
Seed Dormancy and Germination Requirements of Torilis scabra (Apiaceae)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Seed Collection
2.2. Seed Morphology and Mass
2.3. Imbibition Tests
2.4. General Procedures of the Germination Test
2.5. Germination and Embryo Length in Fresh Seeds
2.6. Effects of Temperature on Embryo Growth
2.7. Effects of Cold or Warm Stratification on Seed Germination
2.8. Effects of Warm plus Cold Stratification on Seed Germination
2.9. Effect of Dry Storage (After-Ripening) on Seed Germination
2.10. Effects of Gibberellin on Seed Germination
2.11. Germination Phenology and Embryo Growth
2.12. Statistical Analyses
3. Results
3.1. Seed Morphology and Mass
3.2. Imbibition Tests
3.3. Germination and Embryo Length of Fresh Seeds
3.4. Effects of Temperature on Embryo Growth
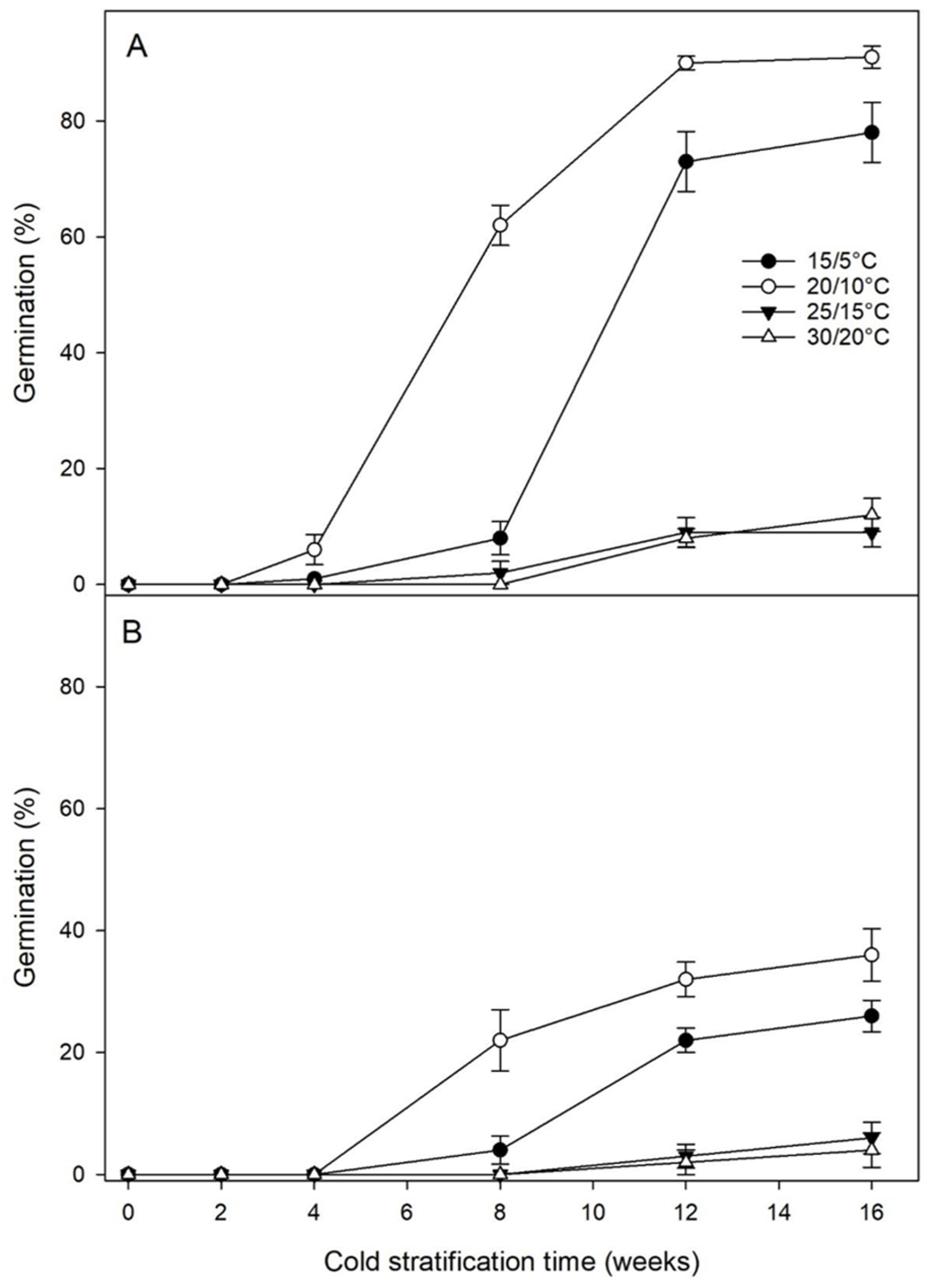
3.5. Effects of Cold or Warm Stratification on Seed Germination
3.6. Effects of Warm plus Cold Stratification on Seed Germination
3.7. Effects of Dry Storage (After-Ripening) on Seed Germination
3.8. Effects of Gibberellin on Seed Germination
3.9. Germination Phenology and Embryo Growth
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Penfield, S.; Springthorpe, V. Understanding chilling responses in Arabidopsis seeds and their contribution to life history. Philos. Trans. R. Soc. B-Biol. Sci. 2012, 367, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Springthorpe, V.; Penfield, S. Flowering time and seed dormancy control use external coincidence to generate life history strategy. eLife 2015, 4, e05557. [Google Scholar] [CrossRef] [PubMed]
- Rajapakshe, R.P.; Cross, A.T.; Turner, S.R.; Tomlinson, S. Understanding the interplay of temperature and moisture on the germination niche to improve management of threatened species impacted by mining. Restor. Ecol. 2022, 30, e13708. [Google Scholar] [CrossRef]
- Turcotte, M.M.; Levine, J.M. Phenotypic plasticity and species coexistence. Trends Ecol. Evol. 2016, 31, 803–813. [Google Scholar] [CrossRef] [PubMed]
- James, A.R. Inter-annual facilitation via pollinator support arises with species-specific germination rates in a model of plant–pollinator communities. Proc. R. Soc. B 2023, 290, 20221485. [Google Scholar] [CrossRef]
- Simons, A.M.; Johnston, M.O. Environmental and genetic sources of diversification in the timing of seed germination: Implications for the evolution of bet hedging. Evolution 2006, 60, 2280–2292. [Google Scholar] [CrossRef] [PubMed]
- Mondoni, A.; Rossi, G.; Orsenigo, S.; Probert, R.J. Climate warming could shift the timing of seed germination in alpine plants. Ann. Bot. 2012, 110, 155–164. [Google Scholar] [CrossRef]
- Tielbörger, K.; Petruů, M.; Lampei, C. Bet-hedging germination in annual plants: A sound empirical test of the theoretical foundations. Oikos 2012, 121, 1860–1868. [Google Scholar] [CrossRef]
- Lewandrowski, W.; Erickson, T.E.; Dalziell, E.L.; Stevens, J.C. Ecological niche and bet-hedging strategies for Triodia (R. Br.) seed germination. Ann. Bot. 2018, 121, 367–375. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Footitt, S. Seed dormancy cycling and the regulation of dormancy mechanisms to time germination in variable field environments. J. Exp. Bot. 2017, 68, 843–856. [Google Scholar] [CrossRef]
- Zhang, K.; Ji, Y.; Fu, G.; Yao, L.; Liu, H.; Tao, J.; Walck, J.L. Dormancy cycles in Aquilegia oxysepala Trautv. et Mey. (Ranunculaceae), a species with non-deep simple morphophysiological dormancy. Plant Soil 2021, 464, 223–235. [Google Scholar] [CrossRef]
- Zhang, K.; Ji, Y.; Song, X.; Yao, L.; Liu, H.; Tao, J. Deep complex morphophysiological dormancy in seeds of Clematis hexapetala Pall. (Ranunculaceae). Sci. Hortic. 2021, 286, 110247. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. The great diversity in kinds of seed dormancy: A revision of the Nikolaeva-Baskin classification system for primary seed dormancy. Seed Sci. Res. 2021, 31, 249–277. [Google Scholar] [CrossRef]
- Martin, A.C. The comparative internal morphology of seeds. Am. Midl. Nat. 2013, 36, 531–660. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: San Diego, CA, USA, 2014. [Google Scholar]
- Baskin, J.M.; Baskin, C.C. Seed germination ecology of poison hemlock, Conium maculatum. Can. J. Bot. 1990, 68, 2018–2024. [Google Scholar] [CrossRef]
- Rawnsley, R.P.; Lane, P.A.; Brown, P.H.; Groom, T. Occurrence and severity of the weeds Anthriscus caucalis and Torilis nodosa in pyrethrum. Aust. J. Exp. Agric. 2007, 46, 711–716. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef]
- Vandelook, F.; Bolle, N.; Van Assche, J.A. Seed Dormancy and germination of the European Chaerophyllum temulum (Apiaceae), a Member of a Trans-Atlantic Genus. Ann. Bot. 2007, 100, 233–239. [Google Scholar] [CrossRef]
- Vandelook, F.; Van de Moer, D.; Van Assche, J.A. Environmental signals for seed germination reflect habitat adaptations in four temperate Caryophyllaceae. Funct. Ecol. 2008, 22, 470–478. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M.; Chester, E.W. Ecological life cycle of Trepocarpus aethusae (Nutt.) ex DC. and comparisons with two other winter annual apiaceae native to Eastern United States. Castanea 2003, 68, 43–55. [Google Scholar]
- Nikolaeva, M.G.; Rasumova, M.V.; Gladkova, V.N. Reference Book on Dormant Seed Germination, 2nd ed.; “Nauka” Publishers (Leningrad Branch): Leningrad, Russia, 1985. [Google Scholar]
- Rosbakh, S.; Phartyal, S.S.; Poschlod, P. Seed germination traits shape community assembly along a hydroperiod gradient. Ann. Bot. 2020, 125, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Baskin, J.M.; Baskin, C.C. Nondeep complex morphophysiological dormancy in seeds of Osmorhiza claytonii (Apiaceae). Am. J. Bot. 1991, 78, 588–593. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. Germination ecophysiology of the woodland herb Osmorhiza longistylis (Umbelliferae). Am. J. Bot. 1984, 71, 687–692. [Google Scholar] [CrossRef]
- Hawkins, T.S.; Baskin, C.C.; Baskin, J.M. Morphophysiological dormancy in seeds of three eastern North American Sanicula species (Apiaceae subf. Saniculoideae): Evolutionary implications for dormancy break. Plant Species Biol. 2010, 25, 103–113. [Google Scholar] [CrossRef]
- Baker, K.S.; Steadman, K.J.; Plummer, J.A.; Merritt, D.J.; Dixon, K.W. The changing window of conditions that promotes germination of two fire ephemerals, Actinotus leucocephalus (Apiaceae) and Tersonia cyathiflora (Gyrostemonaceae). Ann. Bot. 2005, 96, 1225–1236. [Google Scholar] [CrossRef]
- Phartyal, S.S.; Kondo, T.; Baskin, J.M.; Baskin, C.C. Temperature requirements differ for the two stages of seed dormancy break in Aegopodium podagraria (Apiaceae), a species with deep complex morphophysiological dormancy. Ann. Bot. 2009, 96, 1086–1095. [Google Scholar] [CrossRef]
- Vandelook, F.; Lenaerts, J.; Jozef, A. The role of temperature in post-dispersal embryo growth and dormancy break in seeds of Aconitum lycoctonum L. Flora 2009, 204, 536–542. [Google Scholar] [CrossRef]
- Baskin, C.C.; Milberg, P.; Anderson, L.; Baskin, J.M. Deep complex morphophysiological dormancy in seeds of Anthriscus sylvestris (Apiaceae). Flora 2000, 195, 245–251. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. The ecological life cycle of Cryptotaenia canadensis (L.) DC. (Umbelliferae), a woodland herb with monocarpic ramets. Am. Midl. Nat. 1988, 119, 165–173. [Google Scholar] [CrossRef]
- Schutte, B.; Knee, M. The effects of rudimentary embryos and elevated oxygen on seed dormancy of Eryngium yuccifolium Michx. (Apiaceae). Seed Sci. Technol. 2005, 33, 53–62. [Google Scholar] [CrossRef]
- Nelson, J.R.; Wilson, A.M.; Goebel, C.J. Factors influencing broadcast seeding in bunchgrass range. Rangel. Ecol. Manag. 1970, 23, 163–170. [Google Scholar] [CrossRef]
- Brooks, S.; Cordell, S.; Perry, L. Broadcast seeding as a potential tool to reestablish native species in degraded dry forest ecosystems in Hawaii. Ecol. Restor. 2009, 27, 300–305. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Walck, J.L.; Jun, T. Non-deep simple morphophysiological dormancy in seeds of Angelica keiskei (Apiaceae). Sci. Hortic. 2019, 255, 202–208. [Google Scholar] [CrossRef]
- Yan, A.; Chen, Z. The control of seed dormancy and germination by temperature, light and nitrate. Bot. Rev. 2020, 86, 39–75. [Google Scholar] [CrossRef]
- Kawano, H.; Kanazawa, Y.; Suzuki, K.; Ohara, M. Seed germination characteristics of Maianthemum dilatatum (Wood) Nels. et Macbr. (Asparagaceae). Plant Species Biol. 2020, 35, 38–48. [Google Scholar] [CrossRef]
- Zhang, K.; Pan, H.; Baskin, C.C.; Baskin, J.M.; Xiong, Z.; Cao, W.; Yao, L.; Tang, B.; Zhang, C.; Tao, J. Epicotyl morphophysiological dormancy in seeds of Paeonia ostii (Paeoniaceae): Seasonal temperature regulation of germination phenology. Environ. Exp. Bot. 2022, 194, 104742. [Google Scholar] [CrossRef]
- Hidayati, S.N.; Walck, J.L.; Taylor, J. Seed germination ecophysiology of two western North American species of Sambucus (Caprifoliaceae sensu lato), and comparisons with eastern North American and European congeners. Plant Species Biol. 2010, 25, 203–213. [Google Scholar] [CrossRef]
- Tsang, A.C.W.; Corlett, R.T. Reproductive biology of the Ilex species (Aquifoliaceae) in Hong Kong, China. Can. J. Bot. 2005, 83, 1645–1654. [Google Scholar] [CrossRef]
- She, M.; Watson, M.F. Flora of China, 3rd ed.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2005; p. 28. [Google Scholar]
- Ye, J. Common Chinese Herbs in Daily Life, 2nd ed.; People’s Publishing House of Shandong: Jinan, China, 2009. [Google Scholar]
- Zhang, Y.; Zhang, X.; Guo, T.; Luo, X. Characteristics of temperature and precipitation change and the influence on climate productive potential in the Northwestern Liaoning. Chin. J. Agrometeorol. 2015, 36, 203. [Google Scholar]
- Nikolaeva, M.G. Factors controlling the seed dormancy pattern. In The Physiology and Biochemistry of Seed Dormancy and Germination, 2nd ed.; Khan, A.A., Ed.; North Holland Publishing Co.: Amsterdam, The Netherlands, 1977; pp. 51–74. [Google Scholar]
- Lee, S.Y.; Rhie, Y.H.; Kim, K.S. Non-deep simple morphophysiological dormancy in seeds of Thalictrum rochebrunianum, an endemic perennial herb in the Korean Peninsula. Hortic. Environ. Biotechnol. 2015, 56, 366–375. [Google Scholar] [CrossRef]
- Nurulla, M.; Baskin, C.C.; Lu, J.; Tan, D.; Baskin, J.M. Intermediate morphophysiological dormancy allows for life-cycle diversity in the annual weed, Turgenia latifolia (Apiaceae). Aust. J. Bot. 2014, 62, 630–637. [Google Scholar] [CrossRef]
- Kang, J.; Yim, S.; Choi, H.; Kim, A.; Lee, K.P.; Lopez-Molina, L.; Martinoia, E.; Lee, Y. Abscisic acid transporters cooperate to control seed germination. Nat. Commun. 2014, 6, 8113. [Google Scholar] [CrossRef] [PubMed]
- Geneve R, L. Impact of temperature on seed dormancy. HortScience 2003, 38, 336–340. [Google Scholar] [CrossRef]
- Walck, J.L.; Baskin, C.C.; Baskin, J.M. Seeds of Thalictrum mirabile (Ranunculaceae) require cold stratification for loss of nondeep simple morphophysiological dormancy. Can. J. Bot. 1999, 77, 1769–1776. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Xu, C.; Liu, H.; Tao, J.; Zhang, K. Seed Dormancy and Germination Requirements of Torilis scabra (Apiaceae). Agronomy 2023, 13, 1250. https://doi.org/10.3390/agronomy13051250
Zhang L, Xu C, Liu H, Tao J, Zhang K. Seed Dormancy and Germination Requirements of Torilis scabra (Apiaceae). Agronomy. 2023; 13(5):1250. https://doi.org/10.3390/agronomy13051250
Chicago/Turabian StyleZhang, Lei, Chaohan Xu, Huina Liu, Jun Tao, and Keliang Zhang. 2023. "Seed Dormancy and Germination Requirements of Torilis scabra (Apiaceae)" Agronomy 13, no. 5: 1250. https://doi.org/10.3390/agronomy13051250