

Potential Impacts of Clove Essential Oil Nanoemulsion as Bio Fungicides against Neoscytalidium Blight Disease of Carum carvi L.

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of the Pathogen
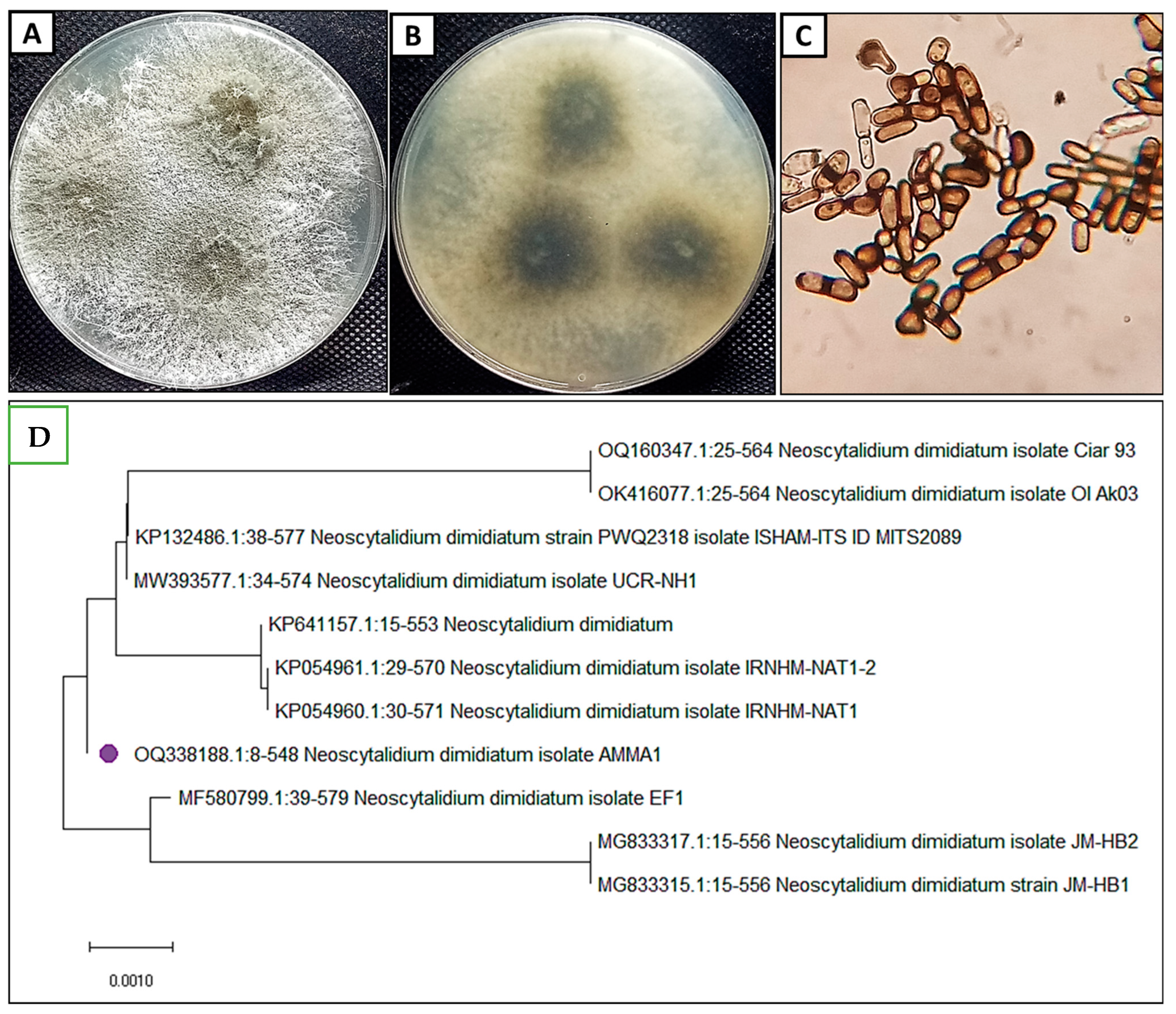
2.2. Identification of the Phytopathogenic Fungus
2.3. Preparation of CEONE
2.4. Characterization of CEONE
2.5. Antifungal Activity of CEONE
2.6. Pathogenicity Test
2.7. Pot Experiment
2.8. Morphological Indicators of Resistance
2.8.1. Disease Index
2.8.2. Plant Growth
2.8.3. Metabolic Indicators for C. carvi Resistance
2.9. Statistical Analysis
3. Results and Discussion
3.1. Morphological and Molecular Identification of the Isolate Pathogen
3.2. Characterization of CEONE
3.2.1. DLS and Zeta Potential
3.2.2. Transmission Electron Microscopy (TEM)
3.3. Antifungal Activity of CEONE
3.4. Disease Assessment
3.5. Vegetative Growth
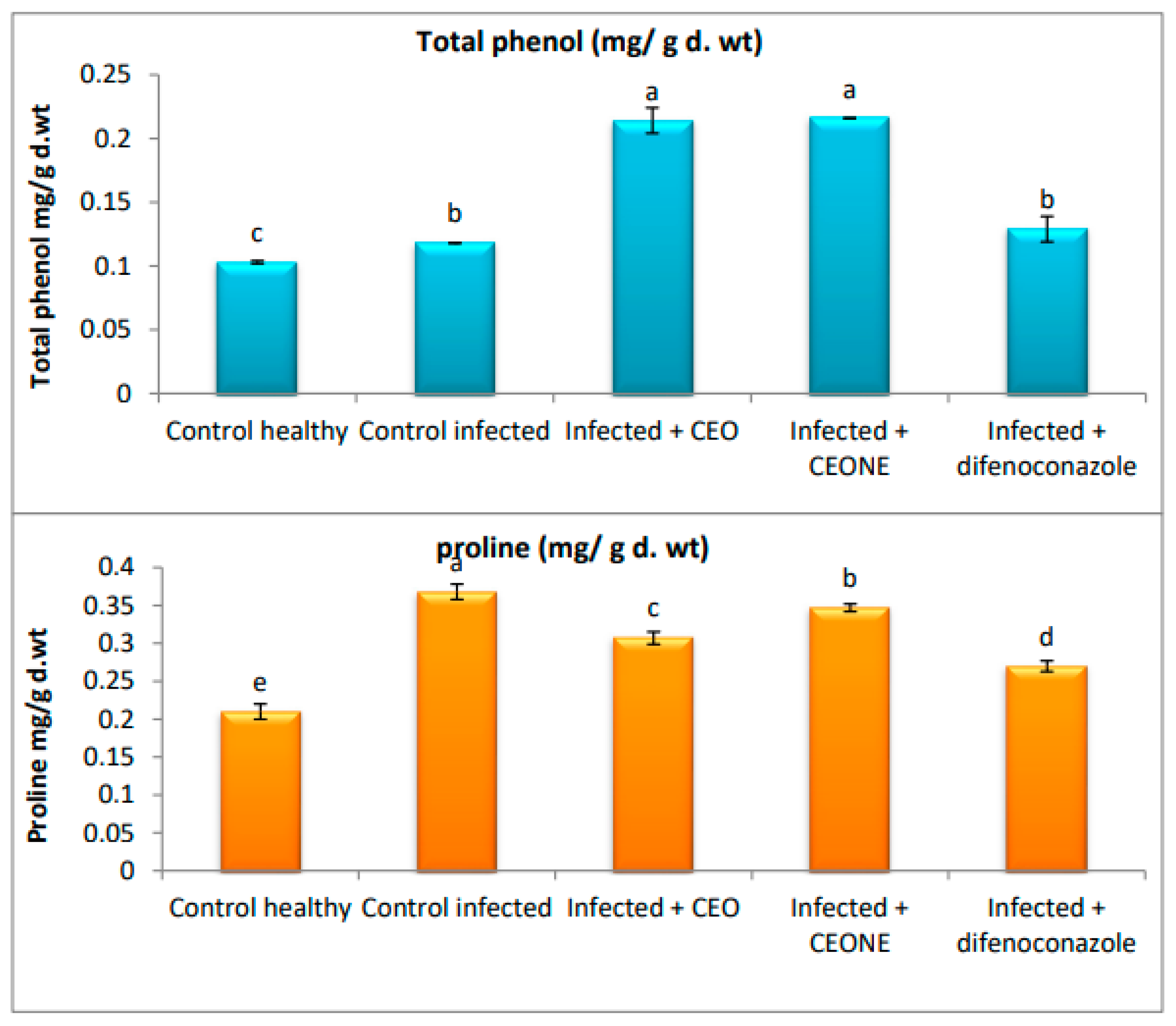
3.6. Proline and Phenol Indicators
3.7. MDA and H2O2
3.8. Antioxidant Enzymes Activity
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gabler, J. Breeding for resistance to biotic and abiotic factors in medicinal and aromatic plants: General situation and current results in annual caraway (Carum carvi L. var. annum). J. Herbs Spices Med. Plants 2002, 9, 1–11. [Google Scholar] [CrossRef]
- Pickersgill, B. Spices. In The Cultural History of Plants; Routledge: Abingdon, UK, 2012; pp. 156–175. [Google Scholar]
- Nabhan, G.P. Cumin, Camels, and Caravans: A Spice Odyssey; University of California Press: Berkeley, CA, USA, 2014; Volume 45. [Google Scholar]
- Mazur, S.; Nawrocki, J. Fungal diseases threat on caraway plantations in the south region of Poland. Acta Fytotech. Zootech. 2004, 7, 201–203. [Google Scholar]
- Khalil, A.M.; Ahmed, A.F.; Mahmoud, E.E.; Abdelaziz, A.M. Influence of organic farming system on microbial biomass and fungal communities of agricultural soil. Afr. J. Mycol. Biotechnol. 2015, 20, 23–40. [Google Scholar]
- Alamri, M.; Alghamdi, H.; Althawadi, S.; Mutabaggani, M.; Dababo, M.A.; Alajlan, F.; Alzayer, M.; Doumith, M.; Alghoribi, M.; Almaghrabi, R.S. Invasive fungal infection of the brain caused by Neoscytalidium dimidiatum in a post-renal transplant patient: A case report. Med. Mycol. Case Rep. 2021, 34, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Elshafie, A.E.; Ba-Omar, T. First report of Albizia lebbeck dieback caused by Scytalidium dimidiatum in Oman. Mycopathologia 2002, 154, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.; Heidarian, A.; Farrokhi-Nejad, R. Citrus branch wilt, decline and death caused by Nattrassia mangiferae and its other hosts in Khuzestan province. Iran. J. Plant Pathol. 2000, 36, Pe77–Pe97. [Google Scholar]
- Al Raish, S.M.; Saeed, E.E.; Sham, A.; Alblooshi, K.; El-Tarabily, K.A.; AbuQamar, S.F. Molecular characterization and disease control of stem canker on royal poinciana (Delonix regia) caused by Neoscytalidium dimidiatum in the United Arab Emirates. Int. J. Mol. Sci. 2020, 21, 1033. [Google Scholar] [CrossRef] [Green Version]
- Abdelaziz, A.M.; Hashem, A.H.; El-Sayyad, G.S.; El-Wakil, D.A.; Selim, S.; Alkhalifah, D.H.M.; Attia, M.S. Biocontrol of soil borne diseases by plant growth promoting rhizobacteria. Trop. Plant Pathol. 2023, 48, 105–127. [Google Scholar] [CrossRef]
- Abdelaziz, A.M.; Kalaba, M.H.; Hashem, A.H.; Sharaf, M.H.; Attia, M.S. Biostimulation of tomato growth and biocontrol of Fusarium wilt disease using certain endophytic fungi. Bot. Stud. 2022, 63, 34. [Google Scholar] [CrossRef]
- Attia, M.S.; Hashem, A.H.; Badawy, A.A.; Abdelaziz, A.M. Biocontrol of early blight disease of eggplant using endophytic Aspergillus terreus: Improving plant immunological, physiological and antifungal activities. Bot. Stud. 2022, 63, 26. [Google Scholar] [CrossRef]
- Abdelaziz, A.M.; Elshaer, M.A.; Abd-Elraheem, M.A.; Ali, O.M.O.M.; Haggag, M.I.; El-Sayyad, G.S.; Attia, M.S. Ziziphus spina-christi extract-stabilized novel silver nanoparticle synthesis for combating Fusarium oxysporum-causing pepper wilt disease: In vitro and in vivo studies. Arch. Microbiol. 2023, 205, 69. [Google Scholar] [CrossRef] [PubMed]
- Farrag, A.; Attia, M.S.; Younis, A.; Abd Elaziz, A. Potential impacts of elicitors to improve tomato plant disease resistance. Al Azhar. Bull. Sci. 2017, 9, 311–321. [Google Scholar]
- Attia, M.S.; El-Wakil, D.A.; Hashem, A.H.; Abdelaziz, A.M. Antagonistic Effect of Plant Growth-Promoting Fungi Against Fusarium Wilt Disease in Tomato: In vitro and In vivo Study. Appl. Biochem. Biotechnol. 2022, 194, 5100–5118. [Google Scholar] [CrossRef] [PubMed]
- Abdelaziz, A.M.; El-Wakil, D.A.; Attia, M.S.; Ali, O.M.; AbdElgawad, H.; Hashem, A.H. Inhibition of Aspergillus flavus growth and aflatoxin production in Zea mays L. using endophytic Aspergillus fumigatus. J. Fungi 2022, 8, 482. [Google Scholar] [CrossRef]
- Attia, M.S.; Younis, A.M.; Ahmed, A.F.; Elaziz, A. Comprehensive management for wilt disease caused by Fusarium oxysporum in tomato plant. Int. J. Innov. Sci. Eng. Technol. 2016, 4, 2348–7968. [Google Scholar]
- Abdelaziz, A.M.; Salem, S.S.; Khalil, A.M.A.; El-Wakil, D.A.; Fouda, H.M.; Hashem, A.H. Potential of biosynthesized zinc oxide nanoparticles to control Fusarium wilt disease in eggplant (Solanum melongena) and promote plant growth. BioMetals 2022, 35, 601–616. [Google Scholar] [CrossRef]
- Hashem, A.H.; Abdelaziz, A.M.; Attia, M.S.; Salem, S.S. Selenium and nano-selenium-mediated biotic stress tolerance in plants. In Selenium and Nano-Selenium in Environmental Stress Management and Crop Quality Improvement; Springer: Berlin/Heidelberg, Germany, 2022; pp. 209–226. [Google Scholar]
- Das, P.; Karankar, V.S. New avenues of controlling microbial infections through anti-microbial and anti-biofilm potentials of green mono-and multi-metallic nanoparticles: A review. J. Microbiol. Methods 2019, 167, 105766. [Google Scholar] [CrossRef] [PubMed]
- Elbasuney, S.; El-Sayyad, G.S.; Attia, M.S.; Abdelaziz, A.M. Ferric oxide colloid: Towards green nano-fertilizer for tomato plant with enhanced vegetative growth and immune response against fusarium wilt disease. J. Inorg. Organomet. Polym. Mater. 2022, 32, 4270–4283. [Google Scholar] [CrossRef]
- Khattab, A.M.; Abo-Taleb, H.A.; Abdelaziz, A.M.; El-Tabakh, M.A.M.; El-Feky, M.M.M.; Abu-Elghait, M. Daphnia magna and Gammarus pulex, novel promising agents for biomedical and agricultural applications. Sci. Rep. 2022, 12, 13690. [Google Scholar] [CrossRef] [PubMed]
- Raveau, R.; Fontaine, J.; Sahraoui, A.L.-H. Essential oils as potential alternative biocontrol products against plant pathogens and weeds: A review. Foods 2020, 9, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar-Veloz, L.M.; Calderón-Santoyo, M.; González, Y.V.; Ragazzo-Sánchez, J.A. Application of essential oils and polyphenols as natural antimicrobial agents in postharvest treatments: Advances and challenges. Food Sci. Nutr. 2020, 8, 2555–2568. [Google Scholar] [CrossRef]
- Veloz-García, R.; Marín-Martínez, R.; Veloz-Rodríguez, R.; Rodríguez-Guerra, R.; Torres-Pacheco, I.; González-Chavira, M.M.; Anaya-López, J.L.; Guevara-Olvera, L.; Feregrino-Pérez, A.A.; Loarca-Piña, G.; et al. Antimicrobial activities of cascalote (Caesalpinia cacalaco) phenolics-containing extract against fungus Colletotrichum lindemuthianum. Ind. Crops Prod. 2010, 31, 134–138. [Google Scholar] [CrossRef]
- Yang, R.; Miao, J.; Chen, X.; Chen, C.; Simal-Gandara, J.; Chen, J.; Wan, C. Essential oils nano-emulsion confers resistance against Penicillium digitatum in ‘Newhall’ navel orange by promoting phenylpropanoid metabolism. Ind. Crops Prod. 2022, 187, 115297. [Google Scholar] [CrossRef]
- Giunti, G.; Palermo, D.; Laudani, F.; Algeri, G.M.; Campolo, O.; Palmeri, V. Repellence and acute toxicity of a nano-emulsion of sweet orange essential oil toward two major stored grain insect pests. Ind. Crops Prod. 2019, 142, 111869. [Google Scholar] [CrossRef]
- Purdy, L.H. Sclerotinia sclerotiorum: History, diseases and symptomatology, host range, geographic distribution, and impact. Phytopathology 1979, 69, 875–880. [Google Scholar] [CrossRef]
- Crous, P.W.; Slippers, B.; Wingfield, M.J.; Rheeder, J.; Marasas, W.F.O.; Philips, A.J.L.; Alves, A.; Burgess, T.; Barber, P.; Groenewald, J.Z. Phylogenetic lineages in the Botryosphaeriaceae. Stud. Mycol. 2006, 55, 235–253. [Google Scholar] [CrossRef] [Green Version]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.K.; Tangthirasunun, N.; Phillips, A.J.; Dai, D.Q.; Wanasinghe, D.N.; Wen, T.C.; Bahkali, A.H.; Hyde, K.D.; Kang, J.C. Morphology and Phylogeny of Neoscytalidium orchidacearum sp. nov. (Botryosphaeriaceae). Mycobiology 2016, 44, 79–84. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Pearce, L.; Lilley, G.; Coloe, S.; Baird, R.; Pedersen, J. PCR identification of dermatophyte fungi Trichophyton rubrum, T. soudanense and T. gourvilii. J. Med. Microbiol. 2002, 51, 117–122. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Al-Shahrani, M.H.; Mahfoud, M.; Anvarbatcha, R.; Athar, T.; Al Asmari, A. Evaluation of antifungal activity and cytotoxicity of Thymus vulgaris essential oil. Pharmacogn. Commun. 2017, 7, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Ghotbi, R.S.; Khatibzadeh, M.; Kordbacheh, S. Preparation of neem seed oil nanoemulsion. In Proceedings of the 5th International Conference on Nanotechnology: Fundamentals and Applications, Prague, Czech Republic, 11–13 August 2014. [Google Scholar]
- Saloko, S.; Darmadji, P.; Setiaji, B.; Pranoto, Y.; Anal, A.K. Encapsulation of coconut shell liquid smoke in chitosan-maltodextrin based nanoparticles. Int. Food Res. J. 2013, 20, 1269. [Google Scholar]
- Al-Bedak, O.A.; Mohamed, R.A.; Seddek, N.H. First detection of Neoscytalidium dimidiatum associated with canker disease in Egyptian Ficus trees. For. Pathol. 2018, 48, e12411. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Dai, G.H.; Andary, C.; Cosson-Mondolot, L.; Boubals, D. Polyphenols and resistance of grapevines to downy mildew. In International Symposium on Natural Phenols in Plant Resistance 381; ISHS: Coimbatore, India, 1993. [Google Scholar]
- Hu, Z.; Richter, H.; Sparovek, G.; Schnug, E. Physiological and biochemical effects of rare earth elements on plants and their agricultural significance: A review. J. Plant Nutr. 2004, 27, 183–220. [Google Scholar] [CrossRef]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Srivastava, S. Peroxidase and poly-phenol oxidase in Brassica juncea plants infected with Macrophomina phaseolina (Tassai) Goid and their implication in disease resistance. J. Phytopathol. 1987, 120, 249–254. [Google Scholar] [CrossRef]
- Matta, A. Accumulation of phenols in tomato plants infected by different forms of Fusarium oxysporum. Phytopathology 1969, 59, 512–513. [Google Scholar]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 2nd ed.; Iowa State University Press: Ames, IA, USA, 1982; Volume 507. [Google Scholar]
- Mohd, M.H.; Salleh, B.; Zakaria, L. Identification and molecular characterizations of Neoscytalidium dimidiatum causing stem canker of red-fleshed dragon fruit (Hylocereus polyrhizus) in Malaysia. J. Phytopathol. 2013, 161, 841–849. [Google Scholar] [CrossRef]
- Derviş, S.; Özer, G.; Türkölmez, Ş. First report of Neoscytalidium dimidiatum causing tuber rot of potato in Turkey. J. Plant Pathol. 2020, 102, 1295–1296. [Google Scholar] [CrossRef]
- Ghosh, V.; Mukherjee, A.; Chandrasekaran, N. Eugenol-loaded antimicrobial nanoemulsion preserves fruit juice against, microbial spoilage. Colloids Surf. B Biointerfaces 2014, 114, 392–397. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, A.M.; Jäger, E.; Jäger, A.; Stepánek, P.; Giacomelli, F.C. Physicochemical aspects behind the size of biodegradable polymeric nanoparticles: A step forward. Colloids Surf. A Physicochem. Eng. Asp. 2013, 436, 1092–1102. [Google Scholar] [CrossRef]
- Sajjadi, S.; Zerfa, M.; Brooks, B.W. Dynamic behaviour of drops in oil/water/oil dispersions. Chem. Eng. Sci. 2002, 57, 663–675. [Google Scholar] [CrossRef]
- Sampathi, S.; Mankala, S.K.; Wankar, J.; Dodoala, S. Nanoemulsion Based Hydrogels of Itraconazole for Transdermal Drug Delivery; NISCAIR-CSIR: New Delhi, India, 2015. [Google Scholar]
- Hammad, E.A.; Hasanin, M.M.H. Antagonistic Effect of Nanoemulsions of Some Essential Oils against Fusarium oxysporum and Root-Knot Nematode Meloidogyne javanica on Coleus Plants. Pak. J. Nematol. 2022, 40, 35–48. [Google Scholar] [CrossRef]
- Morcia, C.; Malnati, M.; Terzi, V. In vitro antifungal activity of terpinen-4-ol, eugenol, carvone, 1, 8-cineole (eucalyptol) and thymol against mycotoxigenic plant pathogens. Food Addit. Contam. Part A 2012, 29, 415–422. [Google Scholar]
- Matan, N.; Nisoa, M.; Matan, N.; Aewsiri, T. Effect of cold atmospheric plasma on antifungal activities of clove oil and eugenol against molds on areca palm (Areca catechu) leaf sheath. Int. Biodeterior. Biodegrad. 2014, 86, 196–201. [Google Scholar] [CrossRef]
- e Oliveira Pereira, F.; Mendes, J.M.; de Oliveira Lima, E. Investigation on mechanism of antifungal activity of eugenol against Trichophyton rubrum. Med. Mycol. 2013, 51, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Gill, A.; Holley, R. Inhibition of membrane bound ATPases of Escherichia coli and Listeria monocytogenes by plant oil aromatics. Int. J. Food Microbiol. 2006, 111, 170–174. [Google Scholar] [CrossRef]
- Braga, P.; Sasso, M.D.; Culici, M.; Alfieri, M. Eugenol and thymol, alone or in combination, induce morphological alterations in the envelope of Candida albicans. Fitoterapia 2007, 78, 396–400. [Google Scholar] [CrossRef]
- Amiri, A.; Dugas, R.; Pichot, A.L.; Bompeix, G. In vitro and in vitro activity of eugenol oil (Eugenia caryophylata) against four important postharvest apple pathogens. Int. J. Food Microbiol. 2008, 126, 13–19. [Google Scholar] [CrossRef]
- Hassanin, M. Pathological Studies on Root Rot and Wilt of Black Cumin (Nigella sativa) and Their Management in Egypt. Ph.D. Thesis, Faculty of Agriculture, Al-Azhar University, Cairo, Egypt, 2013; 137p. [Google Scholar]
- Shahbazi, Y. Antioxidant, antibacterial, and antifungal properties of nanoemulsion of clove essential oil. Nanomed. Res. J. 2019, 4, 204–208. [Google Scholar]
- Šernaitė, L.; Rasiukevičiūtė, N.; Valiuškaitė, A. The Extracts of cinnamon and clove as potential biofungicides against strawberry grey mould. Plants 2020, 9, 613. [Google Scholar] [CrossRef] [PubMed]
- Mahdi, A.A.; Al-Maqtari, Q.A.; Mohammed, J.K.; Al-Ansi, W.; Cui, H.; Lin, L. Enhancement of antioxidant activity, antifungal activity, and oxidation stability of Citrus reticulata essential oil nanocapsules by clove and cinnamon essential oils. Food Biosci. 2021, 43, 101226. [Google Scholar] [CrossRef]
- Bagherabadi, S.; Zafari, D.; Maharachchikumbura, S.S. Neoscytalidium dimidiatum as One of the Fungal Agents Associated with Walnut Decline in Iran. J. Nuts 2022, 13, 31–39. [Google Scholar]
- Güney, I.G.; Özer, G.; Türkölmez, Ş.; Derviş, S. Canker and leaf scorch on olive (Olea europaea L.) caused by Neoscytalidium dimidiatum in Turkey. Crop Prot. 2022, 157, 105985. [Google Scholar] [CrossRef]
- Tuan, L.N.A.; Du, B.D.; Ha, L.D.T.; Dzung, L.T.K.; Van Phu, D.; Hien, N.Q. Induction of chitinase and brown spot disease resistance by oligochitosan and nanosilica–oligochitosan in dragon fruit plants. Agric. Res. 2019, 8, 184–190. [Google Scholar] [CrossRef]
- Hou, T.; Sana, S.S.; Li, H.; Xing, Y.; Nanda, A.; Netala, V.R.; Zhang, Z. Essential oils and its antibacterial, antifungal and anti-oxidant activity applications: A review. Food Biosci. 2022, 47, 101716. [Google Scholar] [CrossRef]
- Milind, P.; Deepa, K. Clove: A champion spice. Int. J. Res. Ayurveda Pharm. 2011, 2, 47–54. [Google Scholar]
- Ertas, O.N.; Guler, T.; Çiftçi, M.; DalkIlIç, B.; Simsek, U.G. The effect of an essential oil mix derived from oregano, clove and anise on broiler performance. Int. J. Poult. Sci. 2005, 4, 879–884. [Google Scholar]
- Sattary, M.; Amini, J.; Hallaj, R. Antifungal activity of the lemongrass and clove oil encapsulated in mesoporous silica nanoparticles against wheat’s take-all disease. Pestic. Biochem. Physiol. 2020, 170, 104696. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Kumar, V.; Kohli, S.K.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Bhardwaj, R.; Zheng, B. Phytohormones regulate accumulation of osmolytes under abiotic stress. Biomolecules 2019, 9, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rejeb, K.B.; Abdelly, C.; Savouré, A. How reactive oxygen species and proline face stress together. Plant Physiol. Biochem. 2014, 80, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Lucas, G.C.; Alves, E.; Pereira, R.B.; Zacaroni, A.; Perina, F.; de Souza, R.M. Indian clove essential oil in the control of tomato bacterial spot. J. Plant Pathol. 2012, 94, 45–51. [Google Scholar]
- Kumar, P.; Lokesh, V.; Doddaraju, P.; Kumari, A.; Singh, P.; Meti, B.S.; Sharma, J.; Gupta, K.J.; Manjunatha, G. Greenhouse and field experiments revealed that clove oil can effectively reduce bacterial blight and increase yield in pomegranate. Food Energy Secur. 2021, 10, e305. [Google Scholar] [CrossRef]
- Shafie, R. Antiviral Activity of Clove Oil Nanoemulsion against Potato virus-Y (PVY). Egypt. J. Phytopathol. 2017, 45, 15–31. [Google Scholar] [CrossRef]
- Fayez, K.A.; Al-Sodany, Y.M.; Abouzaid, A. Impact of greenhouse environmental factors and fungicide Trinol (triadimend) treatment on berry leaves infected with powdery mildew (Uncinula necator (Schwein.) Burrill: Role of host antioxidant systems against pathogen infection. Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 3058–3072. [Google Scholar]
- Attia, M.S.; El-Naggar, H.A.; Abdel-Daim, M.M.; El-Sayyad, G.S. The potential impact of Octopus cyanea extracts to improve eggplant resistance against Fusarium-wilt disease: In vivo and in vitro studies. Environ. Sci. Pollut. Res. 2021, 28, 35854–35869. [Google Scholar] [CrossRef]
- Albalawi, M.A.; Abdelaziz, A.M.; Attia, M.S.; Saied, E.; Elganzory, H.H.; Hashem, A.H. Mycosynthesis of Silica Nanoparticles Using Aspergillus niger: Control of Alternaria solani Causing Early Blight Disease, Induction of Innate Immunity and Reducing of Oxidative Stress in Eggplant. Antioxidants 2022, 11, 2323. [Google Scholar] [CrossRef]
- Lattanzio, V.; Lattanzio, V.M.; Cardinali, A. Role of phenolics in the resistance mechanisms of plants against fungal pathogens and insects. Phytochem. Adv. Res. 2006, 661, 23–67. [Google Scholar]
- Aldinary, A.M.; Abdelaziz, A.M.; Farrag, A.A.; Attia, M.S. Biocontrol of tomato Fusarium wilt disease by a new Moringa endophytic Aspergillus isolates. Mater. Today Proc. 2021, in press. [Google Scholar] [CrossRef]
- Brewer, M. Natural antioxidants: Sources, compounds, mechanisms of action, and potential applications. Compr. Rev. Food Sci. Food Saf. 2011, 10, 221–247. [Google Scholar] [CrossRef]
- Moghaddam, M.R.B.; Le Roy, K.; Xiang, L.; Rolland, F.; Ende, W.V.D. Sugar signalling and antioxidant network connections in plant cells. FEBS J. 2010, 277, 2022–2037. [Google Scholar] [CrossRef]
- Sharma, A.; Sharma, N.K.; Srivastava, A.; Kataria, A.; Dubey, S.; Sharma, S.; Kundu, B. Clove and lemongrass oil based non-ionic nanoemulsion for suppressing the growth of plant pathogenic Fusarium oxysporum f. sp. lycopersici. Ind. Crops Prod. 2018, 123, 353–362. [Google Scholar] [CrossRef]
- Jain, A.; Singh, A.; Singh, S.; Singh, H.B. Microbial consortium-induced changes in oxidative stress markers in pea plants challenged with Sclerotinia sclerotiorum. J. Plant Growth Regul. 2013, 32, 388–398. [Google Scholar] [CrossRef]
- Hosseinifard, M.; Stefaniak, S.; Javid, M.G.; Soltani, E.; Wojtyla, Ł.; Garnczarska, M. Contribution of exogenous proline to abiotic stresses tolerance in plants: A review. Int. J. Mol. Sci. 2022, 23, 5186. [Google Scholar] [CrossRef]
- Dallagnol, L.J.; Rodrigues, F.A.; Martins, S.C.V.; Cavatte, P.C.; DaMatta, F.M. Alterations on rice leaf physiology during infection by Bipolaris oryzae. Australas. Plant Pathol. 2011, 40, 360–365. [Google Scholar] [CrossRef]
- Badawy, A.; Alotaibi, M.; Abdelaziz, A.; Osman, M.; Khalil, A.; Saleh, A.; Mohammed, A.; Hashem, A. Enhancement of Seawater Stress Tolerance in Barley by the Endophytic Fungus Aspergillus ochraceus. Metabolites 2021, 11, 428. [Google Scholar] [CrossRef]
- Dwivedy, A.K.; Kumar, M.; Upadhyay, N.; Prakash, B.; Dubey, N. Plant essential oils against food borne fungi and mycotoxins. Curr. Opin. Food Sci. 2016, 11, 16–21. [Google Scholar] [CrossRef]
- Basavegowda, N.; Baek, K.-H. Synergistic antioxidant and antibacterial advantages of essential oils for food packaging applications. Biomolecules 2021, 11, 1267. [Google Scholar] [CrossRef]
- Jiang, S.; Han, S.; He, D.; Cao, G.; Fang, K.; Xiao, X.; Yi, J.; Wan, X. The accumulation of phenolic compounds and increased activities of related enzymes contribute to early defense against walnut blight. Physiol. Mol. Plant Pathol. 2019, 108, 101433. [Google Scholar] [CrossRef]
- Alici, E.H.; Arabaci, G. Determination of SOD, POD, PPO and cat enzyme activities in Rumex obtusifolius L. Annu. Res. Rev. Biol. 2016, 11, 1–7. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Disease Indicators Levels | DI (Disease Index) (%) | Protection (%) | ||||
|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | |||
| Control infected. | 0 | 0 | 0 | 5 | 5 | 87.5 | 0 |
| Infected plants treated with CEO | 3 | 3 | 2 | 0 | 2 | 37.50 | 57.14 |
| Infected plants treated with CEONE | 4 | 2 | 4 | 0 | 0 | 25 | 71.42 |
| Infected plants treated with difenoconazole fungicide. | 5 | 4 | 0 | 0 | 1 | 20 | 77.14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hashem, A.H.; Abdelaziz, A.M.; Hassanin, M.M.H.; Al-Askar, A.A.; AbdElgawad, H.; Attia, M.S. Potential Impacts of Clove Essential Oil Nanoemulsion as Bio Fungicides against Neoscytalidium Blight Disease of Carum carvi L. Agronomy 2023, 13, 1114. https://doi.org/10.3390/agronomy13041114
Hashem AH, Abdelaziz AM, Hassanin MMH, Al-Askar AA, AbdElgawad H, Attia MS. Potential Impacts of Clove Essential Oil Nanoemulsion as Bio Fungicides against Neoscytalidium Blight Disease of Carum carvi L. Agronomy. 2023; 13(4):1114. https://doi.org/10.3390/agronomy13041114
Chicago/Turabian StyleHashem, Amr H., Amer M. Abdelaziz, Mahmoud M. H. Hassanin, Abdulaziz A. Al-Askar, Hamada AbdElgawad, and Mohamed S. Attia. 2023. "Potential Impacts of Clove Essential Oil Nanoemulsion as Bio Fungicides against Neoscytalidium Blight Disease of Carum carvi L." Agronomy 13, no. 4: 1114. https://doi.org/10.3390/agronomy13041114