
Resistance Breeding to Northern Corn Leaf Blight with Dominant Genes, Polygene, and Their Combinations—Effects to Yield Traits
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Experiment Methods
2.2. Statistic Methods for Gene Effects Comparison
2.3. Statistic Methods for Prediction Losses and Gene Effects to Yield Traits
3. Results
3.1. ANOVA Results for E1 and E2
3.2. E1 Inbred Results
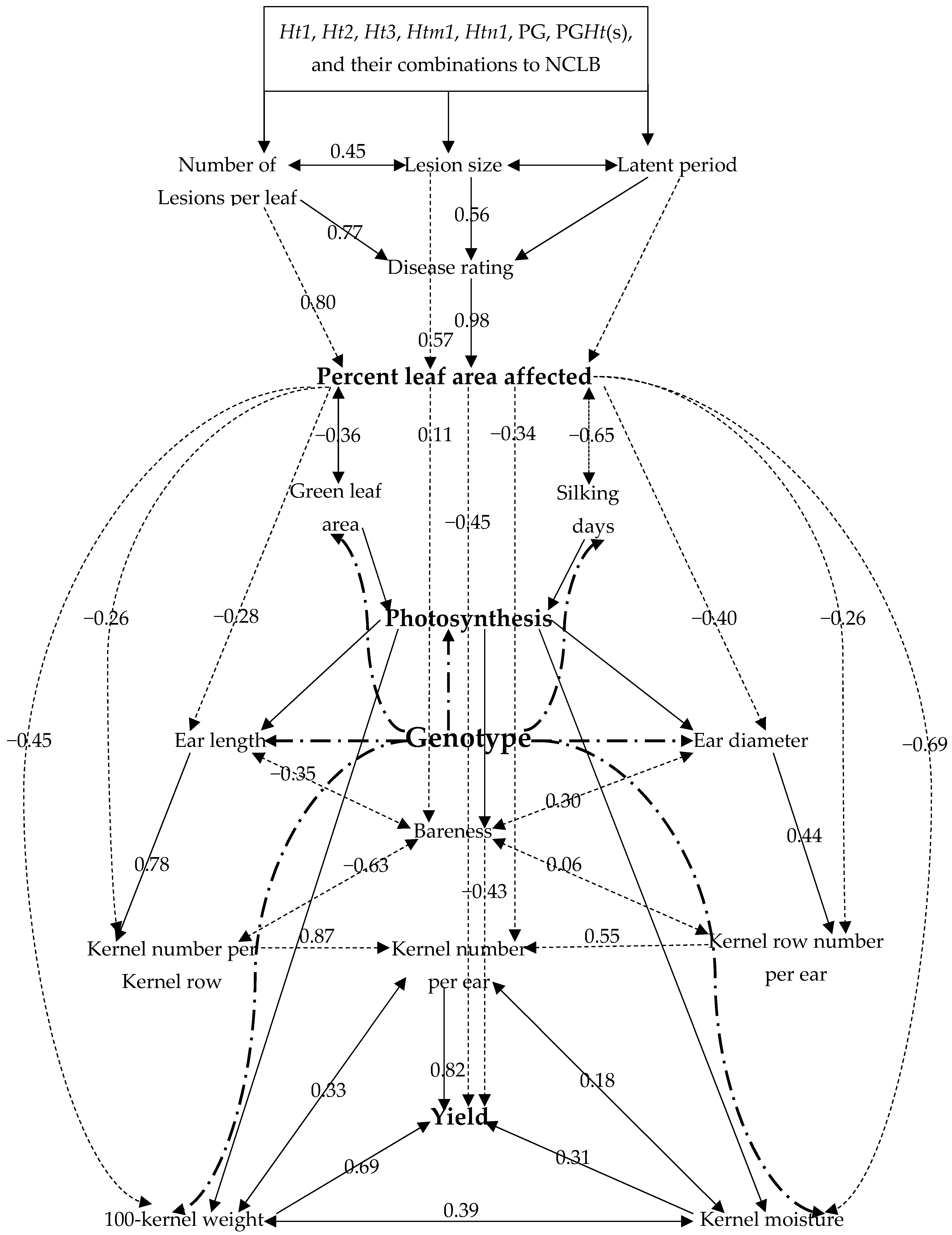
3.3. Correlation Coefficients among Disease and Yield Traits of Crosses from E1
3.4. Effects of Ht1, Ht2, Ht3, Htm1, Htn1 and Partial Resistance
3.5. Effects of PG, PGHt1, PGHt2, PGHt3, PGHtm1, and PGHtn1
3.6. Effects of Gene Combinations
3.7. Predict Losses of Yield Traits with PLAA and Effects of Resistant Genes to Yield Traits
3.8. E2 Results
3.9. Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, X.; Reid, L.M.; Woldemariam, T.; Tenuta, A.U. Survey of corn diseases and pests in ontario and québec in 2010. Can. Plant Dis. Surv. 2011, 91, 77–80. [Google Scholar]
- Jindal, K.K.; Reid, L.M.; Tenuta, A.U.; Woldemariam, T.; Zhu, X. Status of corn diseases in ontario, 2015 crop season. Can. Plant Dis. Surv. 2016, 96, 102–108. [Google Scholar]
- Zhu, X.; Kebede, A.; Tenuta, A.U.; Hooker, D.; Woldemariam, T. Status of corn diseases in ontario, 2022 crop season. Can. Plant Dis. Surv. 2023; 103, in press. [Google Scholar]
- Nwanosike, M.; Mabagala, R.; Kusolwa, P. Effect of northern leaf blight (exserohilum turcicum) severity on yield of maize (Zea mays l.) in morogoro, tanzania. Int. J. Sci. Res. 2015, 4, 466–475. [Google Scholar]
- Raymundo, A.; Hooker, A. Measuring the relationship between northern corn leaf blight and yield losses. Plant Dis. 1981, 65, 325–327. [Google Scholar] [CrossRef]
- Zhu, X.; Reid, L.M.; Presello, D.; Woldmariam, T. Survey of corn pests in ontario and quebec in 2000. Can. Plant Dis. Surv. 2001, 81, 48–50. [Google Scholar]
- Kloppers, R.; Tweer, S. Northern Corn Leaf Blight Fact Sheet; PANNA Seed (Pty) Ltd.: Greytown, South Africa, 2009. [Google Scholar]
- Wang, P.; Souma, K.; Kobayashi, Y.; Iwabuchi, K.; Sato, C.; Masuko, T. Influences of northern leaf blight on corn silage fermentation quality, nutritive value and feed intake by sheep. Anim. Sci. J. 2010, 81, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Hooker, A. A new type of resistance in corn to helminthosporium turcicum. Plant Dis. Report. 1961, 45, 780–781. [Google Scholar]
- Hooker, A. Inheritance of chlorotic-lesion resistance to helminthosporium turcicum in seedling corn. Phytopathology 1963, 53, 660–662. [Google Scholar]
- Hooker, A. Monogenic resistance in zea mays l. To helminthosporium turcicum 1. Crop Sci. 1963, 3, 381–383. [Google Scholar] [CrossRef]
- Ullstrup, A. Sources of resistance to northern corn leaf blight. Plant Dis. Rep 1963, 47, 107–108. [Google Scholar]
- Hooker, A. A second major gene locus in corn for chlorotic-lesion resistance to helminthosporium turicum 1. Crop Sci. 1977, 17, 132–135. [Google Scholar] [CrossRef]
- Hooker, A. Resistance to helminthosporium turcicum from tripsacum floridanum incorporated into corn. Maize Genet. Coop. Newsl. 1981, 55, 87–88. [Google Scholar]
- Gevers, H. A new major gene for resistance to helminthosporium turcicum leaf blight of maize [breeding, fungus diseases]. Plant Dis. Report. 1975, 59, 296–300. [Google Scholar]
- Robbins, W.; Warren, H. Inheritance of resistance to exserobilum turcicum in pi 20935, «mayorbela» variety of maize. Maydica 1993, 38, 209–213. [Google Scholar]
- Ogliari, J.B.; Guimarães, M.A.; Camargo, L.E.A. Chromosomal locations of the maize (zea mays l.) htp and rt genes that confer resistance to exserohilum turcicum. Genet. Mol. Biol. 2007, 30, 630–634. [Google Scholar] [CrossRef]
- Ogliari, J.B.; Guimarães, M.A.; Geraldi, I.O.; Camargo, L.E.A. New resistance genes in the zea mays: Exserohilum turcicum pathosystem. Genet. Mol. Biol. 2005, 28, 435–439. [Google Scholar] [CrossRef]
- Wang, H.; Xiao, Z.; Wang, F.; Xiao, Y.; Zhao, J.; Zheng, Y.; Qiu, F. Mapping of htnb, a gene conferring non-lesion resistance before heading toexserohilum turcicum (pass.), in a maize inbred line derived from the indonesian variety bramadi. Genet. Mol. Res. 2012, 11, 2523–2533. [Google Scholar] [CrossRef]
- Carson, M. A new gene in maize conferring the” chlorotic halo” reaction to infection by exserohilum turcicum. Plant Dis. 1995, 79, 717–720. [Google Scholar] [CrossRef]
- Hakiza, J.; Lipps, P.; Martin, S.S.; Pratt, R. Heritability and number of genes controlling partial resistance to exserohilum turcicum in maize inbred h99. Maydica 2004, 49, 173–182. [Google Scholar]
- Hooker, A.; Kim, S.K. Monogenic and multigenic resistance to helminthosporium turcicum in corn. Plant Dis. Report. 1973, 57, 586. [Google Scholar]
- Ferguson, L.M.; Carson, M. Spatial diversity of setosphaeria turcica sampled from the eastern united states. Phytopathology 2004, 94, 892–900. [Google Scholar] [CrossRef] [PubMed]
- Jordan, E.G.; Perkins, J.M.; Schall, R.; Pedersen, W. Occurrence of race 2 of exserohilum turcicum on corn in the central and eastern united states. Plant Dis. 1983, 67, 1163–1165. [Google Scholar] [CrossRef]
- Lipps, P.; Pratt, R.; Hakiza, J. Interaction of ht and partial resistance to exserohilum turcicum in maize. Plant Dis. 1997, 81, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Weems, J.D.; Bradley, C.A. Exserohilum turcicum race population distribution in the north central united states. Plant Dis. 2018, 102, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Jindal, K.K.; Tenuta, A.U.; Woldemariam, T.; Zhu, X.; Hooker, D.C.; Reid, L.M. Occurrence and distribution of physiological races of exserohilum turcicum in ontario, canada. Plant Dis. 2019, 103, 1450–1457. [Google Scholar] [CrossRef]
- Raymundo, A.; Hooker, A. Single and combined effects of monogenic and polygenic resistance on certain components of northern corn leaf blight development. Phytopathology 1982, 72, 99–103. [Google Scholar] [CrossRef]
- Hallauer, A.R.; Miranda, J.B. Quantitative Genetics in Maize Breeding, 2nd ed.; Iowa State University Press: Ames, IA, USA, 1988; pp. 115–158. [Google Scholar]
- Malik, H.N.; Malik, S.I.; Hussain, M.; Chughtai, S.; Javed, H.I. Genetic correlation among various quantitative characters in maize (zea mays l.) hybrids. J. Agric. Soc. Sci. 2005, 3, 262–265. [Google Scholar]
- Boćanski, J.; Srećkov, Z.; Nastasić, A. Genetic and phenotypic relationship between grain yield and components of grain yield of maize (zea mays l.). Genetika 2009, 41, 145–154. [Google Scholar] [CrossRef]
- Mogesse, W.; Zeleke, H. Estimates of combining ability and association among morpho-agronomic traits of single cross maize (zea mays l.) hybrids. J. Agric. Prod. 2022, 3, 78–87. [Google Scholar] [CrossRef]
- Jindal, K.K.; Zhu, X.; Tenuta, A.; Javed, N.; Daayf, F.; Reid, L.R. Maize inbreds for multiple resistance breeding against major foliar, ear and stalk rot diseases. Maydica 2019, 64, 22. [Google Scholar]
- Zhu, X.; Reid, L.M.; Woldemariam, T.; Wu, J.; Jindal, K.K.; Kebede, A. Resistance breeding to northern corn leaf blight with dominant genes, polygene, and their combinations—Effects to disease traits. Agronomy 2023, 13, 1096. [Google Scholar] [CrossRef]
- Zhu, X.; Reid, L.; Smith, M. A gene for resistance to northern leaf blight is inhibited by CO325. Maize Genet. Coop. Newsl. 2002, 76, 53–53. [Google Scholar]
- Reid, L.M.; Zhu, X. Screening Corn for Resistance to Common Diseases in Canada; AAFC Technical Bulletin, Agriculture and Agri-Food Canada: Ottawa, ON, Canada, 2003; pp. 19–21.
- Reid, L.M.; Zhu, X.; Morrison, M.J.; Woldemariam, T.; Voloaca, C.; Wu, J.; Xiang, K. A non-destructive method for measuring maize kernel moisture in a breeding program. Maydica 2010, 55, 163–171. [Google Scholar]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research; John Wiley & Sons: New York, NY, USA, 1984. [Google Scholar]
- IRRI. Rice Breeding Course. In Unit 6: Multi-Environment Trials—Design and Analysis. Unit 7: Broad-Sense Heritability Estimates and Selection Response; International Rice Research Institute. Available online: http://www.knowledgebank.irri.org/ricebreedingcourse (accessed on 1 March 2023).
- Welch, B.L. The generalization of ‘student’s’problem when several different population varlances are involved. Biometrika 1947, 34, 28–35. [Google Scholar]
- De Smith, M. A Comprehensive Handbook of Statistical Concept, Techniques and Software Tools. 2018. Available online: www.statsref.com (accessed on 1 March 2023).
- OCC. 2016 Crop Year Report, 2016 Hybrid Corn Performance Trials. Available online: http://gocorn.net/v2006/CornReports/2016cornreport/2016performancetrials.html# (accessed on 1 March 2023).
- Reid, L.M.; Zhu, X.; Jindal, K.K.; Woldemariam, T.; Wu, J.; Voloaca, C. CO468, CO469, CO470, CO471, CO472, and CO473 corn inbred lines with improved northern corn leaf blight resistance. Can. J. Plant Sci. 2019, 99, 972–984. [Google Scholar] [CrossRef]


{kind=link}
| Name/Code | Purpose | Heterotic Group | Assumed Resistant Gene | PLAA (%) | Yield (t h−1) | KM (%) | KNPE | 100 KW (g) |
|---|---|---|---|---|---|---|---|---|
| A619 | Tester | NS-Oh43 | None | 84.1 | 3.5 | 40.4 | 194 | 24.0 |
| A619Ht1 | Tester | NS-Oh43 | Ht1Ht1 | 42.1 | 3.2 | 39.4 | 183 | 24.3 |
| A619Ht2 | Tester and R source | NS-Oh43 | Ht2Ht2 | 35.0 | 5.4 | 45.2 | 269 | 29.1 |
| A619Ht3 | Tester | NS-Oh43 | Ht3Ht3 | 40.2 | 4.1 | 39.4 | 197 | 26.5 |
| A632HTN | R Source | SS-B14 | Htn1Htn1 | 46.2 | 5.0 | 31.1 | 252 | 26.2 |
| A553N | R Source | Tropical | Htn1Htn1 | 16.5 | 0.7 | 56.9 | 48 | 23.1 |
| 73353 | R Source | Tropical | Htm1Htm1 | 11.5 | 3.2 | 39.6 | 165 | 32.0 |
| H102 | R Source | Tropical | Htm1Htm1 | 8.0 | 3.8 | 55.6 | 205 | 24.2 |
| Pa91 | Inbred check | SS | None | 39.2 | 3.8 | 55.6 | 260 | 24.2 |
| Pa91Ht1 | Inbred check | SS | Ht1Ht1 | 20.1 | 3.1 | 56.8 | 221 | 21.5 |
| Pa91Ht2 | R Source | SS | Ht2Ht2 | 16.0 | 2.6 | 60.3 | 204 | 23.7 |
| Pa91Ht3 | R Source | SS | Ht3Ht3 | 18.4 | 5.5 | 59.7 | 332 | 24.1 |
| CO353 | MR check | SS | Unknown | 21.2 | 5.7 | 47.0 | 246 | 29.4 |
| CO388 | Line and tester | SS-B73 | None | 80.0 | 4.3 | 32.5 | 253 | 23.7 |
| BLT01 | Line and tester | SS-B73 | Htn1Htn1 | 57.8 | 5.8 | 34.2 | 305 | 28.8 |
| BLT02 | Line and tester | SS-B73 | Htn1Htn1 | 51.8 | 5.5 | 30.4 | 295 | 30.3 |
| BLT03 | Line and tester | SS-B73 | Htm1Htm1 | 56.5 | 4.9 | 31.1 | 262 | 27.3 |
| CO428 | Line | NS-Oh43 | PG PG | 9.7 | 5.9 | 42.9 | 287 | 26.7 |
| BLT05 | Line | NS-Oh43 | PGHtm1 PGHtm1 | 24.2 | 6.8 | 40.0 | 294 | 27.9 |
| BLT06 | Line | NS-Oh43 | PGHtn1 PGHtn1 | 7.0 | 5.9 | 42.1 | 304 | 25.6 |
| BLT07 | Line | NS-Oh43 | PGHt2 PGHt2 | 11.0 | 6.0 | 42.6 | 304 | 25.4 |
| BLT09 | Line | NS-Oh43 | PGHtn1 PGHtn1 | 10.8 | 4.6 | 46.2 | 299 | 26.9 |
| BLT10 | Line | NS-Oh43 | PGHtm1 PGHtm1 | 2.7 | 6.1 | 42.4 | 292 | 29.7 |
| BLT11 | Line | NS-Oh43 | PGHt1 PGHt1 | 5.9 | 5.2 | 38.2 | 298 | 23.7 |
| BLT12 | Line | NS-Oh43 | PGHt2 PGHt2 | 2.7 | 6.4 | 42.4 | 302 | 29.8 |
| BLT13 | Line | NS-Oh43 | PGHt3 PGHt3 | 3.3 | 6.3 | 42.5 | 304 | 29.6 |
| CL30 | Tester | NS-Flint | None | 97.7 | 1.3 | 16.2 | 155 | 16.2 |
| CO442 | Tester and S check | NS-Iodent | None | 96.6 | 4.4 | 17.2 | 402 | 14.9 |
| T1 | Tester | NS-Iodent | None | 97.7 | 3.4 | 15.8 | 203 | 24.6 |
| T2 | Tester | NS-Iodent | None | 92.2 | 3.6 | 18.4 | 281 | 18.4 |
| T3 | Tester | NS-Iodent | Partial | 69.6 | 4.7 | 23.3 | 280 | 24.8 |
| T4 | Tester | SS-B14 | None | 90.0 | 5.0 | 20.5 | 306 | 23.2 |
| T5 | Tester | SS-B14 | None | 92.7 | 3.6 | 14.5 | 251 | 18.9 |
| CV% | 26.4 | 15.7 | 12.7 | 12.5 | 8.4 | |||
| LSD(0.05) | 12.9 | 1.8 | 4.7 | 59.5 | 2.8 | |||
| H | 0.93 | 0.94 | 0.94 | 0.93 | 0.90 |
| Genotypes | Female/Male Genes | # | PLAA (%) | Yield (t h−1) | KM (%) | KNPE | 100KW (g) |
|---|---|---|---|---|---|---|---|
| 7 susceptible hybrid checks | -/- | 42 | 77.7a | 10.7bc | 20.6a | 505.9bc | 26.1a b |
| Resistant hybrid check1 | Ht?/Ht? | 6 | 35.8 b | 12.7a b | 22.3a | 590.6a | 28.1a b |
| Resistant hybrid check2 | Ht?/Ht? | 6 | 29.8 b | 14.1a | 20.8a | 581.7a b | 30.6a |
| Resistant hybrid check3 | Ht?/Ht? | 6 | 70.7a | 9.1c | 15.8 b | 474.7c | 24.5 b |
| Mean | 60 | 68.0 | 11.1 | 20.3 | 518.8 | 26.6 | |
| Same female cross with different males | |||||||
| CO388 × A619 | -/- | 6 | 75.8a | 12.9a | 29.7a | 468.1a | 31.7 b |
| CO388 × A619Ht1 | -/Ht1 | 6 | 39.8 b | 13.7a | 31.6a | 499.5a | 33.6a b |
| CO388 × A619Ht2 | -/Ht2 | 6 | 46.2 b | 13.3a | 33.6a | 481.2a | 35.6a |
| CO388 × A619Ht3 | -/Ht3 | 6 | 43.3 b | 14.0a | 33.5a | 492.5a | 34.1a b |
| SS × NS Mean | 24 | 51.3A | 13.4A b | 32.1 b | 485.3A | 33.7A | |
| (BLT01-BLT03) × A619 | (Htm1, Htn1)/- | 18 | 65.2a | 13.3 b | 30.7 b | 505.6a | 33.3 b |
| (BLT01-BLT03) × A619Ht1 | (Htm1, Htn1)/Ht1 | 18 | 33.9 b | 14.1a b | 31.1 b | 511.8a | 35.8a b |
| (BLT01-BLT03) × A619Ht2 | (Htm1, Htn1)/Ht2 | 18 | 32.2 b | 14.6a | 36.6a | 525.7a | 36.5a |
| (BLT01-BLT03) × A619Ht3 | (Htm1, Htn1)/Ht3 | 18 | 25.4 b | 14.9a | 35.2a | 522.2a | 35.9a b |
| SS × NS Mean | 72 | 39.2A b | 14.2A | 33.4 b | 516.3A | 35.4A | |
| CO428 × A619 | PG/- | 6 | 50.8a | 8.7a | 36.5a | 371.8a | 28.4a b |
| CO428 × A619Ht1 | PG/Ht1 | 6 | 24.9 b | 8.1a | 37.3a | 383.3a | 26.0c |
| CO428 × A619Ht2 | PG/Ht2 | 6 | 21.1 b | 9.2a | 40.4a | 394.2a | 29.8a |
| CO428 × A619Ht3 | PG/Ht3 | 6 | 18.3 b | 8.8a | 38.6a | 403.3a | 28.1 b |
| NS × NS Mean | 24 | 28.8 b | 8.7 b | 38.2A | 388.2 b | 28.1 b | |
| (BLT05-BLT13) × A619 | PGHt(s)/- | 42 | 48.3a | 8.6 b | 34.1 b | 375.2 b | 29.3 b |
| (BLT05-BLT13) × A619Ht1 | PGHt(s)/Ht1 | 42 | 22.1 b | 9.7a | 35.3a b | 405.2a | 30.4a |
| (BLT05-BLT13) × A619Ht2 | PGHt(s)/Ht2 | 42 | 12.7c | 10.2a | 37.7a | 419.9a | 30.5a |
| (BLT05-BLT13) × A619Ht3 | PGHt(s)/Ht3 | 42 | 15.7c | 9.7a | 40.0a | 407.3a | 30.2a |
| NS × NS Mean | 168 | 24.7 b | 9.5 b | 36.8A | 401.9 b | 30.1 b | |
| CO428 × CO388 | PG/- | 6 | 27.0a | 13.5a | 35.8 b | 503.7a | 33.6a |
| CO428 × BLT03 | PG/Htm1 | 6 | 18.1 b | 13.8a | 37.3a | 502.4a | 33.8a |
| CO428 × BLT02 | PG/Htn1 | 6 | 18.1 b | 12.9a | 37.4a | 452.7 b | 35.9a |
| NS × SS Mean | 18 | 21.0 b | 13.4A b | 36.8A | 486.3A | 34.4A | |
| (BLT05-BLT13) × CO388 | PGHt(s)/- | 42 | 25.4a | 13.3a | 36.7a | 509.1a | 33.5a |
| (BLT05-BLT13) × BLT03 | PGHt(s)/Htm1 | 30 | 17.3 b | 13.7a | 36.1a | 489.1a | 33.8a |
| (BLT05-BLT13) × (BLT01, BLT02) | PGHt(s)/Htn1 | 48 | 22.2a | 13.4a | 35.4a | 495.5a | 33.9a |
| NS × SS Mean | 120 | 22.1 b | 13.4A b | 36.0A | 498.7A | 33.7A | |
| Different females crossed with the same male | |||||||
| CO388 × 4 testers | -/- | 24 | 83.9a | 9.9a | 23.1 b | 427.7a | 27.6 b |
| BLT03 × 4 testers | Htm1/- | 24 | 66.2 b | 10.8a | 25.1a b | 462.2a | 30.3a |
| (BLT01, BLT02) × 4 testers | Htn1/- | 60 | 61.1 b | 10.7a | 26.9a | 454.3a | 30.2a |
| SS × (early flint, Iodent) Mean | 120 | 68.1A | 10.5 b | 25.5E | 449.6 b | 29.6D | |
| CO388 × A619 | -/- | 6 | 75.8a | 12.9a | 29.7a | 468.1a | 31.7a |
| BLT03 × A619 | Htm1/- | 6 | 61.7 b | 15.0a | 31.1a | 570.5a | 34.2a |
| (BLT01, BLT02) × A619 | Htn1/- | 12 | 67.0 b | 12.5a | 30.5a | 473.2a | 32.8a |
| SS × NS Mean | 24 | 67.9A | 13.2A | 30.5C | 496.3A b | 32.9BC | |
| CO388 × A619Ht(s) | -/(Ht1-Ht3) | 18 | 43.1a | 13.6a | 32.9a | 491.1 b | 34.4 b |
| BLT03 × A619Ht(s) | Htm1(Ht1-Ht3) | 18 | 31.1 b | 14.7a | 33.1a | 525.2a | 36.0a |
| (BLT01, BLT02) × A619Ht(s) | Htn1/(Ht1-Ht3) | 36 | 30.2 b | 14.4a | 34.9a | 517.2a b | 36.1a |
| SS × NS Mean | 72 | 33.7C | 14.3A | 33.9 b | 512.7A | 35.6A | |
| CO388 × T3 | -/Partial | 6 | 69.1a | 9.7 b | 26.6a | 434.1a | 29.3a |
| BLT03 × T3 | Htm1/Partial | 6 | 41.7 b | 11.2a | 27.9a | 460.9a | 32.6a |
| (BLT01, BLT02) × T3 | Htn1/Partial | 12 | 39.6 b | 10.8a b | 28.1a | 462.4a | 31.1a |
| SS × Iodent Mean | 24 | 47.5 b | 10.6 b | 27.7D | 455.0A b | 31.0C | |
| CO428 × A619 | PG/- | 6 | 50.8a | 8.7a | 36.5a | 371.8a | 28.4a |
| BLT11 × A619 | PGHt1/- | 6 | 49.0a | 8.9a | 34.0a | 405.6a | 27.6a |
| (BLT07, BLT12) × A619 | PGHt2/- | 12 | 53.6a | 8.4a | 33.9a | 367.5a | 28.5a |
| BLT13 × A619 | PGHt3/- | 6 | 46.5a | 9.2a | 33.2a | 389.2a | 30.2a |
| (BLT05, BLT10) × A619 | PGHtm1/- | 12 | 42.5a | 8.8a | 35.1a | 367.1a | 31.2a |
| BLT06 × A619 | PGHtn1/- | 6 | 50.5a | 7.5a | 33.9a | 362.6a | 27.7a |
| NS × NS Mean | 48 | 48.6 b | 8.6D | 34.4 b | 374.8D | 29.2D | |
| CO428 × A619Ht(s) | PG/(Ht1-Ht3) | 18 | 21.5a | 8.7 b | 38.7a | 393.6 b | 28.0 b |
| BLT11 × A619Ht(s) | PGHt1/(Ht1-Ht3) | 18 | 16.3 b | 10.0a | 37.3a | 437.2a | 28.4a b |
| (BLT07, BLT12) × A619Ht(s) | PGHt2/(Ht1-Ht3) | 36 | 15.3 b | 9.6a b | 38.6a | 396.0 b | 30.9a |
| BLT13 × A619Ht(s) | PGHt3/(Ht1-Ht3) | 18 | 16.9 b | 10.5a | 37.0a | 452.4a | 29.1a b |
| (BLT05, BLT10) × A619Ht(s) | PGHtm1/(Ht1-Ht3) | 36 | 16.9 b | 10.0a | 37.3a | 405.0 b | 31.1a |
| BLT06 × A619Ht(s) | PGHtn1/(Ht1-Ht3) | 18 | 17.2 b | 9.4a b | 37.5a | 384.1 b | 31.0a |
| NS × NS Mean | 144 | 17.0E | 9.7C | 37.8A | 408.6C | 30.1CD | |
| CO428 × CO388 | PG/- | 6 | 27.0a | 13.5a | 35.8a | 503.7a | 33.6a b |
| (BLT07, BLT12) × CO388 | PGHt2/- | 12 | 21.3a | 14.0a | 39.2a | 500.9a | 35.2a |
| BLT13 × CO388 | PGHt3/- | 6 | 27.6a | 13.3a | 37.5a | 533.0a | 31.6 b |
| (BLT05, BLT10) × CO388 | PGHtm1/- | 12 | 23.4a | 13.9a | 34.9a | 513.6a | 35.0a |
| (BLT06, BLT09) × CO388 | PGHtn1/- | 12 | 30.5a | 12.1a | 35.5a | 501.0a | 31.2 b |
| NS × SS Mean | 48 | 25.6D | 13.3A | 36.5A b | 508.5A | 33.5 b | |
| CO428 × (BLT02, BLT03) | PG/(Htm1, Htn1) | 12 | 18.1a b | 13.4 b | 37.3a b | 477.6 b | 34.8a |
| BLT11 × (BLT02, BLT03) | PGHt1/(Htm1, Htn1) | 18 | 14.4 b | 15.4a | 39.5a | 562.6a | 34.6a b |
| (BLT07, BLT12) × (BLT02, BLT03) | PGHt2/(Htm1, Htn1) | 24 | 21.6a | 13.6a b | 35.6 b | 470.9 b | 33.4a b |
| BLT13 × (BLT02, BLT03) | PGHt3/(Htm1, Htn1) | 6 | 20.9a b | 13.7a b | 38.2a | 527.2a | 33.8a b |
| (BLT05, BLT10) × (BLT02, BLT03) | PGHtm1/(Htm1, Htn1) | 18 | 17.0a b | 13.7a b | 36.3 b | 481.9 b | 35.6a |
| (BLT06, BLT9) × (BLT02, BLT03) | PGHtn1/(Htm1, Htn1) | 12 | 20.7a b | 13.1 b | 35.9 b | 497.9a b | 32.3 b |
| NS × SS Mean | 90 | 18.6E | 13.9A | 37.0A | 499.7A b | 34.1A b | |
| LSD(0.05) | 18.4 | 2.7 | 6.4 | 81.6 | 3.7 | ||
| Heritability | 0.92 | 0.84 | 0.89 | 0.82 | 0.83 | ||
| Combined results and their percentages shown in parentheses | |||||||
| (CO388, BLT01-BLT03) × A619 (CO428, BLT05-BLT13) × A619 | - | 72 | 55.0a (100.0) | 10.1b (100.0) | 33.1c (100.0) | 415.3b (100.0) | 30.4b (100.0) |
| (CO388, BLT01-BLT03) × A619Ht1 (CO428, BLT05-BLT13) × A619Ht1 | Ht1 | 72 | 26.8b (47.2) | 11.0a (108.7) | 34.1b (102.9) | 437.9ab (105.4) | 31.7a (104.2) |
| (CO388, BLT01-BLT03) × A619Ht2 (CO428, BLT05-BLT13) × A619Ht2 | Ht2 | 72 | 21.1c (38.3) | 11.5a (113.3) | 37.3a (112.7) | 449.3a (108.2) | 32.4a (106.5) |
| (CO388, BLT01-BLT03) × A619Ht3 (CO428, BLT05-BLT13) × A619Ht3 | Ht3 | 72 | 20.7c (37.6) | 11.3a (111.3) | 38.1a (115.1) | 442.8a (106.6) | 31.8a (104.5) |
| CO388 × (A619, A619Ht(s), 5 testers) and (CO428, BLT05-BLT13) × CO388 | - | 102 | 47.9a (100.0) | 12.3b (100.0) | 31.7a (100.0) | 479.6b (100.0) | 31.9b (100.0) |
| BLT03 × (A619, A619Ht(s), 5 testers) and (CO428, BLT05-BLT13) × BLT03 | Htm1 | 90 | 35.2b (73.3) | 13.1a (106.2) | 32.3a (101.9) | 498.3a (103.9) | 33.2a (104.0) |
| (BLT01, BLT02) × (A619, A619Ht(s), 5 testers); and (CO428, BLT05-BLT13) × (BLT01, BLT02) | Htn1 | 162 | 34.4b (71.8) | 12.8a (103.4) | 33.0a (103.4) | 489.8a (102.1) | 33.4a (104.5) |
| CO428 × (A619, A619Ht(s)) and CO428 × (CO388, BLT02, BLT03) | PG | 42 | 25.5a (100.0) | 10.7b (100.0) | 37.6a (100.0) | 430.2b (100.0) | 30.8c (100.0) |
| BLT11 × (A619, A619Ht(s)) and BLT11 × (CO388, BLT02, BLT03) | PGHt1 | 42 | 21.4a (84.1) | 12.2a (114.2) | 37.6a (99.9) | 493.7a (114.8) | 30.6c (99.5) |
| (BLT07, BLT12) × (A619, A619Ht(s)) and (BLT07, BLT12) × (CO388, BLT02, BLT03) | PGH2 | 84 | 23.4a (91.9) | 11.2a (104.5) | 37.1a (98.8) | 428.3b (99.6) | 31.9a (103.6) |
| BLT13 × (A619, A619Ht(s)) and BLT13 × (CO388, BLT02, BLT03) | PGHt3 | 36 | 23.0a (90.4) | 11.8a (110.3) | 36.6a (97.3) | 479.8a (111.5) | 31.0c (100.6) |
| (BLT05, BLT10) × (A619, A619Ht(s)) and (BLT05, BLT10) × (CO388, BLT02, BLT03) | PGHtm1 | 78 | 21.3a (83.6) | 11.4a (106.5) | 36.4a (96.7) | 435.3b (101.2) | 33.1a (107.6) |
| (BLT06, BLT09) × (A619, A619Ht(s)) and (BLT06, BLT09) × (CO388, BLT02, BLT03) | PGHtn1 | 60 | 24.3a (95.5) | 10.6b (99.3) | 36.4a (96.8) | 435.0b (101.1) | 30.7c (99.7) |
| Genotype | Female/Male Genes | # | PLAA (%) | Yield (t h−1) | KM (%) | KNPE | 100KW (g) |
|---|---|---|---|---|---|---|---|
| Susceptible hybrid checks | -/- | 42 | 77.7a | 10.7 b | 20.6a | 505.9 b | 26.1a |
| Resistant hybrid checks | Ht?/Ht? | 18 | 45.5 b | 11.9a | 19.6a | 549.0a | 27.7a |
| CO388 × 4 testers | -/- | 24 | 83.9a | 9.9 b | 23.1 b | 427.7 b | 27.6 b |
| (BLT01- BLT03) × 4 testers | (Htm1, Htn1)/- | 72 | 62.8 b | 10.7a | 26.3a | 456.9a | 30.2a |
| CO388 × T3 | -/Partial | 6 | 69.1a | 9.7 b | 26.6a | 434.1a | 29.3b |
| (BLT01- BLT03) × T3 | (Htm1, Htn1)/Partial | 18 | 40.3 b | 10.9a | 28.0a | 461.9a | 31.6a |
| CO388 × A619 | -/- | 6 | 75.8a | 12.9b | 29.7b | 468.1 b | 31.7b |
| CO388 × A619Ht(s) | -/(Ht1, Ht2, Ht3) | 18 | 43.1c | 13.6a b | 32.9a | 491.1a b | 34.4a b |
| (BLT01- BLT03) × A619 | (Htm1, Htn1)/- | 18 | 65.2 b | 13.3 b | 30.7a b | 505.6a | 33.3a b |
| (BLT01- BLT03) × A619Ht(S) | (Htm1, Htn1)/(Ht1, Ht2, Ht3) | 54 | 30.5d | 14.5a | 34.3a | 519.9a | 36.1a |
| CO428 × A619 | PG/- | 6 | 50.8a | 8.7 b | 36.5a b | 371.8 b | 28.4b |
| CO428 × A619Ht(s) | PG/(Ht1, Ht2, Ht3) | 18 | 21.5 b | 8.7 b | 38.7a | 393.6a b | 28.0 b |
| (BLT05-BLT13) × A619 | PGHt(s)/- | 42 | 48.3a | 8.6 b | 34.1 b | 375.2a b | 29.3a b |
| (BLT05-BLT13) × A619Ht(S) | PGHt(s)/(Ht1, Ht2, Ht3) | 138 | 16.2 b | 9.9a | 37.7a | 411.9a | 30.3a |
| CO428 × CO388 | PG/- | 6 | 27.0a | 13.5a | 35.8a | 503.7a | 33.6a |
| CO428 × (BLT02, BLT03) | PG/(Htm1, Htn1) | 12 | 18.1 b | 13.4a | 37.3a | 477.6a | 34.8a |
| (BLT05-BLT13) × CO388 | PGHt(s)/- | 42 | 25.4a | 13.3a | 36.7a | 509.1a | 33.5a |
| (BLT05-BLT13) × (BLT02, BLT03) | PGHt(s)/(Htm1, Htn1) | 72 | 18.7 b | 14.0a | 36.9a | 503.1a | 34.0a3) |
| Combined results and their percentages shown in parentheses | |||||||
| CO388 × (A619, CL30, CO442, T1, T2) | -/- | 30 | 82.3a (100.0) | 10.5b (100.0) | 24.4b (100.0) | 435.7b (100.0) | 28.4b (100.0) |
| (BLT01- BLT03) × (A619, CL30, CO442, T1, T2) | (Htm1, Htn1)/- | 90 | 63.3b (76.9) | 11.2ab (107.5) | 27.2b (111.6) | 466.7b (107.1) | 30.8b (108.5) |
| CO388 × (A619Ht(s), T3) | -/(Ht1, Ht2, Ht3, Partial) | 24 | 49.6c (60.3) | 12.7a (121.1) | 31.3a (128.3) | 476.8b (109.4) | 33.1a (116.7) |
| (BLT01- BLT03) × (A619Ht(s), T3) | (Htm1, Htn1)/(Ht1, Ht2, Ht3, Partial) | 72 | 33.0d (40.1) | 13.6a (130.1) | 32.7a (134.2) | 505.4a (116.0) | 34.9a (123.0) |
| CO428 × (A619, CO388) | PG/- | 12 | 38.9a (100.0) | 11.1a (100.0) | 36.2ab (100.0) | 437.7a (100.0) | 31.0a (100.0) |
| (BLT05–BLT13) × (A619, CO388) | PGHt(s)/- | 84 | 36.9a (94.8) | 10.9a (98.7) | 35.4b (97.9) | 442.2a (101.0) | 31.4a (101.3) |
| CO428 × (A619Ht(s), BLT02, BLT03) | PG/Ht(s) | 30 | 20.1b (51.7) | 10.6a (95.2) | 38.2a (105.6) | 427.2a (97.6) | 30.7a (99.1) |
| (BLT05–BLT13) × (A619Ht(s), BLT02, BLT03) | PGHt(s)/Ht(s) | 210 | 17.3b (44.4) | 11.2a (101.2) | 37.3a (103.3) | 441.9a (100.9) | 31.5a (101.7) |
| Genotypes | Traits | # | Trait Range | Regression | PL (%) | PERG (%) | ||
|---|---|---|---|---|---|---|---|---|
| Max/Mean/Min | m | β | r | Max/Mean/Min | ||||
| 7 susceptible and 3 resistant hybrid checks | PLAA | 60 | 97.7/68.0/9.2 | |||||
| Yield | 60 | 17.8/11.1/6.0 | 16.106 | −0.074 | −0.713 ** | 44.9/31.3/4.2 | 40.7 | |
| KM | 60 | 35.3/20.3/10.8 | 26.034 | −0.084 | −0.423 ** | 31.4/21.9/2.9 | 28.5 | |
| KNPE | 60 | 710/518/325 | 631.23 | −1.652 | −0.550 ** | 25.6/17.8/2.4 | 23.1 | |
| 100 KW | 60 | 36.9/26.6/16.1 | 35.021 | −0.124 | −0.670 ** | 34.7/24.2/3.3 | 31.4 | |
| (CO388, BLT01, BLT02, BLT03) × (A619, A619Ht1, A619Ht2, A619Ht3) | PLAA | 96 | 97.7/42.2/5.5 | |||||
| Yield | 96 | 19.7/14.0/5.5 | 18.017 | −0.095 | −0.697 ** | 51.4/22.2/2.9 | 48.5 | |
| KM | 96 | 51.7/33.1/25.6 | 38.561 | −0.130 | −0.637 ** | 32.9/14.2/1.8 | 31.1 | |
| KNPE | 96 | 709/509/253 | 614.27 | −2.503 | −0.649 ** | 39.8/17.2/2.2 | 37.6 | |
| 100 KW | 96 | 40.8/35.0/25.9 | 38.869 | −0.093 | −0.722 ** | 23.3/10.1/1.3 | 32.0 | |
| (CO428, BLT05, BLT06, BLT07, BLT09, BLT10, BLT11, BLT12, BLT13) × (CO388, BLT01, BLT02, BLT03) | PLAA | 156 | 64.1/21.1/1.6 | |||||
| Yield | 156 | 19.7/13.7/5.4 | 15.318 | −0.079 | −0.485 ** | 32.9/10.8/0.8 | 32.1 | |
| KM | 156 | 60.2/36.5/25.7 | 40.201 | −0.175 | −0.424 ** | 27.9/9.2/0.7 | 27.2 | |
| KNPE | 156 | 772/505/354 | 547.36 | −2.026 | −0.444 ** | 23.7/7.8/0.6 | 23.1 | |
| 100 KW | 156 | 41.0/33.9/24.0 | 35.348 | −0.068 | −0.331 ** | 12.3/4.1/0.3 | 12.0 | |
| (CO428, BLT05, BLT06, BLT07, BLT09, BLT10, BLT11, BLT12, BLT13) × (A619, A619Ht1, A619Ht2, A619Ht3) | PLAA | 204 | 94.3/24.3/1.2 | |||||
| Yield | 204 | 13.3/9.5/4.1 | 10.886 | −0.059 | −0.591 ** | 50.9/13.1/0.6 | 50.3 | |
| KM | 204 | 64.6/37.0/23.4 | 41.282 | −0.175 | −0.509 ** | 40.0/10.3/0.5 | 39.5 | |
| KNPE | 204 | 590/402/238 | 444.47 | −1.765 | −0.526 ** | 37.5/9.7/0.5 | 37.0 | |
| 100 KW | 204 | 36.8/29.8/20.1 | 31.529 | −0.071 | −0.481 ** | 21.3/5.5/0.3 | 21.0 | |
| Overall regression models for predicted losses (PL) based on above results | Yield | PL (%) for yield = (0.5042)PLAA, r = 0.995 ** | ||||||
| KM | PL (%) for KM = (0.3634)PLAA, r = 0.979 ** | |||||||
| KNPE | PL (%) for KNPE = (0.3483)PLAA, r = 0.959 ** | |||||||
| 100 KW | PL (%) for 100 KW = (0.2721)PLAA, r = 0.943 ** | |||||||
| Genotype | Assumed Resistant Genes | PLAA | RY (%) | ||||
|---|---|---|---|---|---|---|---|
| None | Once | Twice | None | Once | Twice | ||
| Grand mean | 0.1 a | 18.2 b | 37.1 c | 89.1 a | 93.4 b | 103.1 c | |
| Check mean | 0.2 a | 36.3 b | 54.3 c | 100.0 a | 100.0 a | 100.0 a | |
| S checks | -/- | 0.2 a | 44.4 b | 66.2 c | 95.9 a | 94.2 a | 95.5 a |
| R checks | Ht?/Ht? | 0.2 a | 17.3 b | 26.7 b | 109.5 a | 113.4 a | 110.6 a |
| Cross mean | 0.1 a | 16.1 b | 35.1 c | 87.8 a | 92.6 b | 103.4 c | |
| S × S cross | -/- | 0.2 a | 39.7 b | 70.0 c | 99.5 a | 94.1 a | 93.0 a |
| S × R cross | -/(Ht1, Ht2, Ht3) | 0.1 a | 13.6 b | 25.9 c | 103.7 a | 118.7 b | 134.8 c |
| S × Partial cross | -/Partial | 0.3 a | 23.0 b | 51.8 c | 91.6 a | 91.7 a | 96.6 a |
| R × S cross | (Htm1, Htn1)/- | 0.2 a | 23.1 b | 50.6 c | 102.4 a | 103.8 a | 101.8 a |
| R × R cross | (Htm1, Htn1)/(Ht1, Ht2, Ht3) | 0.0 a | 11.6 b | 14.9 c | 101.4 a | 109.9 b | 135.9 c |
| R × Partial cross | (Htm1, Htn1)/Partial | 0.1 a | 8.8 b | 31.8 c | 100.8 a | 102.4 a | 98.8 a |
| PG × S cross | PG/- | 0.1 a | 7.9 b | 25.7 c | 87.6 ab | 82.2 a | 99.5 b |
| PG × R cross | PG/Ht(s) | 0.0 a | 5.0 b | 9.5 c | 67.8 a | 92.4 b | 94.8 b |
| PGHt(s) × S cross | PGHt(s)/- | 0.1 a | 19.6 b | 46.5 c | 84.3 a | 87.1 a | 99.2 b |
| PGHt(s) × R cross | PGHt(s)/Ht(s) | 0.1 a | 3.9 b | 11.0 c | 69.1 a | 77.2 b | 96.1 c |
| Real grand mean | 0.1 a | 18.2 b | 37.1 c | 9.5 a | 10.4 b | 13.0 c | |
| Real check mean | 0.2 a | 36.3 b | 54.3 c | 10.7 a | 11.1 ab | 12.6 b | |
| Real cross mean | 0.1 a | 16.2 b | 35.3 c | 9.3 a | 10.3 b | 13.0 c | |
| CV(%) | 199.8 | 40.4 | 30.3 | 12.6 | 14.6 | 10.6 | |
| LSD(0.05) | 0.4 | 11.8 | 18.1 | 18.2 | 22.0 | 17.6 | |
| LSD(0.05) among treatments | 4.4 | 6.3 | |||||
| Heritability | 0.19 | 0.92 | 0.94 | 0.90 | 0.89 | 0.91 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, X.; Kebede, A.; Woldemariam, T.; Wu, J.; Jindal, K.K.; Reid, L.M. Resistance Breeding to Northern Corn Leaf Blight with Dominant Genes, Polygene, and Their Combinations—Effects to Yield Traits. Agronomy 2023, 13, 1269. https://doi.org/10.3390/agronomy13051269
Zhu X, Kebede A, Woldemariam T, Wu J, Jindal KK, Reid LM. Resistance Breeding to Northern Corn Leaf Blight with Dominant Genes, Polygene, and Their Combinations—Effects to Yield Traits. Agronomy. 2023; 13(5):1269. https://doi.org/10.3390/agronomy13051269
Chicago/Turabian StyleZhu, Xiaoyang, Aida Kebede, Tsegaye Woldemariam, Jinhe Wu, Krishan K. Jindal, and Lana M. Reid. 2023. "Resistance Breeding to Northern Corn Leaf Blight with Dominant Genes, Polygene, and Their Combinations—Effects to Yield Traits" Agronomy 13, no. 5: 1269. https://doi.org/10.3390/agronomy13051269