
Screening Canola Genotypes for Resistance to Ammonium Toxicity


Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Design
2.1.1. Evaluating Growth Response of Canola to Soil Ammonium Levels
2.1.2. Screening Canola Genotypes for Resistance to Ammonium Toxicity
2.1.3. Data Collection
2.1.4. Soil pH
2.1.5. Soil Moisture
2.1.6. Residual Ammonium in Soil
2.1.7. Nitrogen Concentration in Roots
2.2. Statistical Analysis
3. Results
3.1. Experiment 1
3.1.1. Shoot Dry Weight
3.1.2. Root Dry Weight
3.1.3. Soil pHCaCl2
3.1.4. The Residual Ammonium in Soil (mg NH4+-N/kg Soil)
3.2. Experiment 2
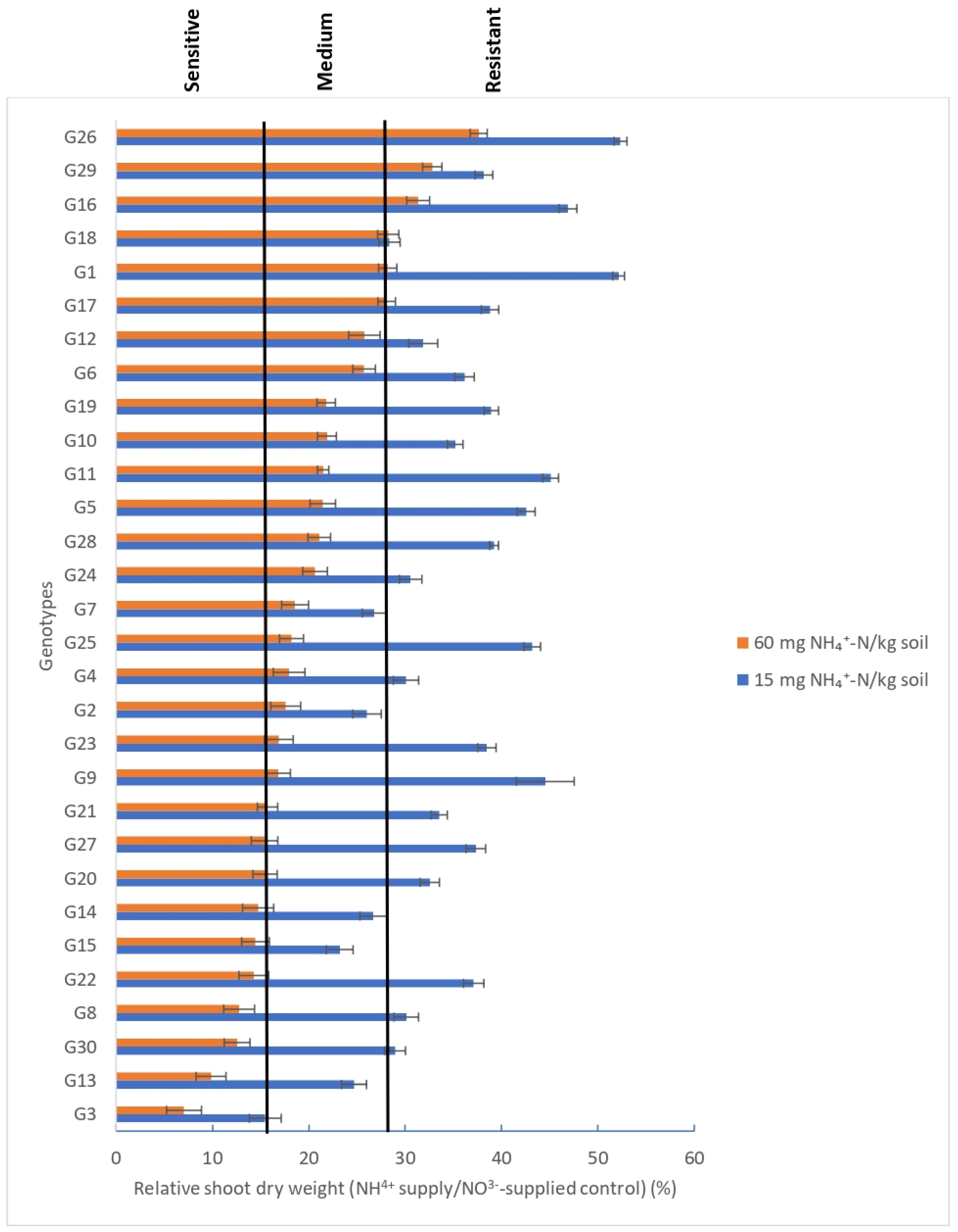
3.2.1. Shoot Dry Weight
3.2.2. Relative Root Dry Weight
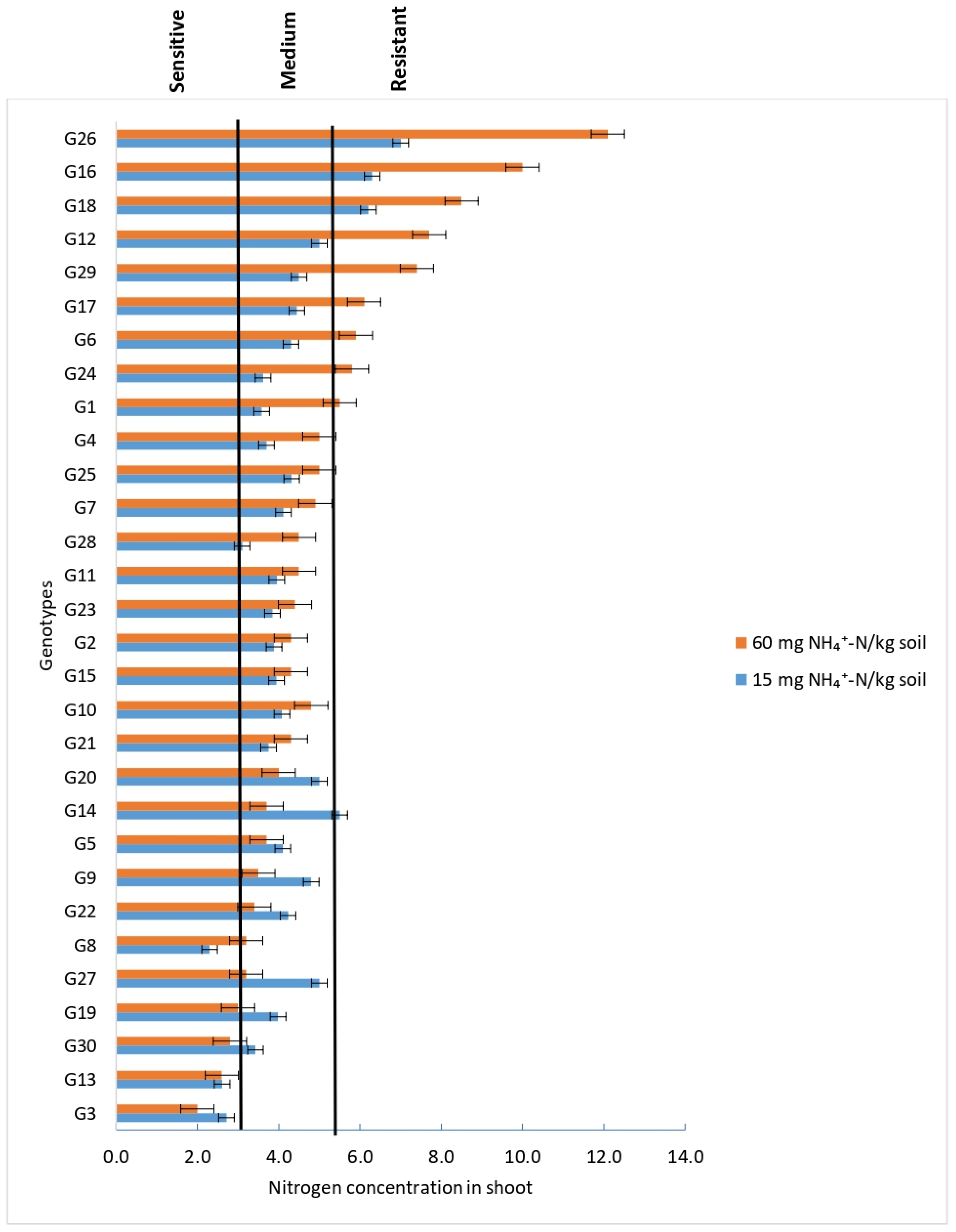
3.2.3. Nitrogen Concentration in Shoots
3.2.4. The Residual Ammonium in Soil (mg NH4+-N/kg Soil)
3.2.5. Soil pH
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Downey, R.K. Rapeseed to canola: Rags to riches. In Agricultural Biotechnology: Economic Growth through New Products, Partnerships and Workforce Development; Eaglesham, A., Hardy, R.W.F., Eds.; National Agricultural Biotechnology Council: Ithaca, NY, USA, 2006; pp. 67–76. [Google Scholar]
- Hickling, D.; Eskin, N.A.M.; Daun, J.K. Canola: Chemistry, Production, Processing, and Utilization; AOCS Press: Urbana, IL, USA, 2011. [Google Scholar]
- AOF. Australian Oilseeds Federation: Crop Report: Quality Standards, Technical Information and Typical Analysis, Issue 13; Australian Oilseed Federation: Mulwala, NSW, Australia, 2014. [Google Scholar]
- Seberry, D.E.; McCaffery, D.W.; Kingha, T.M. Quality of Australian Canola 2015–2016; Australian Oilseeds Federation, Department of Primary Industries NSW: Sydney, NSW, Australia, 2015; Volume 22, pp. 1–43. [Google Scholar]
- Ngezimana, W.; Agenbag, G.A. Effects of nitrogen and sulphur on canola (Brassica napus L.) vegetative and reproductive growth under controlled conditions. Afr. J. Agric. Res. 2013, 8, 4887–4894. [Google Scholar]
- Seymour, M. Canola Response to Split Nitrogen Applications at Eradu 2013; Department of Agriculture and Food Australia: Canberra, ACT, Australia, 2013. [Google Scholar]
- Ludewig, U.; Neuhäuser, B.; Dynowski, M. Molecular mechanisms of ammonium transport and accumulation in plants. FEBS Lett. 2007, 581, 2301–2308. [Google Scholar] [CrossRef] [PubMed]
- Zaman, M.; Blennerhassett, J.D. Effects of the different rates of urease and nitrification inhibitors on gaseous emissions of ammonia and nitrous oxide, nitrate leaching and pasture production from urine patches in an intensive grazed pasture system. Agric. Ecosyst. Environ. 2010, 136, 236–246. [Google Scholar] [CrossRef]
- Tong, D.; Xu, R. Effects of urea and (NH4+)2SO4 on nitrification and acidification of Ultisols from Southern China. J. Environ. Sci. 2012, 24, 682–689. [Google Scholar] [CrossRef] [PubMed]
- Angus, J.F.; Gupta, V.V.S.R.; Pitson, G.D.; Good, A.J. Effects of banded ammonia and urea fertiliser on soil properties and the growth and yield of wheat. Crop Pasture Sci. 2014, 65, 337–352. [Google Scholar] [CrossRef]
- Harris, R.H.; Armstrong, R.D.; Wallace, A.J.; Belyaeva, O.N. Effect of nitrogen fertiliser management on soil mineral nitrogen, nitrous oxide losses, yield and nitrogen uptake of wheat growing in waterlogging-prone soils of south-eastern Australia. Soil Res. 2016, 54, 619–633. [Google Scholar] [CrossRef]
- Sigurdarson, J.J.; Svane, S.; Karring, H. The molecular processes of urea hydrolysis in relation to ammonia emissions from agriculture. Rev. Environ. Sci. Bio/Technol. 2018, 17, 241–258. [Google Scholar] [CrossRef]
- Bremner, J. Problems in the use of urea as a nitrogen fertilizer. Soil Use Manag. 1990, 6, 70–71. [Google Scholar] [CrossRef]
- Owen, A.G.; Jones, D. Competition for amino acids between wheat roots and rhizosphere microorganisms and the role of amino acids in plant N acquisition. Soil Biol. Biochem. 2001, 33, 651–657. [Google Scholar] [CrossRef]
- Nadarajan, S.; Sukumaran, S. Chemistry and toxicology behind chemical fertilizers. In Controlled Release Fertilizers for Sustainable Agriculture; Elsevier: Amsterdam, The Netherlands, 2021; pp. 195–229. [Google Scholar]
- Walch-Liu, P.; Neumann, G.; Engels, C. Response of shoot and root growth to supply of different nitrogen forms is not related to carbohydrate and nitrogen status of tobacco plants. J. Plant Nutr. Soil Sci. 2001, 164, 97–103. [Google Scholar] [CrossRef]
- Britto, D.T.; Kronzucker, H.J. NH4+ toxicity in higher plants: A critical review. J. Plant Physiol. 2002, 159, 567–584. [Google Scholar] [CrossRef]
- Esteban, R.; Ariz, I.; Cruz, C.; Moran, J.F. Mechanisms of ammonium toxicity and the quest for tolerance. Plant Sci. 2016, 248, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Kronzucker, H.J.; Britto, D.T.; Davenport, R.J.; Tester, M. Ammonium toxicity and the real cost of transport. Trends Plant Sci. 2001, 6, 335–337. [Google Scholar] [CrossRef] [PubMed]
- Britto, D.T.; Siddiqi, M.Y.; Glass, A.D.M.; Kronzucker, H.J. Futile transmembrane NH4+ cycling: A cellular hypothesis to explain ammonium toxicity in plants. Proc. Natl. Acad. Sci. USA 2001, 98, 4255–4258. [Google Scholar] [CrossRef] [PubMed]
- Piwpuan, N.; Jampeetong, A.; Brix, H. Ammonium tolerance and toxicity of Actinoscirpus grossus—A candidate species for use in tropical constructed wetland systems. Ecotoxicol. Environ. Saf. 2014, 107, 319–328. [Google Scholar] [CrossRef]
- Bodirsky, B.L.; Popp, A.; Lotze-Campen, H.; Dietrich, J.P.; Rolinski, S.; Weindl, I.; Schmitz, C.; Müller, C.; Bonsch, M.; Humpenöder, F. Reactive nitrogen requirements to feed the world in 2050 and potential to mitigate nitrogen pollution. Nat. Commun. 2014, 5, 3858. [Google Scholar] [CrossRef]
- Ravishankara, A.; Daniel, J.S.; Portmann, R.W. Nitrous oxide (N2O): The dominant ozone-depleting substance emitted in the 21st century. Science 2009, 326, 123–125. [Google Scholar] [CrossRef]
- Park, S.; Croteau, P.; Boering, K.A.; Etheridge, D.M.; Ferretti, D.; Fraser, P.J.; Kim, K.R.; Krummel, P.B.; Langenfelds, R.L.; van Ommen, T.D.; et al. Trends and seasonal cycles in the isotopic composition of nitrous oxide since 1940. Nat. Geosci. 2012, 5, 261–265. [Google Scholar] [CrossRef]
- Lea, P.J.; Miflin, B.J. Nitrogen assimilation and its relevance to crop improvement. Annu. Plant Rev. 2010, 42, 1–40. [Google Scholar]
- Rahman, N.; Forrestal, P.J. Ammonium Fertilizer Reduces Nitrous Oxide Emission Compared to Nitrate Fertilizer While Yielding Equally in a Temperate Grassland. Agriculture 2021, 11, 1141. [Google Scholar] [CrossRef]
- Wu, S.-F.; Wu, L.-H.; Shi, Q.-W.; Wang, Z.-Q.; Chen, X.-Y.; Li, Y.-S. Effects of a new nitrification inhibitor 3,4-dimethylpyrazole phosphate (DMPP) on nitrate and potassium leaching in two soils. J. Environ. Sci. 2007, 19, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Cahalan, E.; Ernfors, M.; Müller, C.; Devaney, D.; Laughlin, R.J.; Watson, C.J.; Hennessy, D.; Grant, J.; Khalil, M.I.; McGeough, K.L.; et al. The effect of the nitrification inhibitor dicyandiamide (DCD) on nitrous oxide and methane emissions after cattle slurry application to Irish grassland. Agric. Ecosyst. Environ. 2015, 199, 339–349. [Google Scholar] [CrossRef]
- Guo, Y.J.; Di, H.J.; Cameron, K.C.; Li, B. Effect of application rate of a nitrification inhibitor, dicyandiamide (DCD), on nitrification rate, and ammonia-oxidizing bacteria and archaea growth in a grazed pasture soil: An incubation study. J. Soils Sediments 2014, 14, 897–903. [Google Scholar] [CrossRef]
- Ning, J.; Ai, S.; Cui, L. Dicyandiamide has more inhibitory activities on nitrification than thiosulfate. PLoS ONE 2018, 13, e0200598. [Google Scholar] [CrossRef]
- Cui, L.; Li, D.; Wu, Z.; Xue, Y.; Xiao, F.; Zhang, L.; Song, Y.; Li, Y.; Zheng, Y.; Zhang, J. Effects of nitrification inhibitors on soil nitrification and ammonia volatilization in three soils with different pH. Agronomy 2021, 11, 1674. [Google Scholar] [CrossRef]
- Saric, M.R.; Loughman, B.C. Genetic Aspects of Plant Nutrition. In Proceedings of the First International Symposium on Genetic Aspects of Plant Nutrition, Organized by the Serbian Academy of Sciences and Arts, Belgrade, Serbia, 30 August–4 September 1982; Springer: Dordrecht, The Netherlands, 2012. [Google Scholar]
- Siddiqui, M.H.; Mohammad, F.; Khan, M.N.; Al-Whaibi, M.H.; Bahkali, A.H. Nitrogen in relation to photosynthetic capacity and accumulation of osmoprotectant and nutrients in Brassica genotypes grown under salt stress. Agric. Sci. China 2010, 9, 671–680. [Google Scholar] [CrossRef]
- Zhong, Y.; Yan, W.; Chen, J.; Shangguan, Z. Net ammonium and nitrate fluxes in wheat roots under different environmental conditions as assessed by scanning ion-selective electrode technique. Sci. Rep. 2014, 4, 7223. [Google Scholar] [CrossRef]
- Wang, F.; Gao, J.; Liu, Y.; Tian, Z.; Muhammad, A.; Zhang, Y.; Jiang, D.; Cao, W.; Dai, T. Higher ammonium transamination capacity can alleviate glutamate inhibition on winter wheat (Triticum aestivum L.) root growth under high ammonium stress. PLoS ONE 2016, 11, e0160997. [Google Scholar] [CrossRef]
- Gashaw, L.; Mugwira, L.M. Ammonium-N and nitrate-N effects on the growth and mineral compositions of triticale, wheat, and rye. Agron. J. 1981, 73, 47–51. [Google Scholar] [CrossRef]
- Muhlestein, D.J. Effects of High Ammonium/Nitrate Ratios on Nitrification and Growth of Wheat in Hydroponic Culture. Ph.D. Thesis, Utah State University, Ann Arbor, MI, USA, 2001. [Google Scholar]
- Schortemeyer, M.; Stamp, P.; Feil, B. Ammonium tolerance and carbohydrate status in maize cultivars. Ann. Bot. 1997, 79, 25–30. [Google Scholar] [CrossRef]
- Chen, G.; Guo, S.; Kronzucker, H.J.; Shi, W. Nitrogen use efficiency (NUE) in rice links to NH4+ toxicity and futile NH4+ cycling in roots. Plant Soil 2013, 369, 351–363. [Google Scholar] [CrossRef]
- Park, H.; Charles, A.; Stutte. Effects of ammonium, nitrate and urea on the growth of soybean plants different in phosphorus sensitivity. Korean Agrinurture Chem. Soc. 1973, 16, 118–127. [Google Scholar]
- Bloom, A.J.; John, F. The influence of ammonium and chloride on potassium and nitrate absorption by barley roots depends on time of exposure and cultivar. Plant Physiol. 1986, 81, 67–69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Rengel, Z. Gradients of pH, ammonium, and phosphorus between the fertiliser band and wheat roots. Aust. J. Agric. Res. 1999, 50, 365–374. [Google Scholar] [CrossRef]
- Balint, T.; Rengel, Z.; Allen, D. Australian canola germplasm differs in nitrogen and sulfur efficiency. Aust. J. Agric. Res. 2008, 59, 167–174. [Google Scholar] [CrossRef]
- Jackie, B.; Mark, S.; Bob, F.; Raj, M.; Sally, S.; Martin, H.; Vanessa, S. Canola Variety Guide for Western Australia; Western Australia: Nedlands, Australia, 2015. [Google Scholar]
- Svečnjak, Z.; Rengel, Z. Nitrogen utilization efficiency in canola cultivars at grain harvest. Plant Soil 2006, 283, 299–307. [Google Scholar] [CrossRef]
- Rose, T.J.; Rengel, Z.; Ma, Q.; Bowden, J.W. Differential accumulation patterns of phosphorus and potassium by canola cultivars compared to wheat. J. Plant Nutr. Soil Sci. 2007, 170, 404–411. [Google Scholar] [CrossRef]
- Singh, B.; Rengel, Z.; Bowden, J.W. Carbon, nitrogen and sulphur cycling following incorporation of canola residue of different sizes into a nutrient-poor sandy soil. Soil Biol. Biochem. 2006, 38, 32–42. [Google Scholar] [CrossRef]
- Rayment, G.E.; Lyons, D.J. Soil Chemical Methods: Australasia; CSIRO Publishing: Collingwood, Australia, 2011; Volume 3. [Google Scholar]
- Searle, P.L. The Berthelot or indophenol reaction and its use in the analytical chemistry of nitrogen. Analyst 1984, 109, 549–568. [Google Scholar] [CrossRef]
- McGill, W.B.; Figueiredo, C.T. Total nitrogen. In Soil Sampling and Methods of Analysis; Carter, M.R., Ed.; Canadian Society of Soil Science and Lewis Publishers—CRC Press: Boca Raton, FL, USA, 1993; pp. 201–211. [Google Scholar]
- Rengel, Z.; Graham, R.D. Importance of seed Zn content for wheat growth on Zn-deficient soil. Plant Soil 1995, 173, 259–266. [Google Scholar] [CrossRef]
- Yau, S.K.; Thurling, N. Variation in nitrogen response among spring rape (Brassica napus) cultivars and its relationship to nitrogen uptake and utilization. Field Crops Res. 1987, 16, 139–155. [Google Scholar] [CrossRef]
- Tabatabaei, S.J.; Bybordi, A. Effect of different ratios of ammonium: Nitrate on photosynthesis and fatty acid composition in canola (Brassica napus L.) under saline conditions. J. Crop Prod. Process. 2012, 2, 83–92. [Google Scholar]
- Cruz, C.; Bio, A.F.M.; Domínguez-Valdivia, M.D.; Aparicio-Tejo, P.M.; Lamsfus, C.; Martins-Loução, M.A. How does glutamine synthetase activity determine plant tolerance to ammonium? Planta 2006, 223, 1068–1080. [Google Scholar] [CrossRef] [PubMed]
- Dowling, C.W. Seed and Seedling Tolerance of Cereal, Oilseed, Fibre and Legume Crops to Injury from Banded Ammonium Fertilizers. Ph.D. Thesis, Griffith University, Brisbane, QLD, Australia, 1998. [Google Scholar]
- Lasa, B.; Frechilla, S.; Lamsfus, C.; Aparicio-Tejo, P.M. The sensitivity to ammonium nutrition is related to nitrogen accumulation. Sci. Hortic. 2001, 91, 143–152. [Google Scholar] [CrossRef]
- Harada, T.; Takaki, H.; Yamada, Y. Effect of nitrogen sources on the chemical components in young plants. Soil Sci. Plant Nutr. 1968, 14, 47–55. [Google Scholar] [CrossRef]
- Malhi, S.S.; Nyborg, M.; Jahn, H.G.; Penney, D.C. Yield and nitrogen uptake of rapessed (Brassica campestris L.) with ammonium and nitrate. Plant Soil 1988, 105, 231–239. [Google Scholar] [CrossRef]
- Heuermann, D.; Hahn, H.; Von Wirén, N. Seed yield and nitrogen efficiency in oilseed rape after ammonium nitrate or urea fertilization. Front. Plant Sci. 2021, 11, 608785. [Google Scholar] [CrossRef]
- Gerwing, J.; Gelderman, R.; Bly, A. Influence of seed placed fertilizer on corn, soybean and sunflower emergence. In Proceedings of the 24th North Central Extension—Industry Soil Fertility Conference, St Louis, MO, USA, 26–27 October 1994; Potash & Phosphate Institute: Manhattan, KS, USA, 1994; pp. 26–27. [Google Scholar]
- Bremner, J.M.; Krogmeier, M.J. Elimination of the adverse effects of urea fertilizer on seed germination, seedling growth, and early plant growth in soil. Proc. Natl. Acad. Sci. USA 1988, 85, 4601–4604. [Google Scholar] [CrossRef]
- Claasen, M.E.T.; Wilcox, G.E. Effect of nitrogen form on growth and composition of tomato and pea tissue. J. Am. Soc. Hortic. Sci. 1974, 99, 171–174. [Google Scholar] [CrossRef]
- Magalhaes, J.R.; Huber, D.M. Response of ammonium assimilation enzymes to nitrogen form treatments in different plant species. J. Plant Nutr. 1991, 14, 175–185. [Google Scholar] [CrossRef]
- Hageman, R.H. Ammonium and nitrate nutrition of higher plants. Am. Soc. Agron. 1980, 67–85. [Google Scholar] [CrossRef]
- Horchani, F.; Hajri, R.; Aschi-Smiti, S. Effect of ammonium or nitrate nutrition on photosynthesis, growth, and nitrogen assimilation in tomato plants. J. Plant Nutr. Soil Sci. 2010, 173, 610–617. [Google Scholar] [CrossRef]
- Roosta, H.R.; Schjoerring, J.K. Effects of ammonium toxicity on nitrogen metabolism and elemental profile of cucumber plants. J. Plant Nutr. 2007, 30, 1933–1951. [Google Scholar] [CrossRef]
- Bittsánszky, A.; Pilinszky, K.; Gyulai, G.; Komives, T. Overcoming ammonium toxicity. Plant Sci. 2015, 231, 184–190. [Google Scholar] [CrossRef]
- Matt, P.; Geiger, M.; Walch-Liu, P.; Engels, C.; Krapp, A.; Stitt, M. The immediate cause of the diurnal changes of nitrogen metabolism in leaves of nitrate-replete tobacco: A major imbalance between the rate of nitrate reduction and the rates of nitrate uptake and ammonium metabolism during the first part of the light period. Plant Cell Environ. 2001, 24, 177–190. [Google Scholar]
- Schjoerring, J.K.; Husted, S.; Mäck, G.; Mattsson, M. The regulation of ammonium translocation in plants. J. Exp. Bot. 2002, 53, 883–890. [Google Scholar] [CrossRef]
- El Omari, R.; Rueda-López, M.; Avila, C.; Crespillo, R.; Nhiri, M.; Cánovas, F.M. Ammonium tolerance and the regulation of two cytosolic glutamine synthetases in the roots of sorghum. Funct. Plant Biol. 2010, 37, 55–63. [Google Scholar] [CrossRef]
- Cruz, C.; Domínguez-Valdivia, M.D.; Aparicio-Tejo, P.M.; Lamsfus, C.; Bio, A.; Martins-Loução, M.A.; Moran, J.F. Intra-specific variation in pea responses to ammonium nutrition leads to different degrees of tolerance. Environ. Exp. Bot. 2011, 70, 233–243. [Google Scholar] [CrossRef]
- Schortemeyer, M.; Feil, B.; Stamp, P. Root morphology and nitrogen uptake of maize simultaneously supplied with ammonium and nitrate in a split-root system. Ann. Bot. 1993, 72, 107–115. [Google Scholar] [CrossRef]
- Schortemeyer, M.; Feil, B. Root morphology of maize under homogeneous or spatially separated supply of ammonium and nitrate at three concentration ratios. J. Plant Nutr. 1996, 19, 1089–1097. [Google Scholar] [CrossRef]
- Chaillou, S.; Rideout, J.; Raper, C.J.; Morot-Gaudry, J.-F. Responses to ammonium and nitrate supplied in combination to the whole root system or separately in a split-root system. Physiol. Plant. 1994, 90, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Feil, B. Growth and ammonium:nitrate uptake ratio of spring wheat cultivars under a homogeneous and a spatially separated supply of ammonium and nitrate. J. Plant Nutr. 1994, 17, 717–728. [Google Scholar] [CrossRef]
- Hachiya, T.; Sakakibara, H. Interactions between nitrate and ammonium in their uptake, allocation, assimilation, and signaling in plants. J. Exp. Bot. 2016, 68, 2501–2512. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Di, T.; Zhu, Y.; Subbarao, G. Transcriptional response of plasma membrane H-ATPase genes to ammonium nutrition and its functional link to the release of biological nitrification inhibitors from sorghum roots. Plant Soil 2016, 398, 301–312. [Google Scholar] [CrossRef]
- Rengel, Z. Handbook of Soil Acidity; CRC Press: New York, NY, USA, 2003. [Google Scholar]
- Sarasketa, A.; González-Moro, M.B.; González-Murua, C.; Marino, D. Nitrogen source and external medium pH interaction differentially affects root and shoot metabolism in Arabidopsis. Front. Plant Sci. 2016, 7, 29. [Google Scholar] [CrossRef] [PubMed]
- DPIRD. Critical Nutrient Levels for Canola in Western Australia; Department of Primary Industries and Regional Development, Western of Australia: Perth, WA, Australia, 2019. [Google Scholar]
- Openshaw, M.D. The Effect of Ammonia on Germination and Development of Seedlings in Soil. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 1970. [Google Scholar]
- Leidi, E.O.; Silberbush, M.; Lips, S.H. Wheat growth as affected by nitrogen type, pH and salinity. I. biomass production and mineral composition. J. Plant Nutr. 1991, 14, 235–246. [Google Scholar] [CrossRef]
- Schroder, J.L.; Zhang, H.; Girma, K.; Raun, W.R.; Penn, C.J.; Payton, M.E. Soil acidification from long-term use of nitrogen fertilizers on winter wheat. Soil Sci. Soc. Am. J. 2011, 75, 957–964. [Google Scholar] [CrossRef]
- Findenegg, G.R.; Beusichem, M.L.v.; Keltjens, W.G. Proton balance of plants: Physiological, agronomical and ecological implications. Neth. J. Agric. Sci. 1986, 34, 371–379. [Google Scholar] [CrossRef]
- Findenegg, G.R. A comparative study of ammonium toxicity at different constatn pH of the nutrient solution. Plant Soil 1987, 103, 239–243. [Google Scholar] [CrossRef]
- Barker, A.V.; Mills, H.A. Ammonium and nitrate nutrition of horticultural crops. Hortic. Rev. 1980, 2, 395–423. [Google Scholar]
- Li, S.-X.; Wang, Z.-H.; Stewart, B. Responses of crop plants to ammonium and nitrate N. Adv. Agron. 2013, 118, 205–397. [Google Scholar]
- Pan, W.; A Jackson, W.; H Moll, R. Nitrate uptake and partitioning by corn root systems: Differential effects of ammonium among genotypes and stages of root development. Plant Physiol. 1985, 77, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, H.; Wang, S.; Shi, L.; Xu, F.; Wang, C.; Cai, H.; Ding, G. The rapeseed genotypes with contrasting NUE response discrepantly to varied provision of ammonium and nitrate by regulating photosynthesis, root morphology, nutritional status, and oxidative stress response. Plant Physiol. Biochem. 2021, 166, 348–360. [Google Scholar] [CrossRef] [PubMed]
- Azimi Sooran, S.; Amirshekari, H.; Shirani Rad, A.H.; Mozaffari, J.; Fotokian, M.H. Evaluation of winter canola genotypes response to terminal drought stress under application of ammonium sulfate. J. Crops Improv. 2018, 20, 627–642. [Google Scholar] [CrossRef]
- Williams, S.T.; Vail, S.; Arcand, M.M. Nitrogen Use Efficiency in Parent vs. Hybrid Canola under Varying Nitrogen Availabilities. Plants 2021, 10, 2364. [Google Scholar] [CrossRef]
- Singh, B. Are nitrogen fertilizers deleterious to soil health? Agronomy 2018, 8, 48. [Google Scholar] [CrossRef]
- Dari, B.; Rogers, C.W.; Walsh, O.S. Understanding factors controlling ammonia volatilization from fertilizer nitrogen applications. Univ. Ida. Ext. Bul. 2019, 926, 1–4. [Google Scholar]
- Shi, Y.; Wang, J.; Le Roux, X.; Mu, C.; Ao, Y.; Gao, S.; Zhang, J.; Knops, J.M.H. Trade-offs and synergies between seed yield, forage yield, and N-related disservices for a semi-arid perennial grassland under different nitrogen fertilization strategies. Biol. Fertil. Soils 2019, 55, 497–509. [Google Scholar] [CrossRef]
- Klapwijk, C.J.; van Wijk, M.T.; Rosenstock, T.S.; van Asten, P.J.A.; Thornton, P.K.; Giller, K.E. Analysis of trade-offs in agricultural systems: Current status and way forward. Curr. Opin. Environ. Sustain. 2014, 6, 110–115. [Google Scholar] [CrossRef]
- Morsi, N.A.A.; Hashem, O.S.M.; El-Hady, M.A.A.; Abd-Elkrem, Y.M.; El-temsah, M.E.; Galal, E.G.; Gad, K.I.; Boudiar, R.; Silvar, C.; El-Hendawy, S.; et al. Assessing Drought Tolerance of Newly Developed Tissue-Cultured Canola Genotypes under Varying Irrigation Regimes. Agronomy 2023, 13, 836. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Property | Unit | Results |
|---|---|---|
| Depth | cm | 0–10 |
| Gravel | % | 5 |
| Texture | Sandy | |
| Ammonium nitrogen | mg/kg | 1 |
| Nitrate nitrogen | mg/kg | 2 |
| Phosphorus (Colwell method) | mg/kg | <2 |
| Potassium (Colwell method) | mg/kg | 30 |
| Sulphur | mg/kg | 2.1 |
| Organic carbon | g/kg | 5.8 |
| Conductivity (1:5 water) | dS/m | 0.02 |
| pHCaCl2 | 5.8 | |
| DTPA-extractable copper | mg/kg | 0.15 |
| DTPA-extractable iron | mg/kg | 17.25 |
| DTPA-extractable manganese | mg/kg | 1.35 |
| DTPA-extractable zinc | mg/kg | 0.19 |
| Genotype # * | Genotype Name | Origin Country |
|---|---|---|
| 1 | Karoo-057DH | Australia |
| 2 | Campino | Europe |
| 3 | Zhongshuang4B | China |
| 4 | Zhongyou821 | China |
| 5 | (SC09-1) | China |
| 6 | CN01-104-2 | China |
| 7 | HAU02 | China |
| 8 | HAU11 | China |
| 9 | GSL1 | India |
| 10 | CB telfer | Australia |
| 11 | ATR Stingray | Australia |
| 12 | AV-Garnet | Australia |
| 13 | (AV-Opal) | Australia |
| 14 | (AV-Ruby) | Australia |
| 15 | Tranby | Australia |
| 16 | ZY001 | China |
| 17 | AG-Outback | Australia |
| 18 | AG-Spectrum | Australia |
| 19 | CB-Argyle | Australia |
| 20 | CB-Tanami | Australia |
| 21 | CB-Trilogy | Australia |
| 22 | Ding474 | China |
| 23 | Charlton | Australia |
| 24 | Oscar | Australia |
| 25 | Purler | Australia |
| 26 | Tarcoola-22 | Australia |
| 27 | Skipton | Australia |
| 28 | Surpass400 | Australia |
| 29 | (SC01-3) | Australia |
| 30 | (SC03-1) | Australia |
| Parameters | NH4+-N Treatments | NO3−-N Treatments |
|---|---|---|
| Shoot dry weight | ** | ** |
| Root dry weight | ** | NS |
| mg NH4+-N/kg soil | ** | ** |
| mg NO3−-N/kg soil | NS | ** |
| Soil pH | ** | ** |
| Parameters | N Treatments | Genotypes | N Treatments × Genotypes |
|---|---|---|---|
| Shoot dry weight | ** | ** | ** |
| Root dry weight | ** | ** | ** |
| mg NH4+-N/kg soil | ** | ** | ** |
| mg NO3−-N/kg soil | NS | NS | NS |
| Nitrogen concentration in shoot | ** | ** | ** |
| Soil pH | ** | NS | NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Awad, O.A.S.; Prendergast, K.S.; Robson, A.; Rengel, Z. Screening Canola Genotypes for Resistance to Ammonium Toxicity. Agronomy 2023, 13, 1150. https://doi.org/10.3390/agronomy13041150
Al-Awad OAS, Prendergast KS, Robson A, Rengel Z. Screening Canola Genotypes for Resistance to Ammonium Toxicity. Agronomy. 2023; 13(4):1150. https://doi.org/10.3390/agronomy13041150
Chicago/Turabian StyleAl-Awad, Omar Ali Shaban, Kit Stasia Prendergast, Alan Robson, and Zed Rengel. 2023. "Screening Canola Genotypes for Resistance to Ammonium Toxicity" Agronomy 13, no. 4: 1150. https://doi.org/10.3390/agronomy13041150