G.; King, R.M. Protocol for monitoring standing crop in grasslands using visual obstruction. Rangel. Ecol. Manag. J. Range Manag. Arch. 2000, 53, 627–633. [Google Scholar]
- Gonzalez, A.; Loreau, M. The causes and consequences of compensatory dynamics in ecological communities. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 393–414. [Google Scholar] [CrossRef]
- Lusk, J.J.; Smith, S.G.; Fuhlendorf, S.D.; Guthery, F.S. Factors influencing northern bobwhite nest-site selection and fate. J. Wildl. Manag. 2006, 70, 564–571. [Google Scholar] [CrossRef]
- Jones, J.; Coggin, D.S.; Cummins, J.L.; Hill, J. Restoring and Managing Native Prairies. In A Handbook for Mississippi Landowners; Wildlife Mississippi: Starkville, MS, USA, 2007. [Google Scholar]
- Temu, V.W.; Rutto, L.K.; Kering, M.K. Compensatory Yield Responses of Young Native Warm-Season Grass Stands to Seasonal Changes in Harvest Frequencies. Agronomy 2022, 12, 2761. [Google Scholar] [CrossRef]
- NOAA Satellite and Information Service. Drought Termination and Amelioration; National Climatic Data Center: Asheville, NC, USA, 2017.
- Pedreira, C.G.; Sollenberger, L.E.; Mislevy, P. Botanical composition, light interception, and carbohydrate reserve status of grazed ‘Florakirk’ Bermudagrass. Agron. J. 2000, 92, 194–199. [Google Scholar]
- Briske, D.D. Developmental Morphology and Physiology f Grasses. In Grazing Management: An Ecological Perspective; Heitschmidt, R.K., Stuth, J.W., Eds.; Timber Press: Portland, OR, USA, 1991; pp. 85–108. [Google Scholar]
- Belesky, D.P.; Fedders, J.M. Defoliation effects on seasonal production and growth rate of cool-season grasses. Agron. J. 1994, 86, 38–45. [Google Scholar] [CrossRef]
- Garcia, F.; Carrère, P.; Soussana, J.F.; Baumont, R. How do severity and frequency of grazing affect sward characteristics and the choices of sheep during the grazing season? Grass Forage Sci. 2003, 58, 138–150. [Google Scholar] [CrossRef]
- Rodolfo, G.R.; Schmitt, D.; Dias, K.M.; Sbrissia, A.F. Levels of defoliation and regrowth dynamics in elephant grass swards. Ciência Rural. 2015, 45, 1299–1304. [Google Scholar] [CrossRef]
- Parsons, A.J.; Leafe, E.L.; Collett, B.; Stiles, W. The physiology of grass production under grazing. I. Characteristics of leaf and canopy photosynthesis of continuously-grazed swards. J. Appl. Ecol. 1983, 20, 117–126. Available online: https://www.jstor.org/stable/2403380 (accessed on 27 March 2023). [CrossRef]
- Mullahey, J.J.; Waller, S.S.; Moser, L.E. Defoliation effects on yield and bud and tiller numbers of two Sandhills grasses. J. Range Manag. 1991, 44, 241–245. [Google Scholar] [CrossRef]
- Buwai, M.; Trlica, M.J. Multiple defoliation effects on herbage yield, vigor, and total nonstructural carbohydrates of five range species. J. Range Manag. 1977, 30, 164–171. [Google Scholar] [CrossRef]
- Richards, J.H.; Mueller, J.H.; Mott, J.J. Tillering in tussock grasses in relation to defoliation and apical bud removaL. Ann. Bot. 1988, 62, 173–179. [Google Scholar] [CrossRef]
- Murphy, J.S.; Briske, D.D. Regulation of tillering by apical dominance: Chronology, interpretive value, and current perspectives. J. Range Manage. 1992, 45, 419–429. [Google Scholar] [CrossRef]
- Holland, E.A.; Detling, J.K. Plant response to herbivory and belowground nitrogen cycling. Ecology 1990, 71, 1040–1049. [Google Scholar] [CrossRef]
- Benot, M.-L.; Morvan-Bertrand, A.; Mony, C.; Huet, J.; Sulmon, C.; Decau, M.-L.; Prud’homme, M.-P.; Bonis, A. Grazing intensity modulates carbohydrate storage pattern in five grass species from temperate grasslands. Acta Oecologica 2019, 95, 108–115. [Google Scholar] [CrossRef]
- Ash, A.J.; Corfield, J.P.; Mclvor, J.G.; Ksiksi, T.S. Grazing management in tropical savannas: Utilization and rest strategies to manipulate rangeland condition. Rangel. Ecol. Manag. 2011, 64, 223–239. [Google Scholar] [CrossRef]
- Eckert, R.E.; Spencer, J.S. Growth and reproduction of grasses heavily grazed under rest-rotation management. Rangel. Ecol. Manag. J. Range Manag. Arch. 1987, 40, 156–159. [Google Scholar] [CrossRef]
- Guevara, J.C.; Stasi, C.R.; Estevez, O.R. Effect of cattle grazing on range perennial grasses in the Mendoza plain, Argentina. J. Arid Environ. 1996, 34, 205–213. [Google Scholar] [CrossRef]
- Davies, K.W.; Boyd, C.S.; Bates, J.D. Eighty years of grazing by cattle modifies sagebrush and bunchgrass structure. Rangel. Ecol. Manag. 2018, 71, 275–280. [Google Scholar] [CrossRef]
- Gutman, M.; Noy-Meir, I.; Pluda, D.; Seligman, N.A.; Rothman, S.; Sternberg, M. Biomass partitioning following defoliation of annual and perennial Mediterranean grasses. Conserv. Ecol. 2002, 5. Available online: http://www.consecol.org/vol5/iss2/art1 (accessed on 27 March 2023).
- Peterson, P.R.; Hutton, S.J.; Abaye, A.O.; Wolf, D.D.; Benson, G.B. Eastern gamagrass responses to defoliation management. In Proceedings of the 2nd Eastern Native Grass Symposium, Baltimore, MD, USA, 17–19 November 1999; pp. 253–258. [Google Scholar]
- Ballaré, C.L.; Scopel, A.L.; Sánchez, R.A. Photocontrol of stem elongation in plant neighbourhoods: Effects of photon fluence rate under natural conditions of radiation. Plant Cell Environ. 1991, 14, 57–65. [Google Scholar] [CrossRef]
- Frank, A.B.; Hofman, L. Light quality and stem numbers in cool-season forage grasses. Crop Sci. 1994, 34, 468–473. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Harvest Dates by Harvest Regime | ||||
|---|---|---|---|---|
| Year | Cuts | Three Cuts | Two Cuts | Single Cut |
| 2013 * | 1st | 5 June 2013 | 5 June 2013 | 5 June 2013 |
| 2nd | 18 November 2013 | 18 November 2013 | 18 November 2013 | |
| 1st (2014) | 1st | 24 June 2014 | 23 July 2014 | 14 September 2014 |
| 2nd | 29 July 2014 | 12 September 2014 | ||
| 3rd | 12 September 2014 | |||
| 2nd (2015) | 1st | 14 June 2015 | 18 June 2015 | 14 October 2015 |
| 2nd | 31 July 2015 | 14 October 2015 | ||
| 3rd | 14 October 2015 | |||
| 3rd (2016) | 1st | 18 June 2016 | 18 June 2016 | 24 October 2016 |
| 2nd | 05 August 2016 | 24October 2016 | ||
| 3rd | 24 October 2016 | |||
| 4th (2017) | 1st | 19 June 2017 | 29 June 2017 | 18 October 2017 |
| 2nd | 17 August 2017 | 18 October 2017 | ||
| 3rd | 17 October 2017 | |||
| 5th (2018) * | N/A | 26 June 2018 | 26 June 2018 | 26 June 2018 |
| Early-Season Heights | Late-Season Heights | |||||
|---|---|---|---|---|---|---|
| Source | DF | Fα | p > Fα | DF | Fα | p > Fα |
| Model | 119 | 49.63 | <0.001 | 95 | 300.75 | <0.001 |
| Year | 4 | 133.37 | <0.001 | 3 | 452.26 | <0.001 |
| System | 1 | 9.60 | 0.002 | 1 | 0.55 | 0.458 |
| Year × System | 4 | 13.37 | <0.001 | 3 | 35.95 | <0.001 |
| Species | 3 | 727.32 | <0.001 | 3 | 1562.85 | <0.001 |
| Year × Species | 12 | 98.54 | <0.001 | 9 | 24.93 | <0.001 |
| System × Species | 3 | 6.93 | 0.001 | 3 | 17.14 | <0.001 |
| Year × System × Species | 12 | 8.41 | <0.001 | 9 | 3.83 | <0.001 |
| Cuts | 2 | 551.60 | <0.001 | 2 | 8889.27 | <0.001 |
| Year × Cuts | 8 | 38.16 | <0.001 | 6 | 215.31 | <0.001 |
| System × Cuts | 2 | 13.41 | <0.001 | 2 | 26.11 | <0.001 |
| Year × System × Cuts | 8 | 2.21 | 0.025 | 6 | 1.38 | 0.218 |
| Species × Cuts | 6 | 32.83 | <0.001 | 6 | 338.78 | <0.001 |
| Year × Species × Cuts | 24 | 6.56 | <0.001 | 18 | 46.20 | <0.001 |
| System × Species × Cuts | 6 | 0.71 | 0.641 | 6 | 5.83 | <0.001 |
| Year × System × Species × Cuts | 24 | 0.49 | 0.982 | 18 | 4.27 | <0.001 |
| Error | 840 | 672 | ||||
| Corrected Total | 959 | 767 | ||||
| Source | DF | p > Fα | ||||
|---|---|---|---|---|---|---|
| PARi | VOH | BD | CD | CBDR | ||
| Model | 47 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Year | 1 | <0.001 | 0.003 | <0.001 | <0.001 | <0.001 |
| System | 3 | <0.001 | <0.001 | <0.001 | 0.001 | 0.001 |
| Year × System | 3 | <0.001 | 0.153 | 0.049 | <0.001 | <0.001 |
| Species | 2 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Year × Species | 2 | 0.160 | <0.001 | <0.001 | <0.001 | <0.001 |
| System × Species | 6 | <0.001 | <0.001 | 0.937 | 0.575 | 0.151 |
| Year × System × Species | 6 | 0.229 | 0.081 | <0.059 | 0.001 | 0.002 |
| Cuts | 1 | <0.001 | 0.607 | 0.096 | <0.001 | <0.001 |
| Year × Cuts | 1 | <0.001 | 0.107 | 0.154 | <0.001 | 0.004 |
| Manage × Cuts | 3 | <0.001 | 0.058 | 0.370 | 0.575 | 0.454 |
| Year × System × Cuts | 3 | 0.020 | 0.894 | 0.699 | 0.357 | 0.832 |
| Species × Cuts | 2 | 0.014 | 0.277 | 0.041 | <0.001 | 0.041 |
| Year × Species × Cuts | 2 | 0.204 | 0.031 | <0.001 | 0.022 | 0.374 |
| System × Species × Cuts | 6 | 0.643 | 0.923 | 0.109 | 0.229 | 0.038 |
| Year × System × Species × Cuts | 6 | 0.927 | 0.249 | 0.840 | 0.715 | 0.637 |
| Error | 336 | |||||
| Corrected Total | 383 | |||||
| Year | Cuts | Species and Harvest Regime | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Big Bluestem | Gamagrass | Indiangrass | Switchgrass | ||||||
| Same | Flipped | Same | Flipped | Same | Flipped | Same | Flipped | ||
| Early-summer Sward Heights | |||||||||
| ----------------------------------------------cm------------------------------------------------------- | |||||||||
| 2014 | Twice | 43.3 A§ | 42.4 A§ | 53.1 A | 56.2 A | 65.4 A | 63.8 A | 53.7 A | 50.8 A |
| Thrice | - | - | - | - | - | - | - | - | |
| 2015 | Once | 62.4 aA | 62.4 aA | 77.9 aA | 75.1 aA | 44.8 aA | 45.4 aA | 66.1 aA | 67.2 aA |
| Twice | 52.3 bA | 52.2 bA | 69.8 bA | 63.0 bB | 44.6 aA | 43.1 aA | 42.4 bB | 45.3 bA | |
| Thrice | 45.7 cA | 45.1 cA | 55.9 cA | 57.3 cA | 39.1 bA | 38.3 bA | 36.4 cB | 39.9 cA | |
| p > α # | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| 2016 | Once | 67.6 aA | 65.1 aA | 82.3 aA | 80.8 aA | 47.9 aA | 49.4 aA | 73.2 aA | 67.5 aB |
| Twice | 53.5 bA | 57.1 bA | 69.0 bA | 72.0 bA | 39.6 bB | 44.7 bA | 58.3 bB | 60.5 bA | |
| Thrice | 44.7 cB | 52.2 bA | 59.2 cA | 62.9 cA | 33.1 cA | 35.8 cA | 44.9 cA | 51.4 cA | |
| p > α | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| 2017 | Once | 60.1 aA | 56.6 aA | 75.4 aA | 71.2 aA | 39.2 aA | 38.0 aA | 71.2 aA | 65.4 aB |
| Twice | 46.5 bA | 45.4 bA | 68.0 bA | 66.6 aA | 34.4 bA | 35.6 aA | 60.5 bA | 61.6 bA | |
| Thrice | 41.9 bA | 40.4 cA | 59.4 cA | 59.1 bA | 36.1a bA | 37.7 aA | 45.7cB | 52.2 cA | |
| p > α | <0.001 | <0.001 | <0.001 | <0.001 | 0.027 | 0.289 | <0.001 | <0.001 | |
| 2018 | Once | 86.0 aA | 80.9 aA | 87.5 aA | 84.2 aA | 46.6 aA | 39.9 bB | 76.4 aA | 73.2 aA |
| Twice | 61.8 bA | 58.8 bA | 74.6 bA | 75.3a bA | 44.7a bA | 41.9a bA | 61.8 bA | 61.2 bA | |
| Thrice | 52.7 cA | 56.0 bA | 67.7 bA | 70.4 bA | 42.3 bA | 44.5 aA | 46.0 cA | 50.1 cA | |
| p > α | <0.001 | <0.001 | <0.001 | <0.001 | 0.022 | 0.018 | <0.001 | <0.001 | |
| Late-fall Sward Heights | |||||||||
| 2014 | Once | 171.5 aA | 159.6 aA | 70.9 aB | 96.3 aA | 197.9 aA | 173.1aB | 194.4 aA | 174.1 aB |
| Twice | 69.8 bA | 61.1 bA | 64.5 aA | 65.5 bA | 69.2 bA | 63.5 bA | 91.0 bA | 95.4 bA | |
| Thrice | 47.4 cA | 49.9 bA | 49.9 bB | 57.3 bA | 66.2 bA | 58.5 bA | 85.6 bA | 86.2 cA | |
| p > α | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| 2015 | Once | 188.9 aA | 165.8 aB | 111.6 aA | 103.9 aB | 187.2 aA | 194.2 aA | 206.1 aA | 185.0 aB |
| Twice | 165.0 bA | 145.5 bB | 82.7 bA | 87.2 bA | 157.9 bA | 160.0 bA | 152.2 bB | 158.9 bA | |
| Thrice | 30.1 cB | 40.1 cA | 45.3 cB | 50.9 cA | 96.5 cA | 90.2 cA | 57.5 cB | 66.7 cA | |
| p > α | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| 2016 | Once | 183.7 aA | 182.6 aA | 104.0 aB | 118.1 aA | 183.4aB | 194.6 aA | 208.5 aA | 205.7 aA |
| Twice | 120.7 bB | 141.3 bA | 86.2 bB | 108.0 bA | 156.5 bA | 164.2 bA | 150.8 bA | 155.6 bA | |
| Thrice | 29.4 cB | 41.3 cA | 50.5cB | 69.9 cA | 92.6 cA | 110.1 cA | 54.7cB | 76.3 cA | |
| p > α | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| 2017 | Once | 186.7 aA | 167.9 aB | 90.8 aA | 91.0 aA | 179.6 aA | 168.7 aB | 207.9 aA | 207.6 aA |
| Twice | 89.8 bA | 70.4 bB | 69.4 bA | 71.6 bA | 148.1 bA | 134.0 bB | 122.7 bA | 121.5 bA | |
| Thrice | 32.9 cB | 37.7 cA | 41.5 cA | 44.6 cA | 42.3 cA | 41.4 cA | 45.6 cA | 47.7 cA | |
| p > α | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| Year | Cuts | Species and Harvest Regime | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Big Bluestem | Gamagrass | Indiangrass | Switchgrass | ||||||
| Same | Flipped | Same | Flipped | Same | Flipped | Same | Flipped | ||
| Basal | |||||||||
| ----------------------------------------------cm------------------------------------------------------- | |||||||||
| 2015 | Once | 21.7 a | 26.2 a | 19.1 aA | 19.8 bA | 19.4 aA | 21.3 aA | 18.6 aA | 21.0 aA |
| Twice | 17.4 aA | 18.5 bA | 19.4 aB | 29.8 aA | 18.4 aA | 22.7 aA | 17.3 aA | 18.8 aA | |
| Thrice | 17.2 aA | 19.6 bA | 24.4 aA | 28.7 aA | 19.4 aA | 21.8 aA | 17.2 aB | 21.1 aA | |
| p > α | 0.134 | <0.001 | 0.257 | <0.001 | 0.853 | 0.828 | 0.541 | 0.455 | |
| 2016 | Once | 26.8 aA | 27.6 bA | 39.7 aA | 36.9 aA | 28.1 aA | 30.2 aA | 25.1 aA | 28.1 aA |
| Twice | 28.2 aA | 29.5 abA | 37.4 aA | 38.6 aA | 30.2 aA | 32.2 aA | 27.5 aA | 29.4 aA | |
| Thrice | 28.3 aA | 31.3 aA | 37.9 aA | 37.5 aA | 32.2 aA | 34.0 aA | 26.4 aB | 31.1 aA | |
| p > α | 0.718 | 0.052 | 0.783 | 0.727 | 0.231 | 0.157 | 0.244 | 0.169 | |
| Canopy | |||||||||
| ----------------------------------------------cm------------------------------------------------------- | |||||||||
| 2015 | Once | 46.3 aA | 39.8 aA | 27.7 bA | 30.3 aA | 20.1 bA | 21.8 bA | 29.1 aA | 24.6 aB |
| Twice | 37.0 abA | 32.5 bA | 33.3 aA | 34.2 aA | 26.5 aA | 25.1 bA | 18.7 bA | 19.5 bA | |
| Thrice | 35.3 bA | 30.6 bA | 37.0 aA | 31.7 aA | 28.8 aA | 30.1 aA | 21.1 bA | 24.2 aA | |
| p > α | 0.101 | 0.035 | 0.257 | 0.332 | 0.022 | 0.007 | <0.001 | 0.007 | |
| 2016 | Once | 57.1 aB | 63.3 aA | 63.7 abB | 81.6 aA | 52.8 bA | 55.6 aA | 55.2 aA | 58.5 aA |
| Twice | 58.9 aA | 60.2 aA | 70.1 aA | 78.1 aA | 60.5 aA | 58.0 aA | 52.0 aB | 59.3 aA | |
| Thrice | 49.0 bA | 53.1 bA | 56.2 bB | 70.0 bA | 52.7 b | 52.5 a | 44.9 bB | 50.1 bA | |
| p > α | <0.001 | 0.002 | 0.009 | <0.001 | 0.069 | 0.310 | 0.001 | <0.001 | |
| Canopy: Basal Diameter | |||||||||
| --------------------------------------------------Ration-------------------------------------------------- | |||||||||
| 2015 | Once | 2.40 aA | 1.56 aB | 1.49 aA | 1.59 aA | 1.09 bA | 1.04 aA | 1.61 aA | 1.23 aB |
| Twice | 2.23 aA | 1.78 aA | 2.08 aA | 1.17 bA | 1.48 aA | 1.19 aA | 1.10 bA | 1.09 aA | |
| Thrice | 2.14 aA | 1.57 aB | 1.59 aA | 1.11 bB | 1.50 aA | 1.45 aA | 1.24 bA | 1.16 aA | |
| p > α | 0.858 | 0.408 | 0.316 | 0.005 | 0.038 | 0.141 | 0.008 | 0.655 | |
| 2016 | Once | 2.14 aA | 2.31 aA | 1.63 abB | 2.23 aA | 1.93 aA | 1.85 aA | 2.22 aA | 2.10 aA |
| Twice | 2.10 aA | 2.05 aA | 1.90 aA | 2.05 abA | 2.05 aA | 1.84 aA | 1.91 bA | 2.04 aA | |
| Thrice | 1.80 bA | 1.72 bA | 1.52 bB | 1.88 bA | 1.67 aA | 1.55 aA | 1.71 bA | 1.62 bA | |
| p > α | 0.054 | 0.001 | 0.045 | 0.021 | 0.163 | 0.112 | 0.007 | <0.001 | |
| Year | Cuts | Species and Harvest Regime | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Big Bluestem | Gamagrass | Indiangrass | Switchgrass | ||||||
| Same | Flipped | Same | Flipped | Same | Flipped | Same | Flipped | ||
| Light Interception | |||||||||
| ----------------------------------------------%------------------------------------------------------- | |||||||||
| 2015 | Once | 93.5 aA * | 93.0 aA | 79.6 aB | 89.0 aA | 76.7 aA | 82.0 aA | 79.2 aA | 84.4 aA |
| Twice | 86.2 bA | 83.6 bA | 66.9 bB | 80.8 bA | 67.0 bA | 73.0 bA | 48.1 bB | 62.1 bA | |
| Thrice | 81.2 bA | 79.4 bA | 58.2 cB | 73.9 cA | 55.5 bA | 62.0 cA | 38.9 cB | 55.4 bA | |
| p > α # | <0.001 | 0.011 | 0.007 | <0.001 | 0.003 | <0.001 | <0.001 | <0.001 | |
| 2016 | Once | 94.8 aA | 93.1 aA | 89.8 aA | 88.7 aA | 85.9 aA | 89.9 aA | 93.3 aA | 92.5 aA |
| Twice | 85.5 bA | 81.6 bA | 75.3 bA | 77.0 bA | 80.3 bA | 78.4 bA | 74.3 bA | 71.2 bA | |
| Thrice | 77.2 cA | 81.6 bA | 65.4 cB | 75.9 bA | 74.3 cA | 78.3 bA | 62.2 cA | 68.0 bA | |
| p > α | <0.001 | 0.009 | <0.001 | <0.001 | <0.001 | 0.002 | <0.001 | <0.001 | |
| Visual Obstruction Height | |||||||||
| ----------------------------------------------cm------------------------------------------------------- | |||||||||
| 2016 | Once | 186.3 aA | 169.7 aA | 81.4 aA | 78.7 aA | 181.9 aA | 172.9 aA | 208.7 aA | 210.0 aA |
| Twice | 73.5 bA | 85.0 bA | 69.4 bA | 71.2 aA | 111.4 bA | 132.8 bA | 136.5 bA | 148.9 bA | |
| Thrice | 21.3 cA | 18.0 cB | 35.3 cA | 35.0 bA | 29.6 cA | 28.9 cA | 32.7 cB | 41.4 cA | |
| p > α | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| 2017 | Once | 176.0 aA | 172.3 aA | 93.1 aA | 72.0 aA | 166.1 aA | 173.7 aA | 205.4 aA | 210.0 aA |
| Twice | 73.7 bA | 56.6 bB | 67.7 aA | 58.5 bA | 120.1 bA | 110.4 bA | 102.1 bA | 110.1 bA | |
| Thrice | 29.9 cA | 26.9 cA | 34.2 bA | 35.3 cA | 35.1 cA | 28.1 cB | 34.5 cA | 39.4 cA | |
| p > α | <0.001 | <0.001 | 0.002 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Temu, V.W.; Kering, M.K. Compensatory Structural Growth Responses of Early-Succession Native Warm-Season Grass Stands to Defoliation Management. Agronomy 2023, 13, 1280. https://doi.org/10.3390/agronomy13051280
Temu VW, Kering MK. Compensatory Structural Growth Responses of Early-Succession Native Warm-Season Grass Stands to Defoliation Management. Agronomy. 2023; 13(5):1280. https://doi.org/10.3390/agronomy13051280
Chicago/Turabian StyleTemu, Vitalis W., and Maru K. Kering. 2023. "Compensatory Structural Growth Responses of Early-Succession Native Warm-Season Grass Stands to Defoliation Management" Agronomy 13, no. 5: 1280. https://doi.org/10.3390/agronomy13051280