
Rain Shelter Cultivation Reduces Root Rot Incidence of Panax notoginseng by Altering Root Exudates and Bacterial Communities under Micro-Irrigation and Fertilization

Abstract
:1. Introduction
2. Materials and Methods
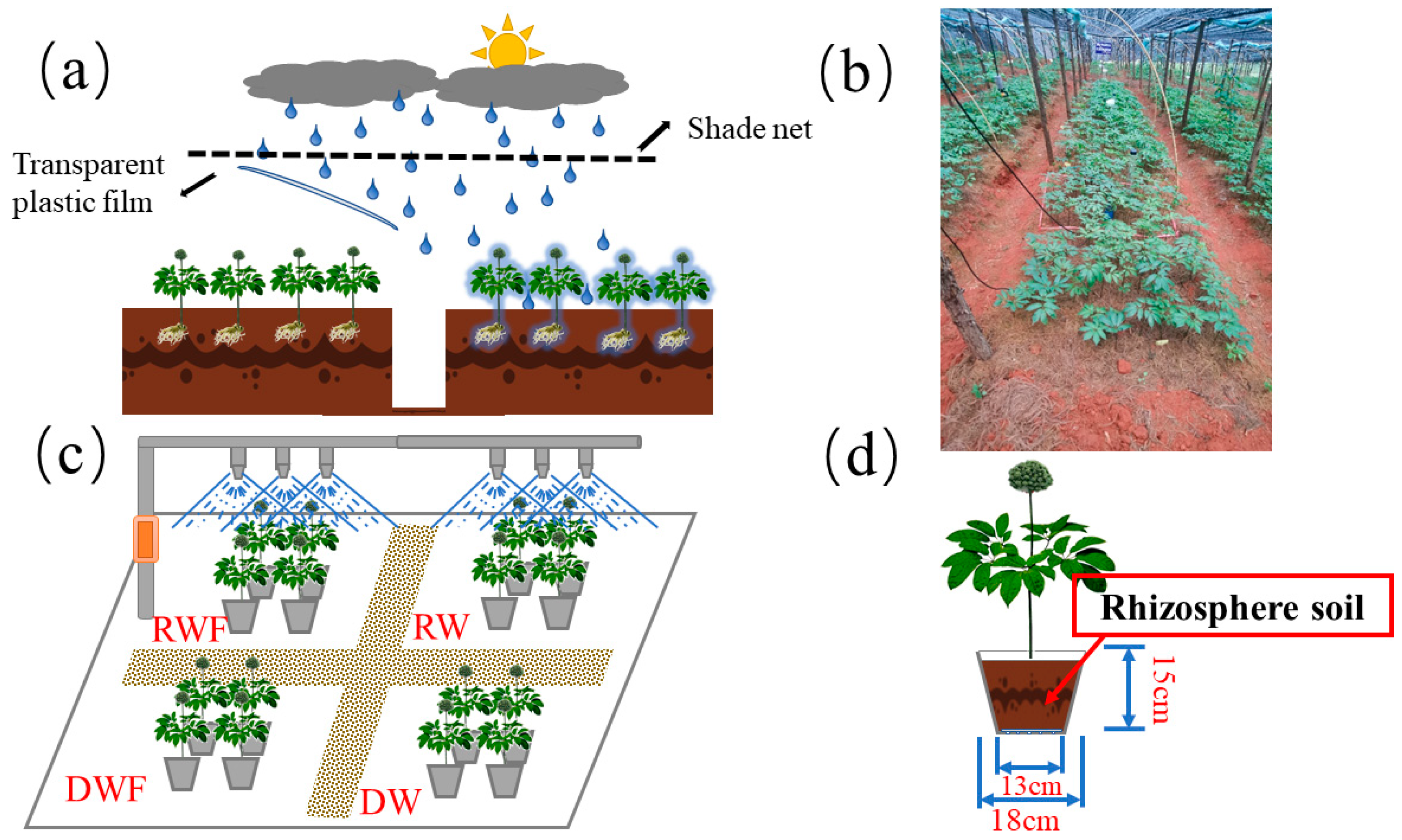
2.1. Experimental Site Descriptions
2.2. Experimental Design
2.3. Sampling and Determination
2.4. DNA Extraction and Illumina MiSeq Sequencing
2.5. Analysis of the Sequencing Data
2.6. Extraction and Determination of Soil Organic Acids
2.7. Statistics of the Incidence Rate of Root Rot
2.8. Statistical Analysis
3. Results
3.1. Soil Physical and Chemical Properties
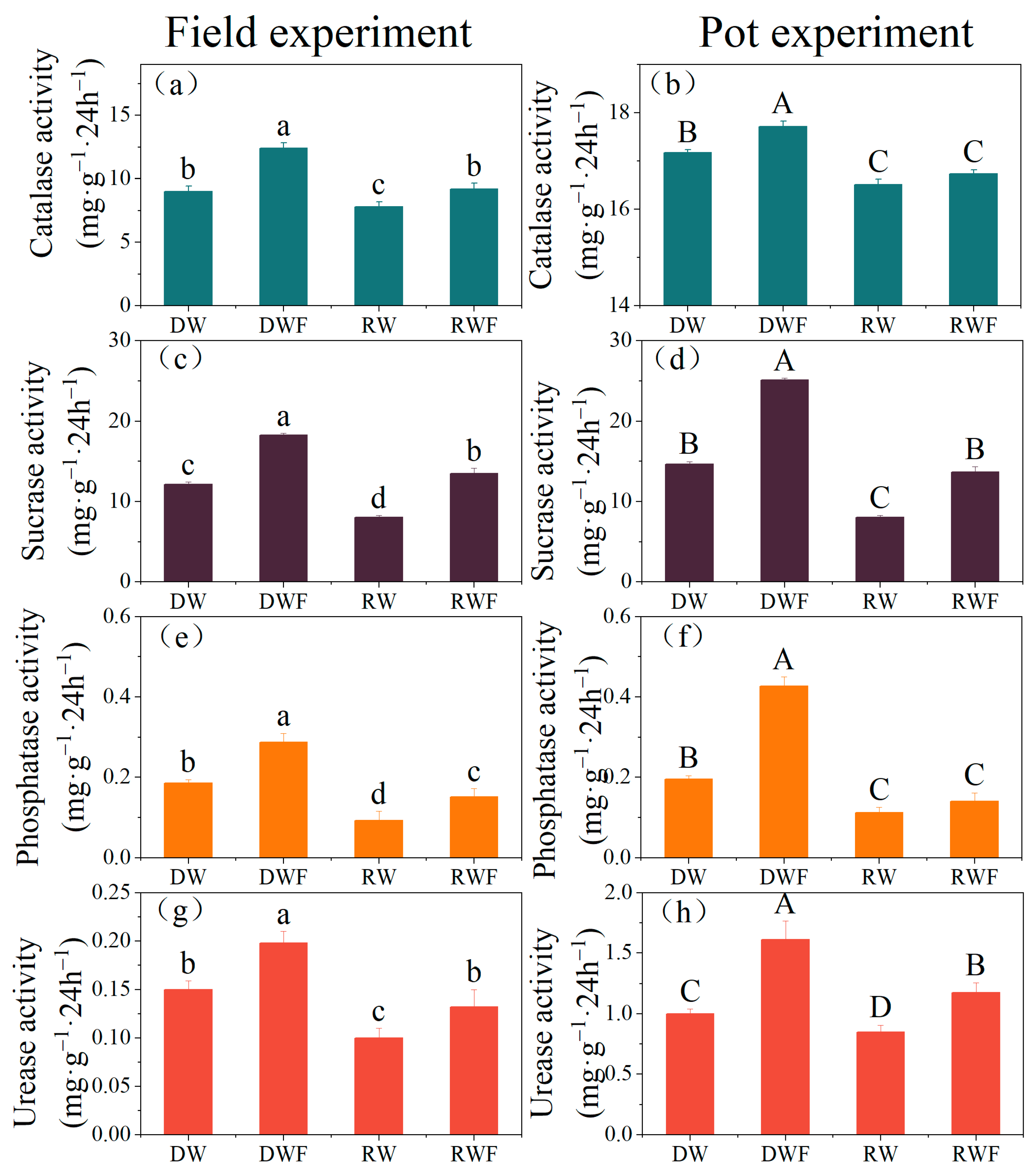
3.2. Soil Enzyme Activities
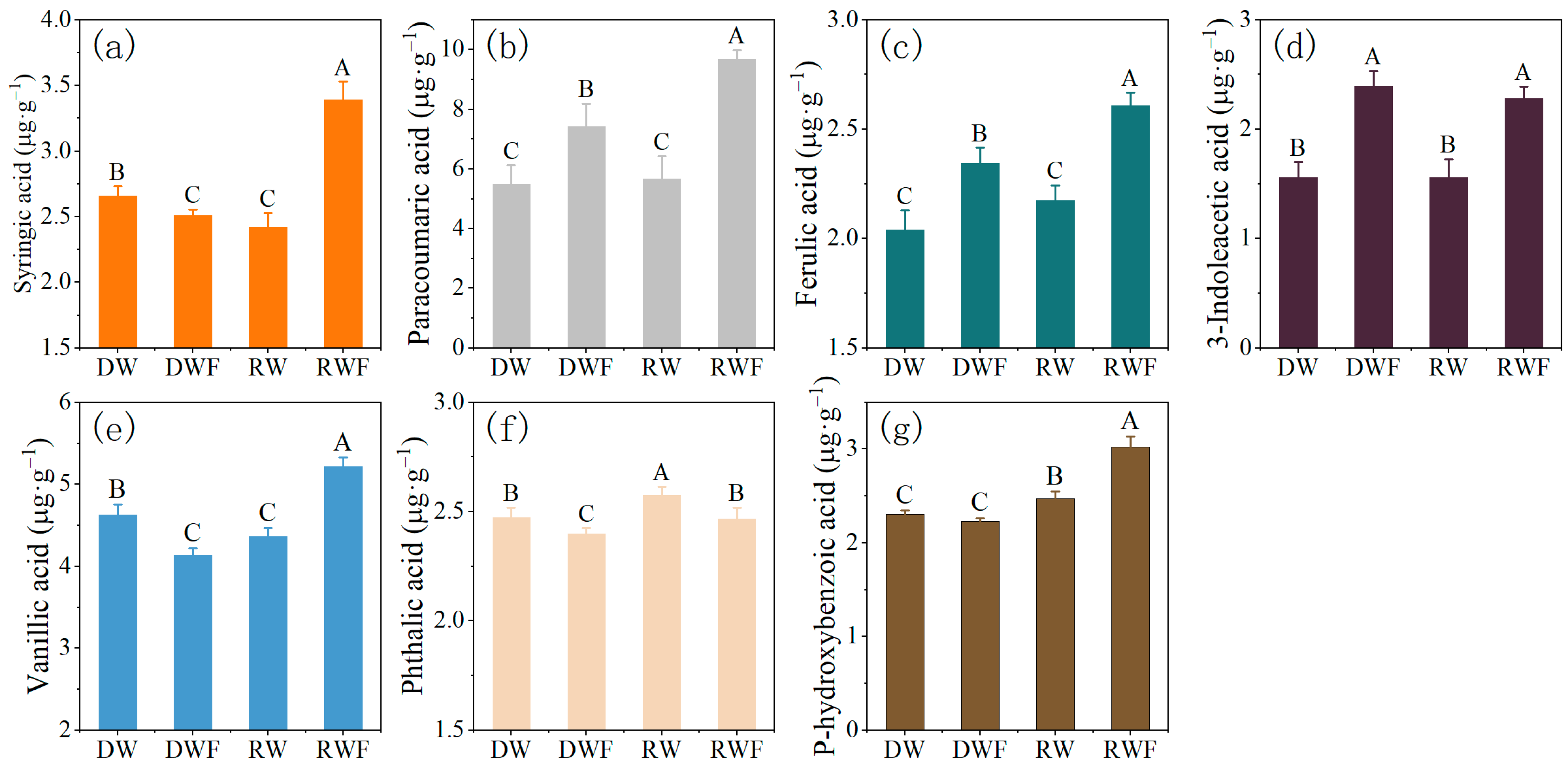
3.3. Root Exudates
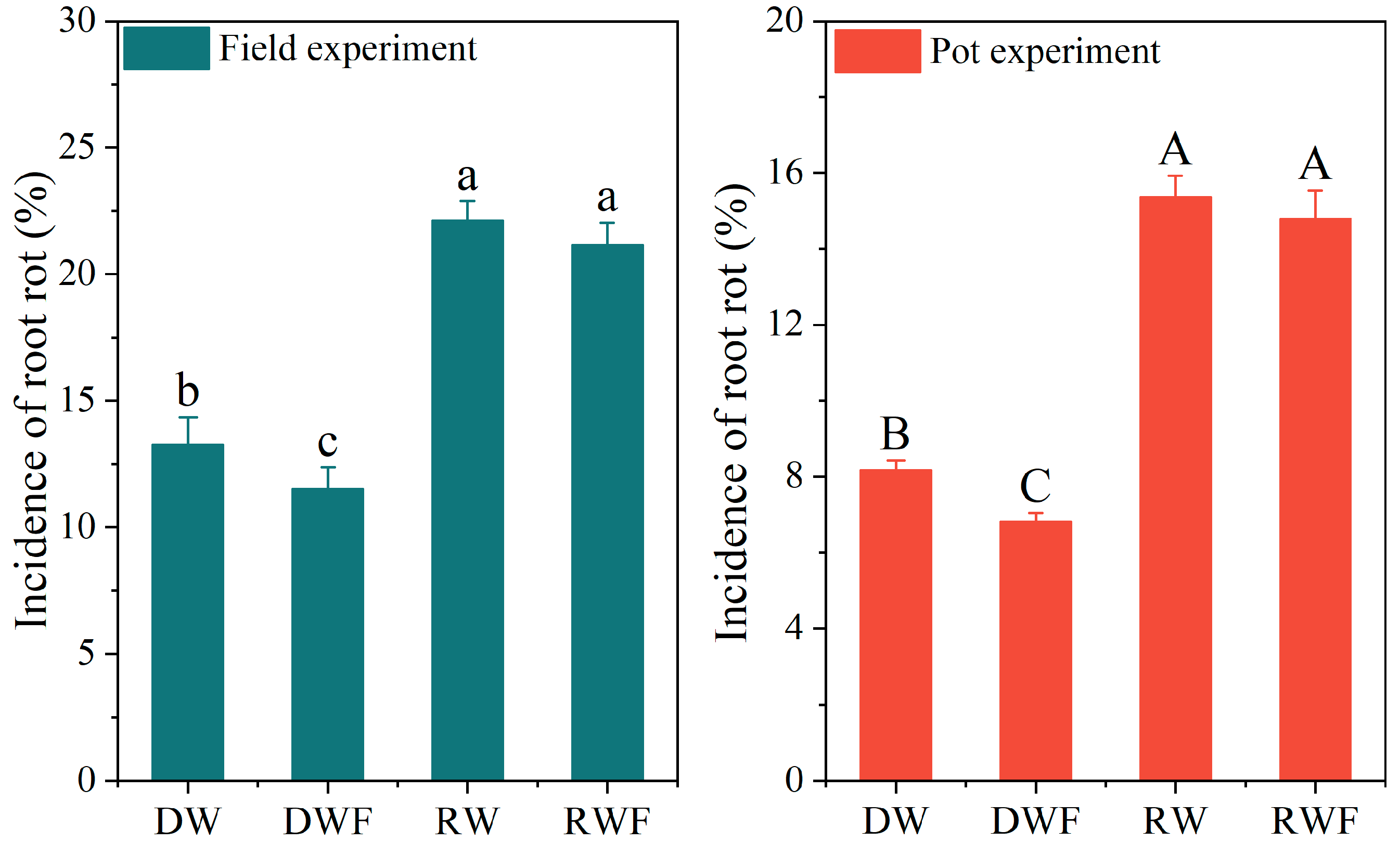
3.4. Root Rot Incidence of P. notoginseng
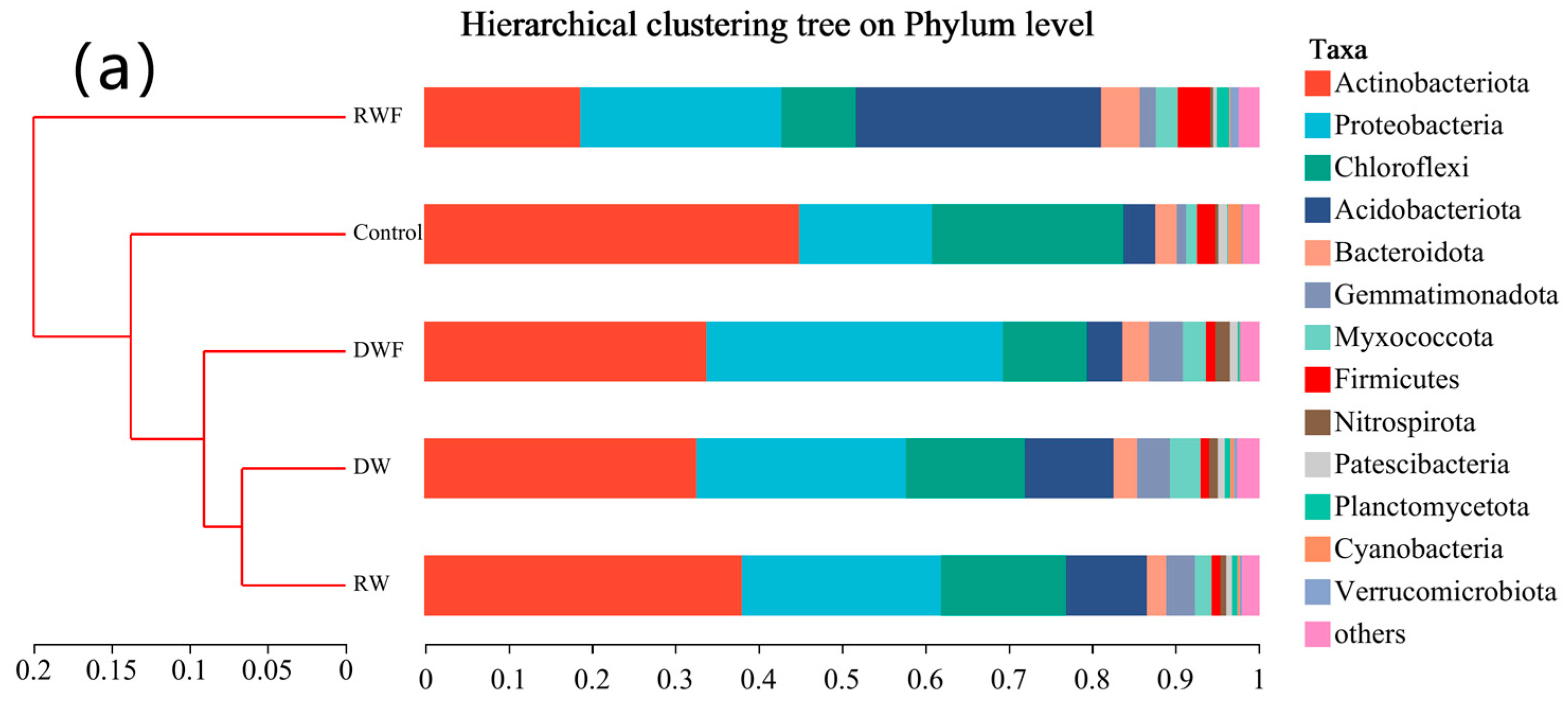
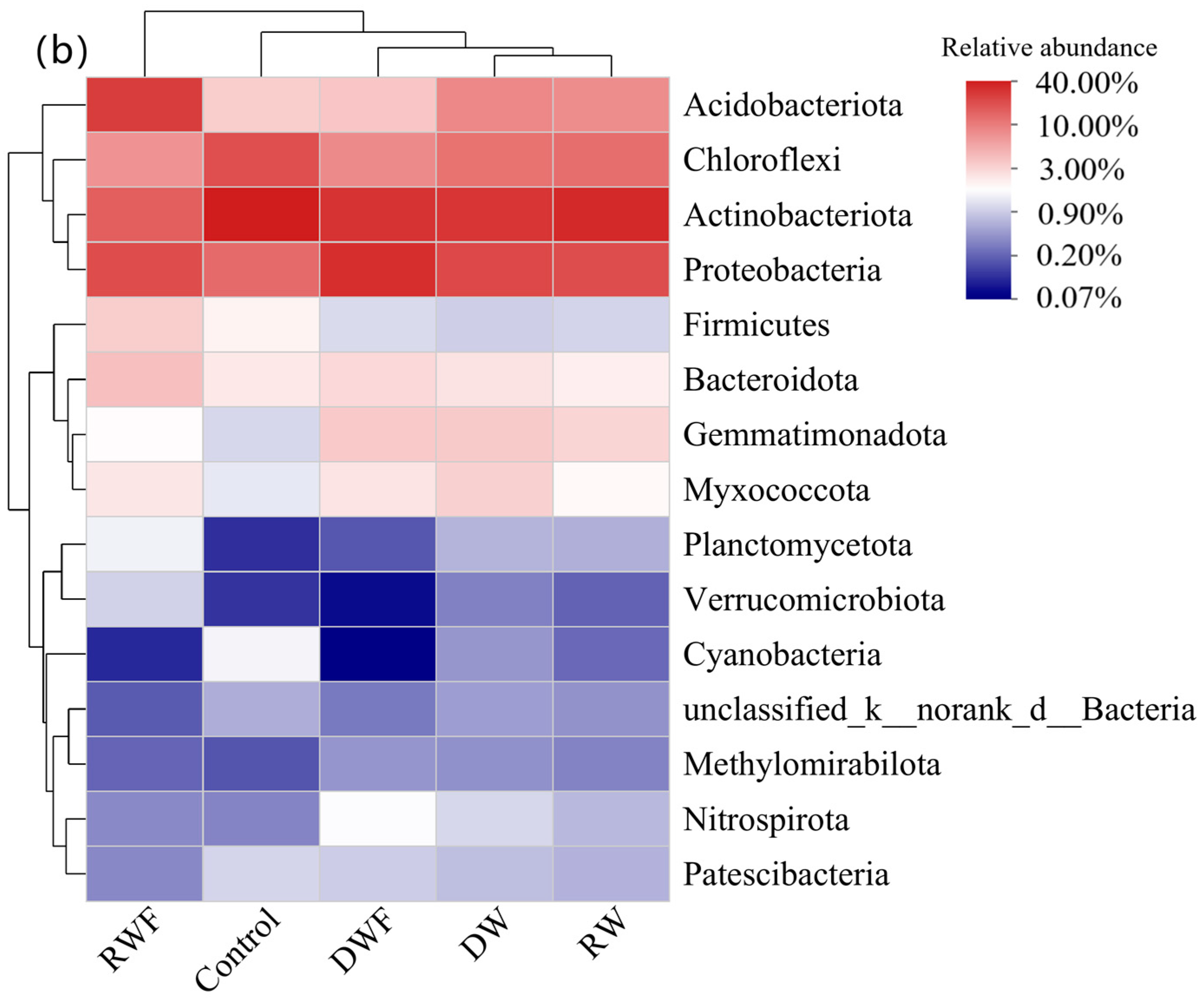
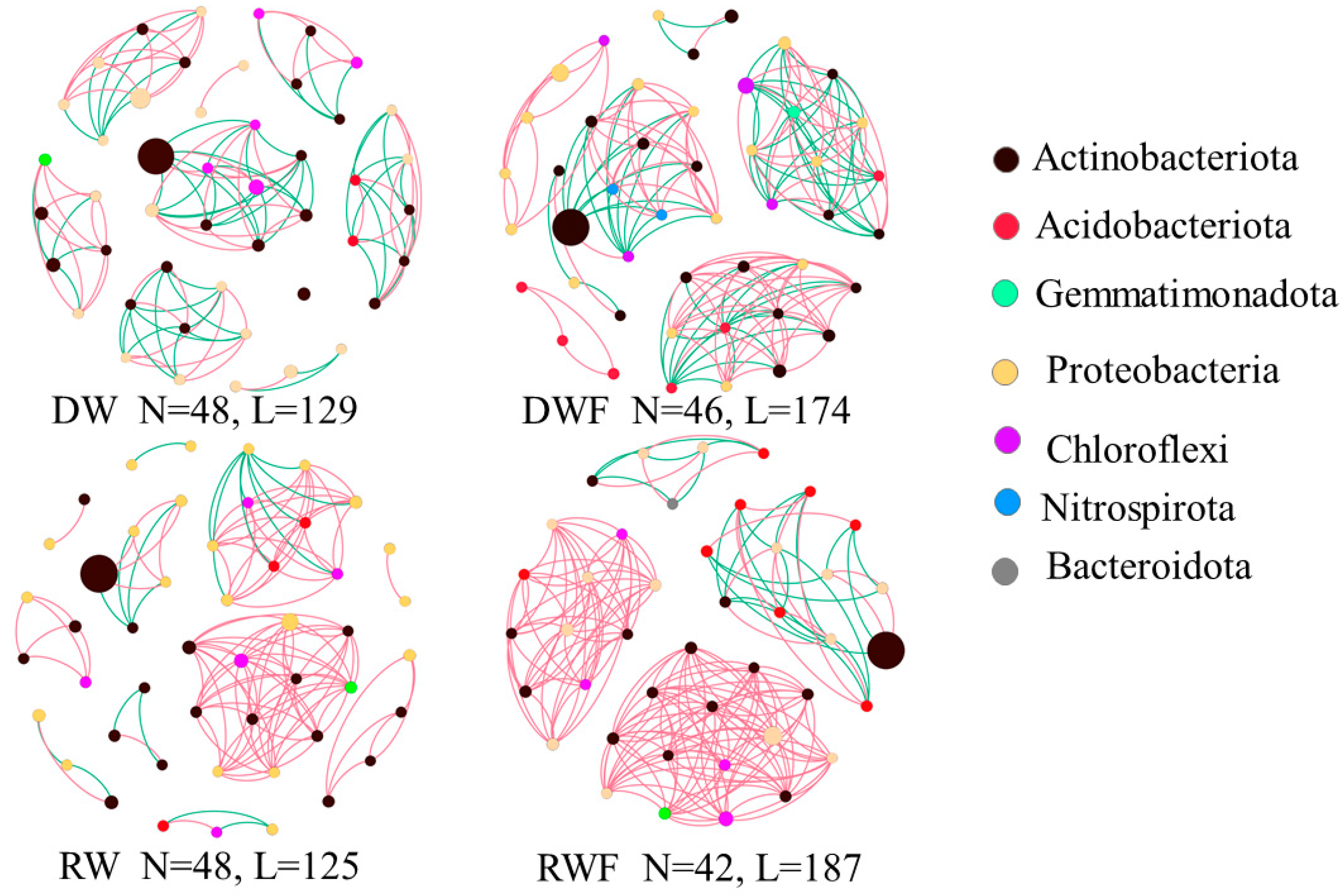
3.5. Bacterial Community in Rhizosphere Soil of P. notoginseng
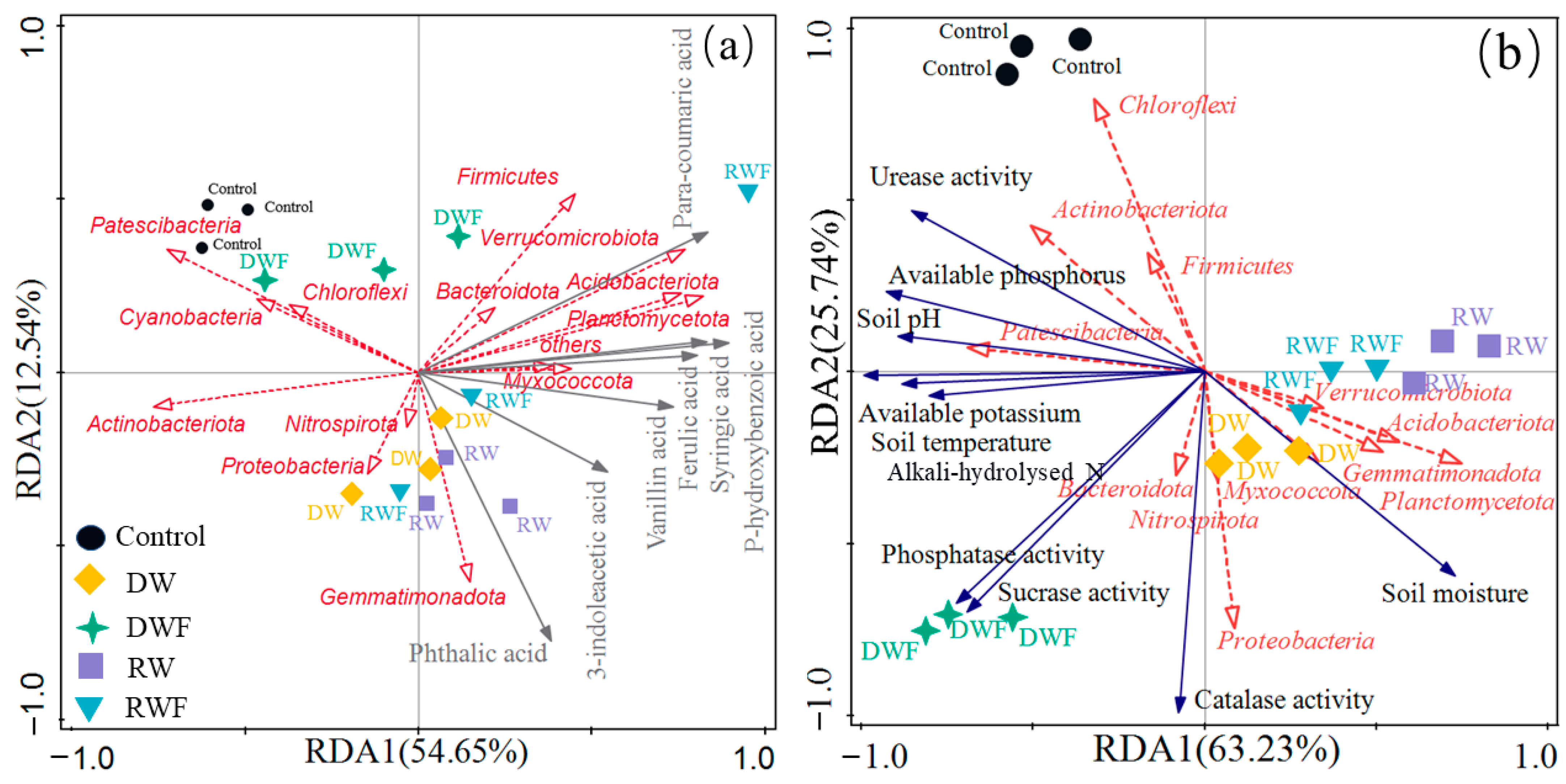
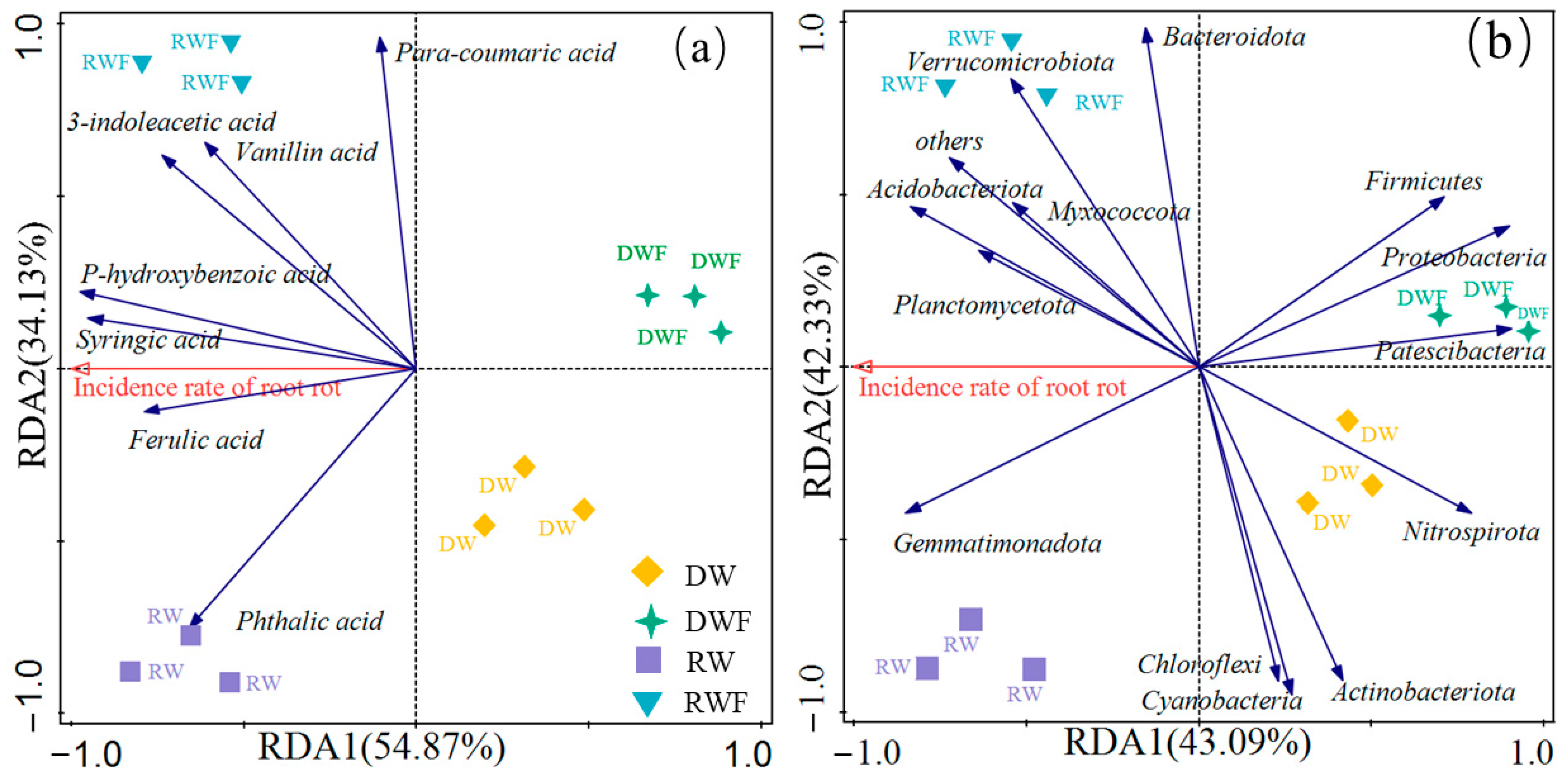
3.6. Correlation Studies
4. Discussion
4.1. Effects of Root Exudates and Soil Physical and Chemical Properties on the Microbial Community of P. notoginseng
4.2. Effects of Bacterial Communities on Root Rot in P. notoginseng
4.3. Rain Shelter with Irrigation and Fertilization Could Effectively Reduce the Secretion of Organic Acids of P. notoginseng and Reduce the Incidence of Root Rot
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, T.; Guo, R.; Zhou, G.; Zhou, X.; Kou, Z.; Sui, F.; Li, C.; Tang, L.; Wang, Z. Traditional uses, botany, phytochemistry, pharmacology and toxicology of Panax notoginseng (Burk.) FH Chen: A review. J. Ethnopharmacol. 2016, 188, 234–258. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhang, X.; Xu, Y.; Mei, X.; Jiang, B.; Liao, J.; Yin, Z.; Zheng, J.; Zhao, Z.; Fan, L. Autotoxic ginsenosides in the rhizosphere contribute to the replant failure of Panax notoginseng. PLoS ONE 2015, 10, e0118555. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Xia, Q.; Li, Y.; Zhao, J.; Yang, S.; Wei, F.; Huang, X.; Zhang, J.; Cai, Z. Root rot-infected Sanqi ginseng rhizosphere harbors dynamically pathogenic microbiotas driven by the shift of phenolic acids. Plant Soil 2021, 465, 385–402. [Google Scholar] [CrossRef]
- Liao, P.; Liu, D.; Xu, T.-R.; Yang, Y.; Cui, X. Soil water stress attenuate the growth and development but enhance the saponin synthesis of Panax notogesing during flowering stage. Ind. Crops Prod. 2017, 108, 95–105. [Google Scholar] [CrossRef]
- Tan, Y.; Cui, Y.; Li, H.; Kuang, A.; Li, X.; Wei, Y.; Ji, X. Rhizospheric soil and root endogenous fungal diversity and composition in response to continuous Panax notoginseng cropping practices. Microbiol. Res. 2017, 194, 10–19. [Google Scholar] [CrossRef]
- Li, H.; Wu, J.; Chen, C.; Xin, W.; Zhang, W. Simultaneous determination of 74 pesticide residues in Panax notoginseng by QuEChERS coupled with gas chromatography tandem mass spectrometry. Food Sci. Hum. Wellness 2021, 10, 241–250. [Google Scholar] [CrossRef]
- Zhalnina, K.; Louie, K.B.; Hao, Z.; Mansoori, N.; da Rocha, U.N.; Shi, S.; Cho, H.; Karaoz, U.; Loqué, D.; Bowen, B.P. Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly. Nat. Microbiol. 2018, 3, 470–480. [Google Scholar] [CrossRef]
- Yuan, J.; Zhao, J.; Wen, T.; Zhao, M.; Li, R.; Goossens, P.; Huang, Q.; Bai, Y.; Vivanco, J.M.; Kowalchuk, G.A. Root exudates drive the soil-borne legacy of aboveground pathogen infection. Microbiome 2018, 6, 156. [Google Scholar] [CrossRef]
- Cipollini, D.; Rigsby, C.M.; Barto, E.K. Microbes as targets and mediators of allelopathy in plants. J. Chem. Ecol. 2012, 38, 714–727. [Google Scholar] [CrossRef]
- Li, C.; Liu, L.; Zheng, L.; Yu, Y.; Mushinski, R.M.; Zhou, Y.; Xiao, C. Greater soil water and nitrogen availability increase C: N ratios of root exudates in a temperate steppe. Soil Biol. Biochem. 2021, 161, 108384. [Google Scholar] [CrossRef]
- Aulakh, M.; Wassmann, R.; Bueno, C.; Kreuzwieser, J.; Rennenberg, H. Characterization of root exudates at different growth stages of ten rice (Oryza sativa L.) cultivars. Plant Biol. 2001, 3, 139–148. [Google Scholar] [CrossRef]
- Badri, D.V.; Chaparro, J.M.; Zhang, R.; Shen, Q.; Vivanco, J.M. Application of natural blends of phytochemicals derived from the root exudates of Arabidopsis to the soil reveal that phenolic-related compounds predominantly modulate the soil microbiome. J. Biol. Chem. 2013, 288, 4502–4512. [Google Scholar] [CrossRef] [PubMed]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Shu, A.; Song, W.; Shi, W.; Li, M.; Zhang, W.; Li, Z.; Liu, G.; Yuan, F.; Zhang, S. Long-term organic fertilizer substitution increases rice yield by improving soil properties and regulating soil bacteria. Geoderma 2021, 404, 115287. [Google Scholar] [CrossRef]
- Li, J.; Yang, Q.; Shi, Z.; Zang, Z.; Liu, X. Effects of deficit irrigation and organic fertilizer on yield, saponin and disease incidence in Panax notoginseng under shaded conditions. Agric. Water Manag. 2021, 256. [Google Scholar] [CrossRef]
- Wen, T.; Xie, P.; Penton, C.R.; Hale, L.; Thomashow, L.S.; Yang, S.; Ding, Z.; Su, Y.; Yuan, J.; Shen, Q. Specific metabolites drive the deterministic assembly of diseased rhizosphere microbiome through weakening microbial degradation of autotoxin. Microbiome 2022, 10, 177. [Google Scholar] [CrossRef]
- Meng, J.-F.; Ning, P.-F.; Xu, T.-F.; Zhang, Z.-W. Effect of rain-shelter cultivation of Vitis vinifera cv. Cabernet gernischet on the phenolic profile of berry skins and the incidence of grape diseases. Molecules 2012, 18, 381–397. [Google Scholar] [CrossRef]
- Sui, Y.; Zhao, Q.; Wang, Z.; Liu, J.; Jiang, M.; Yue, J.; Lan, J.; Liu, J.; Liao, Q.; Wang, Q. A Comparative Analysis of the Microbiome of Kiwifruit at Harvest under Open-Field and Rain-Shelter Cultivation Systems. Front. Microbiol. 2021, 12, 2878. [Google Scholar] [CrossRef]
- Ryti, R. On the determination of soil pH. Agric. Food Sci. 1965, 37, 51–60. [Google Scholar] [CrossRef]
- Brooks, P.; Stark, J.M.; McInteer, B.; Preston, T. Diffusion method to prepare soil extracts for automated nitrogen-15 analysis. Soil Sci. Soc. Am. J. 1989, 53, 1707–1711. [Google Scholar] [CrossRef]
- Watanabe, F.; Olsen, S. Test of an ascorbic acid method for determining phosphorus in water and NaHCO3 extracts from soil. Soil Sci. Soc. Am. J. 1965, 29, 677–678. [Google Scholar] [CrossRef]
- Fang, Z.; Harris, J.M.; Ruzicka, J.; Hansen, E.H. Simultaneous flame photometric determination of lithium, sodium, potassium, and calcium by flow injection analysis with gradient scanning standard addition. Anal. Chem. 1985, 57, 1457–1461. [Google Scholar] [CrossRef]
- Han, X.; Cheng, Z.; Meng, H. Soil properties, nutrient dynamics, and soil enzyme activities associated with garlic stalk decomposition under various conditions. PLoS ONE 2012, 7, e50868. [Google Scholar] [CrossRef] [PubMed]
- Lebeis, S.L.; Paredes, S.H.; Lundberg, D.S.; Breakfield, N.; Gehring, J.; McDonald, M.; Malfatti, S.; Glavina del Rio, T.; Jones, C.D.; Tringe, S.G. Salicylic acid modulates colonization of the root microbiome by specific bacterial taxa. Science 2015, 349, 860–864. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Wen, T.; Yu, G.-H.; Hong, W.-D.; Yuan, J.; Niu, G.-Q.; Xie, P.-H.; Sun, F.-S.; Guo, L.-D.; Kuzyakov, Y.; Shen, Q.-R. Root exudate chemistry affects soil carbon mobilization via microbial community reassembly. Fundam. Res. 2022, 2, 697–707. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, T.; Ye, C.; Lu, R.; Liu, Y.; Huang, H.; He, X.; Yang, M.; Zhu, S. Leaching alleviates phenol-mediated root rot in Panax notoginseng by modifying the soil microbiota. Plant Soil 2021, 468, 491–507. [Google Scholar] [CrossRef]
- Brockett, B.F.; Prescott, C.E.; Grayston, S.J. Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada. Soil Biol. Biochem. 2012, 44, 9–20. [Google Scholar] [CrossRef]
- Frindte, K.; Pape, R.; Werner, K.; Löffler, J.; Knief, C. Temperature and soil moisture control microbial community composition in an arctic–alpine ecosystem along elevational and micro-topographic gradients. ISME J. 2019, 13, 2031–2043. [Google Scholar] [CrossRef]
- Yao, M.; Rui, J.; Niu, H.; Heděnec, P.; Li, J.; He, Z.; Wang, J.; Cao, W.; Li, X. The differentiation of soil bacterial communities along a precipitation and temperature gradient in the eastern Inner Mongolia steppe. Catena 2017, 152, 47–56. [Google Scholar] [CrossRef]
- Wang, X.; Song, D.; Liang, G.; Zhang, Q.; Ai, C.; Zhou, W. Maize biochar addition rate influences soil enzyme activity and microbial community composition in a fluvo-aquic soil. Appl. Soil Ecol. 2015, 96, 265–272. [Google Scholar] [CrossRef]
- Woo, H.L.; Hazen, T.C.; Simmons, B.A.; DeAngelis, K.M. Enzyme activities of aerobic lignocellulolytic bacteria isolated from wet tropical forest soils. Syst. Appl. Microbiol. 2014, 37, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Li, P.; Sayer, E.J.; Zhang, B.; Wang, J.; Qiao, C.; Peng, Z.; Diao, L.; Chi, Y.; Liu, W. Initial soil organic matter content influences the storage and turnover of litter, root and soil carbon in grasslands. Ecosystems 2018, 21, 1377–1389. [Google Scholar] [CrossRef]
- Li, P.; Zhang, X.; Hao, M. Effects of vegetation restoration on soil physicochemical properties, enzyme activities, and fungal community of reconstructed soil in a mining area on Loess Plateau. Bull. Soil Water Conserv. 2019, 11, 2295. [Google Scholar]
- Liu, S.; Wang, Z.; Niu, J.; Dang, K.; Zhang, S.; Wang, S.; Wang, Z. Changes in physicochemical properties, enzymatic activities, and the microbial community of soil significantly influence the continuous cropping of Panax quinquefolius L. (American ginseng). Plant Soil 2021, 463, 427–446. [Google Scholar] [CrossRef]
- Zhang, J.; Wei, L.; Yang, J.; Ahmed, W.; Wang, Y.; Fu, L.; Ji, G. Probiotic consortia: Reshaping the rhizospheric microbiome and its role in suppressing root-rot disease of Panax notoginseng. Front. Microbiol. 2020, 11, 701. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Jousset, A.; Guo, S.; Karlsson, I.; Zhao, Q.; Wu, H.; Kowalchuk, G.A.; Shen, Q.; Li, R.; Geisen, S. Soil protist communities form a dynamic hub in the soil microbiome. ISME J. 2018, 12, 634–638. [Google Scholar] [CrossRef]
- Getzke, F.; Thiergart, T.; Hacquard, S. Contribution of bacterial-fungal balance to plant and animal health. Curr. Opin. Microbiol. 2019, 49, 66–72. [Google Scholar] [CrossRef]
- Gu, Y.; Banerjee, S.; Dini-Andreote, F.; Xu, Y.; Shen, Q.; Jousset, A.; Wei, Z. Small changes in rhizosphere microbiome composition predict disease outcomes earlier than pathogen density variations. ISME J. 2022, 16, 2448–2456. [Google Scholar] [CrossRef]
- Crits-Christoph, A.; Diamond, S.; Butterfield, C.N.; Thomas, B.C.; Banfield, J.F. Novel soil bacteria possess diverse genes for secondary metabolite biosynthesis. Nature 2018, 558, 440–444. [Google Scholar] [CrossRef]
- Mendes, R.; Kruijt, M.; De Bruijn, I.; Dekkers, E.; Van Der Voort, M.; Schneider, J.H.; Piceno, Y.M.; DeSantis, T.Z.; Andersen, G.L.; Bakker, P.A. Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, N.; Tiedje, J.M.; Quensen III, J.F.; Meng, Q.; Hao, J.J. Microbial communities associated with potato common scab-suppressive soil determined by pyrosequencing analyses. Plant Dis. 2012, 96, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Wang, Z.; Li, N.; Wang, Y.; Feng, J.; Zhang, X. Bacillus amyloliquefaciens B1408 suppresses Fusarium wilt in cucumber by regulating the rhizosphere microbial community. Appl. Soil Ecol. 2019, 136, 55–66. [Google Scholar] [CrossRef]
- Cha, J.-Y.; Han, S.; Hong, H.-J.; Cho, H.; Kim, D.; Kwon, Y.; Kwon, S.-K.; Crüsemann, M.; Bok Lee, Y.; Kim, J.F. Microbial and biochemical basis of a Fusarium wilt-suppressive soil. ISME J. 2016, 10, 119–129. [Google Scholar] [CrossRef]
- Goh, Y.K.; Zoqratt, M.Z.H.M.; Goh, Y.K.; Ayub, Q.; Ting, A.S.Y. Discovering naturally-occurring microbiota in disease suppressive soil: Potential role of biological elements in suppressing Ganoderma boninense. Biol. Control 2022, 165, 104787. [Google Scholar] [CrossRef]
- Kappell, A.D.; Wei, Y.; Newton, R.J.; Van Nostrand, J.D.; Zhou, J.; McLellan, S.L.; Hristova, K.R. The polycyclic aromatic hydrocarbon degradation potential of Gulf of Mexico native coastal microbial communities after the Deepwater Horizon oil spill. Front. Microbiol. 2014, 5, 205. [Google Scholar] [CrossRef]
- Liu, J.; Li, Y.; Jiang, G.; Bian, X.; Li, F.; Geng, W. Allelopathic effects of cotton in continuous cropping. Allelopath. J. 2008, 21, 299–305. [Google Scholar]
- Liu, L.; Liu, D.; Jin, H.; Feng, G.; Zhang, J.; Wei, M.; Zhao, Z. Overview on the mechanisms and control methods of sequential cropping obstacle of Panax notoginseng FH Chen. J Mt. Agric. Biol 2011, 30, 70–75. [Google Scholar]
- Wu, Z.; Hao, Z.; Sun, Y.; Guo, L.; Huang, L.; Zeng, Y.; Wang, Y.; Yang, L.; Chen, B. Comparison on the structure and function of the rhizosphere microbial community between healthy and root-rot Panax notoginseng. Appl. Soil Ecol. 2016, 107, 99–107. [Google Scholar] [CrossRef]
- Degryse, F.; Verma, V.; Smolders, E. Mobilization of Cu and Zn by root exudates of dicotyledonous plants in resin-buffered solutions and in soil. Plant Soil 2008, 306, 69–84. [Google Scholar] [CrossRef]
- He, D.; Shen, W.; Eberwein, J.; Zhao, Q.; Ren, L.; Wu, Q.L. Diversity and co-occurrence network of soil fungi are more responsive than those of bacteria to shifts in precipitation seasonality in a subtropical forest. Soil Biol. Biochem. 2017, 115, 499–510. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Irrigation Regimes | Irrigation (mm) | Fertilizer (kg ha−1) | Rainfall (mm) | Drainage (mm) | Total Water Volume (mm) | |
|---|---|---|---|---|---|---|
| Field experiment | DW | 512 | 0 | 0 | 0 | 512 |
| DWF | 512 | 1680 | 0 | 0 | 512 | |
| RW | 424 | 0 | 856 | 768 | 512 | |
| RWF | 424 | 1680 | 856 | 768 | 512 | |
| Pot experiment | DW DWF | 672 | 0 | 0 | 0 | 672 |
| 672 | 1680 | 0 | 0 | 672 | ||
| RW | 432 | 0 | 856 | 613 | 672 | |
| RWF | 432 | 1680 | 856 | 613 | 672 | |
| Treatments | Soil pH | Available Potassium | Alkali-Hydrolyzed Nitrogen | Available Phosphorus | Soil Temperature | Soil Moisture | |
|---|---|---|---|---|---|---|---|
| mg·kg−1 | mg·kg−1 | mg·kg−1 | °C | % | |||
| Field experiment | DW | 6.61 ± 0.1 bb | 158.64 ± 16.11 ab | 20.2 ± 0.26 b | 25.25 ± 0.88 b | 26.01 ± 0.12 b | 24.16 ± 0.35 b |
| DWF | 6.83 ± 0.05 a | 186.29 ± 21.32 a | 22.4 ± 0.23 a | 29.45 ± 0.56 a | 26.28 ± 0.1 a | 23.6 ± 0.22 b | |
| RW | 6.49 ± 0.05 c | 129.32 ± 8.88 b | 17.8 ± 0.25 c | 23.93 ± 0.79 c | 24.19 ± 0.11 d | 30.25 ± 0.33 a | |
| RWF | 6.67 ± 0.06 b | 135.56 ± 10.84 b | 19.2 ± 0.31 b | 24.48 ± 1.12 b | 24.69 ± 0.13 c | 30.41 ± 0.26 a | |
| Pot experiment | DW | 7.86 ± 0.1 B | 198.36 ± 15.16 B | 30.41 ± 3.56 C | 59.07 ± 2.22 C | 24.86 ± 0.29 A | 33.15 ± 0.22 B |
| DWF | 8.02 ± 0.09 A | 287.34 ± 21.32 A | 52.74 ± 1.56 A | 81.32 ± 3.56 A | 25.3 ± 0.26 A | 32.10 ± 0.11 C | |
| RW | 7.76 ± 0.12 B | 119.26 ± 8.68 D | 25.09 ± 1.23 D | 50.4 ± 1.79 D | 23.74 ± 0.33 B | 42.76 ± 0.56 A | |
| RWF | 7.72 ± 0.07 B | 148.92 ± 10.13 C | 41.21 ± 2.12 B | 66.71 ± 2.12 B | 23.65 ± 0.11 B | 42.43 ± 0.63 A |
| Treatments | Shannon | Chao1 |
|---|---|---|
| Control | 6.09 ± 0.02 C | 3036 ± 50 C |
| DW | 6.50 ± 0.09 A | 3459 ± 70 A |
| DWF | 6.35 ± 0.03 B | 3284 ± 85 B |
| RW | 5.96 ± 0.04 D | 3296 ± 77 B |
| RWF | 6.23 ± 0.06 B | 2709 ± 62 D |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Liang, J.; Tang, Z.; Yang, Q. Rain Shelter Cultivation Reduces Root Rot Incidence of Panax notoginseng by Altering Root Exudates and Bacterial Communities under Micro-Irrigation and Fertilization. Agronomy 2023, 13, 1257. https://doi.org/10.3390/agronomy13051257
Zhang Y, Liang J, Tang Z, Yang Q. Rain Shelter Cultivation Reduces Root Rot Incidence of Panax notoginseng by Altering Root Exudates and Bacterial Communities under Micro-Irrigation and Fertilization. Agronomy. 2023; 13(5):1257. https://doi.org/10.3390/agronomy13051257
Chicago/Turabian StyleZhang, Yan, Jiaping Liang, Zhenya Tang, and Qiliang Yang. 2023. "Rain Shelter Cultivation Reduces Root Rot Incidence of Panax notoginseng by Altering Root Exudates and Bacterial Communities under Micro-Irrigation and Fertilization" Agronomy 13, no. 5: 1257. https://doi.org/10.3390/agronomy13051257