
Water Relation, Gas Exchange Characteristics and Yield Performance of Selected Mungbean Genotypes under Low Soil Moisture Condition

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
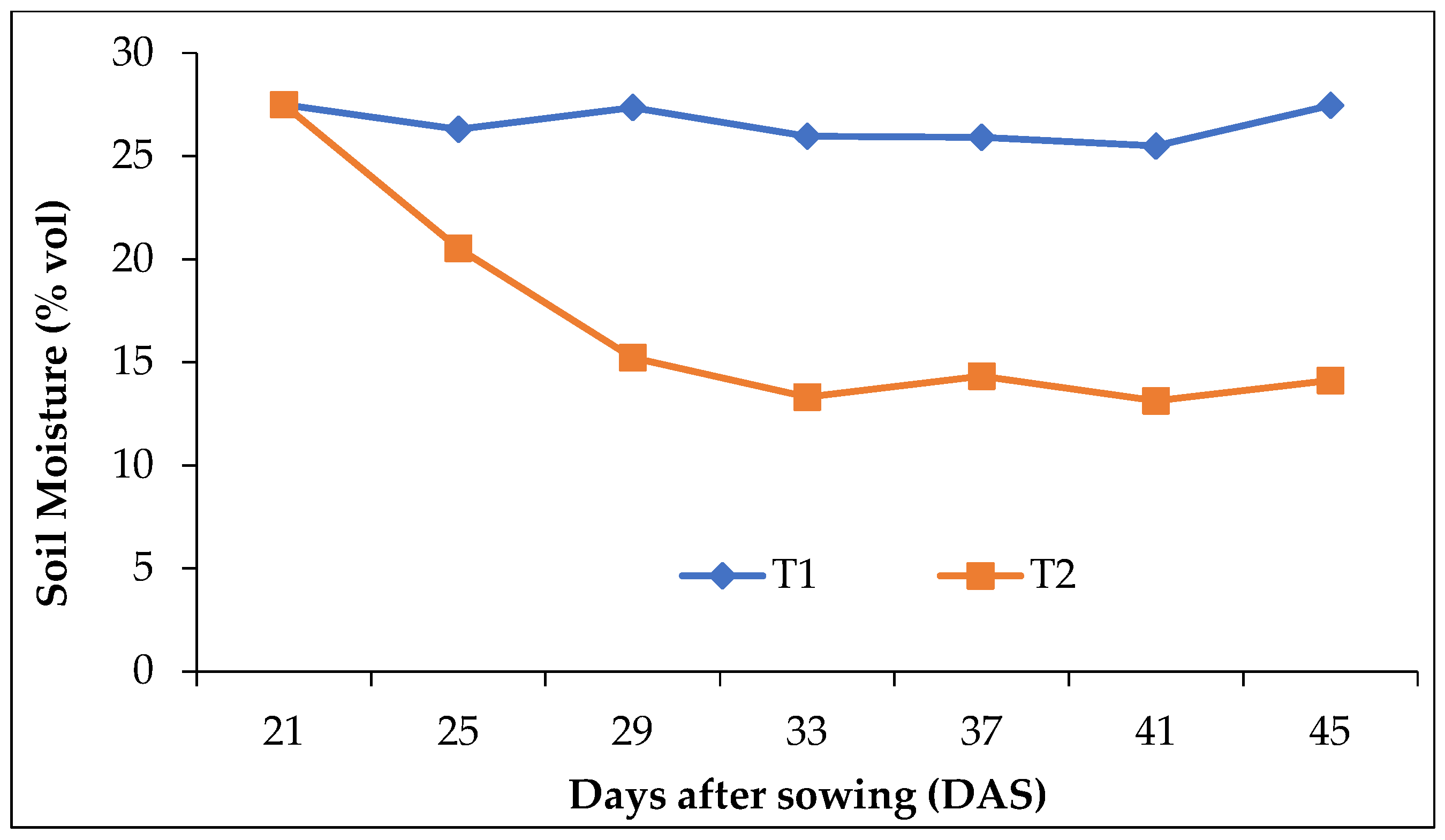
2.1. Experimental Variables and Treatments
2.2. Preparation of Pot and Fertilizer Application
2.3. Seed Sowing and Treatment Imposition
2.4. Collection of Data
2.5. Estimation of Water Relation Parameters
2.6. Gas Exchange Characteristics
2.7. Estimation of Total Chlorophyll Content
2.8. Proline Determination
2.9. Yield Attributes and Yield
2.10. Analysis of the Data
3. Results
3.1. Water Status of Mungbean
3.1.1. Relative Water Content
3.1.2. Water Saturation Deficit
3.1.3. Water Retention Capacity
3.1.4. Water Uptake Capacity
3.2. Gas Exchange Characteristics
3.2.1. Photosynthetic Rate
3.2.2. Stomatal Conductance
3.2.3. Transpiration Rate
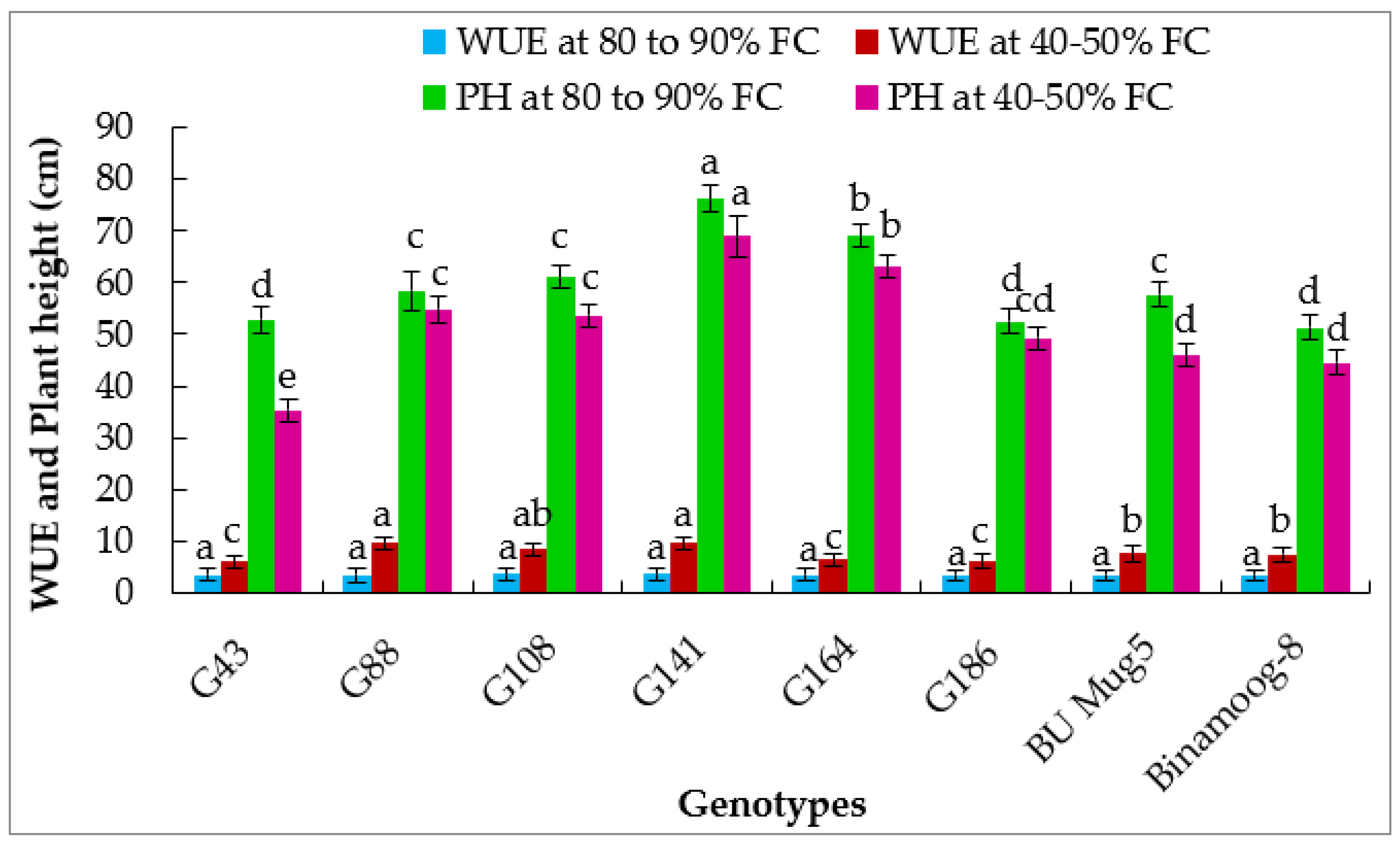
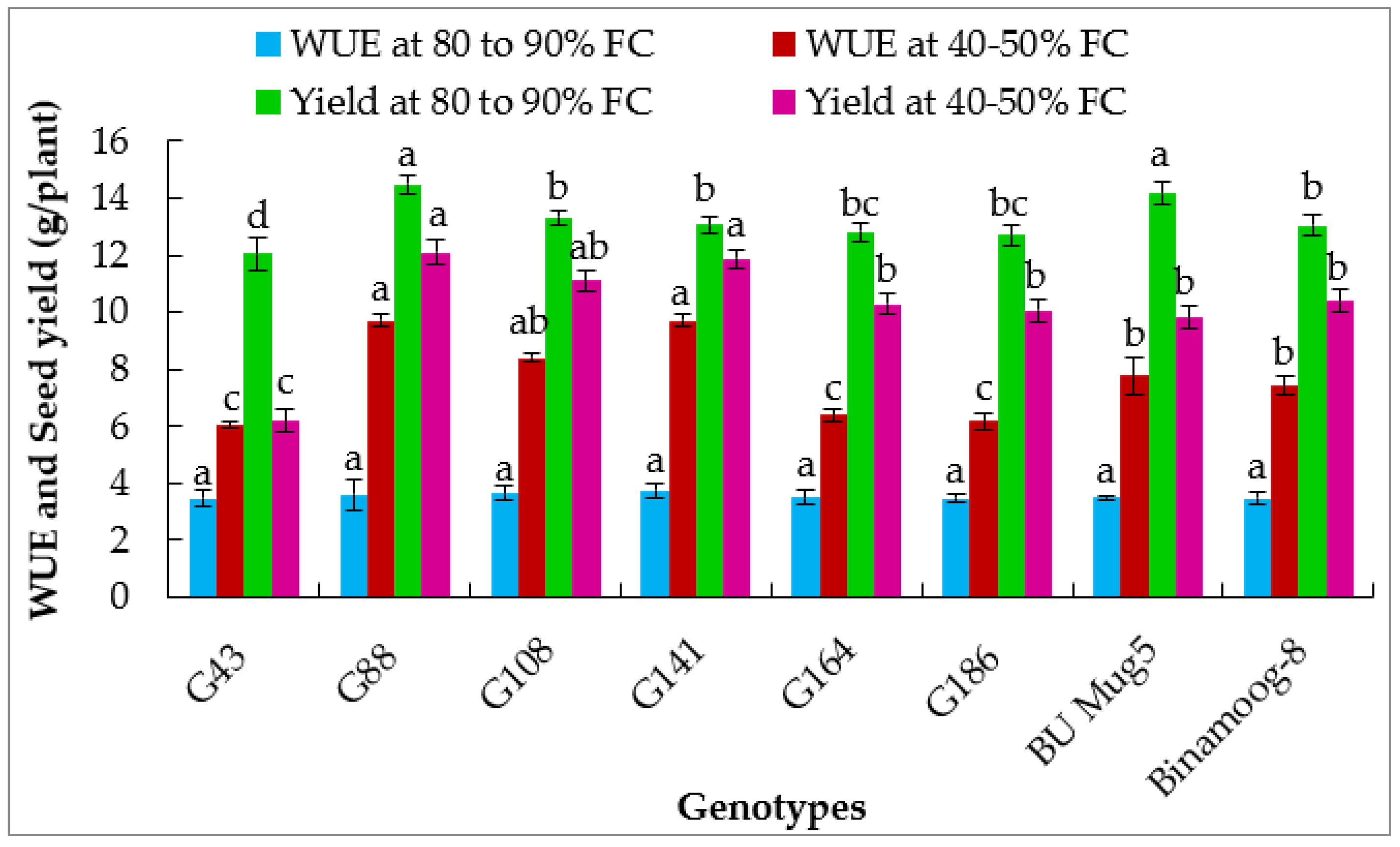
3.2.4. Photosynthetic Water Use Efficiency
3.3. Chlorophyll Contents
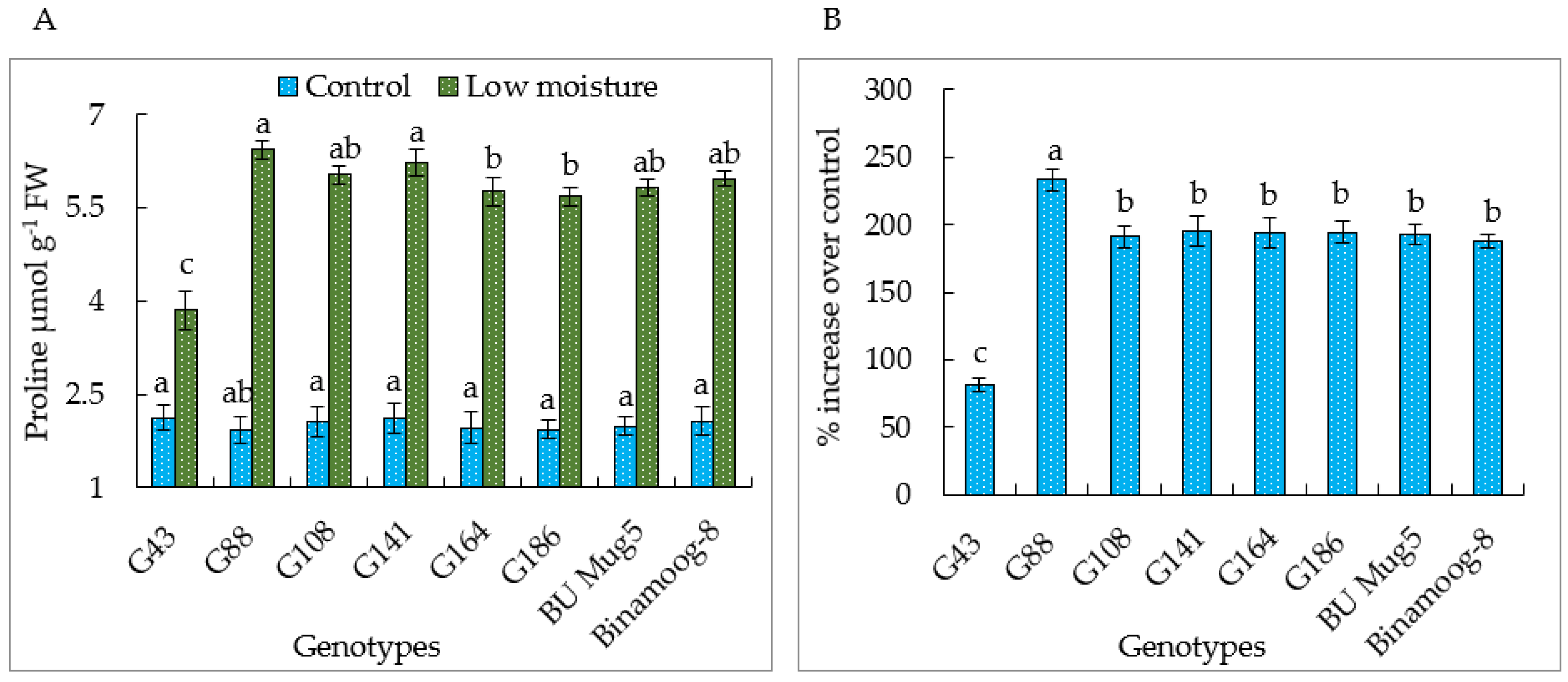
3.4. Proline Content
3.5. Yield Attributes and Yield
3.5.1. Plant Height
3.5.2. Number of Branches per Plant
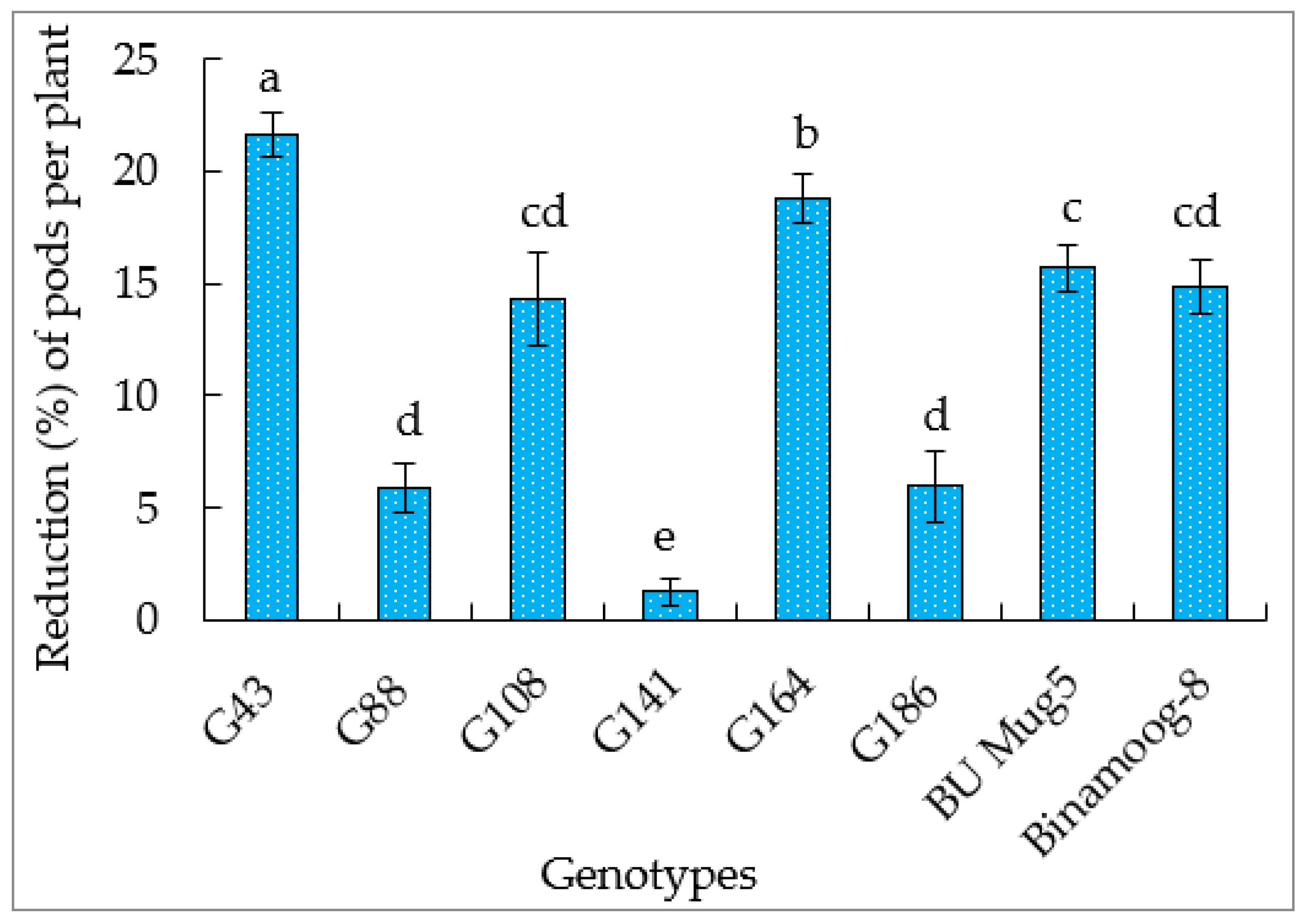
3.5.3. Number of Pods per Plant
3.5.4. Pod Length
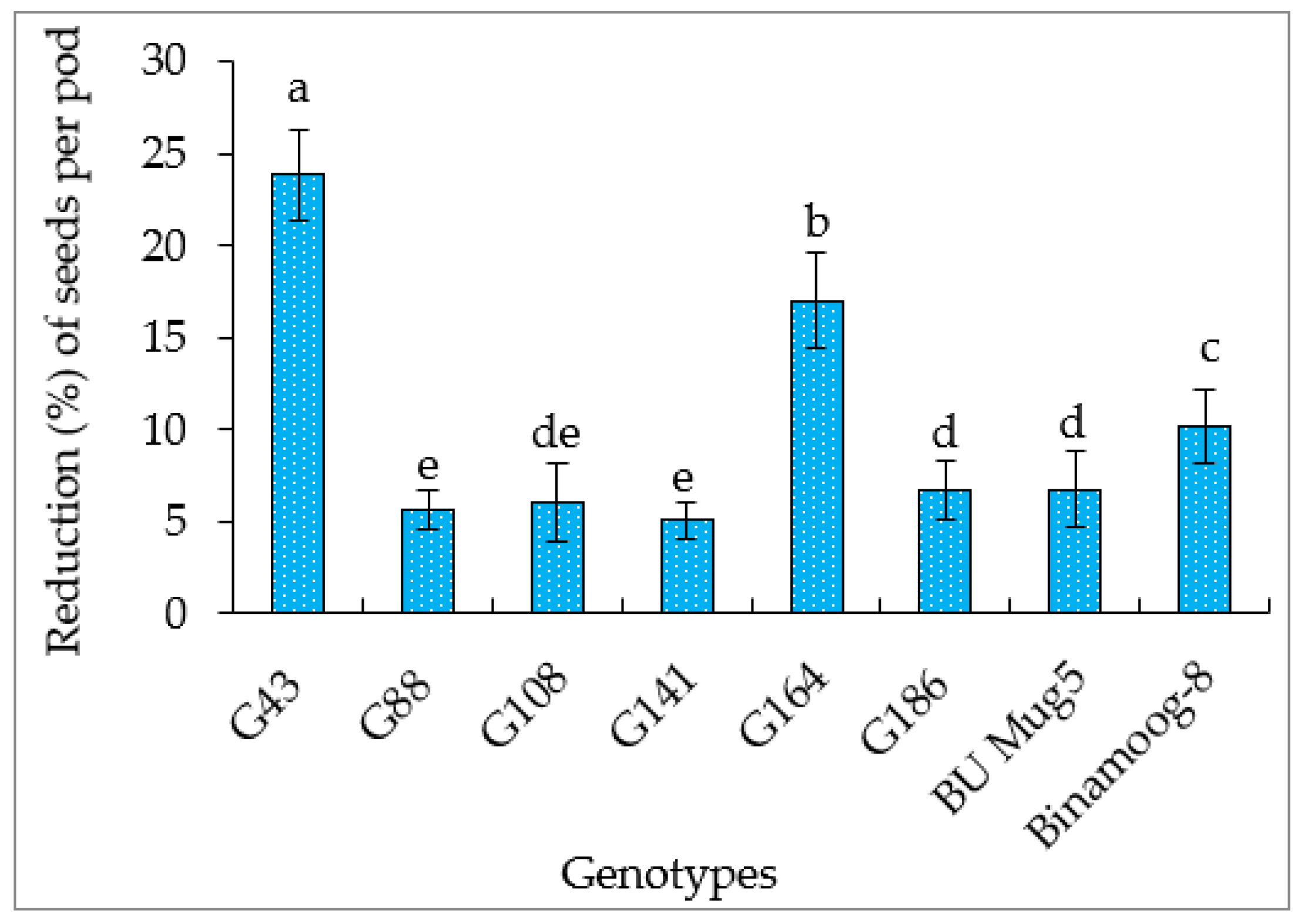
3.5.5. Number of Seeds per Pod
3.5.6. Thousand Seed Weight
3.5.7. Seed Yield
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, C.M.; Singh, P.; Tiwari, C.; Purwar, S.; Kumar, M.; Pratap, A.; Singh, S.; Chugh, V.; Mishra, A.K. Improving Drought Tolerance in Mungbean (Vigna radiate L.Wilczek): Morpho-Physiological, Biochemical and Molecular Perspectives. Agronomy 2021, 11, 1534. [Google Scholar] [CrossRef]
- Hozayn, M.; El-Monem, A.A.A.; El-Hamid, E.M.E.M.A.; Qados, A.M.S.A. Amelioration of salinity stress in mungbean (Vigna radiata L.). plant by soaking in arginine. Res. J. Appl. Sci. 2013, 9, 393–401. [Google Scholar]
- Guo, X.; Li, T.; Tang, K.; Liu, R.H. Effect of germination on phytochemical profiles and antioxidant activity of mung bean sprouts (Vigna radiata). J. Agric. Food Chem. 2012, 60, 11050–11055. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.P.; Singh, B.B.; Pratap, A. Genetic improvement of mungbean and urdbean and their role in enhancing pulse production in India. Indian J. Genet. Plant Breed. 2016, 76, 550–567. [Google Scholar] [CrossRef] [Green Version]
- Lindström, K.; Mousavi, S.A. Effectiveness of nitrogen fixation in rhizobia. Microb. Biotechnol. 2020, 13, 1314–1335. [Google Scholar] [CrossRef] [Green Version]
- Allito, B.B.; Nana, E.M.; Alemneh, A.A. Rhizobia strain and legume genome interaction effects on nitrogen fixation and yield of grain legume: A review. Mol. Soil Biol. 2015, 6, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Basu, S.; Ramegowda, V.; Kumar, A.; Pereira, A. Plant adaptation to drought stress. F1000Research 2016, 5, 1–10. [Google Scholar] [CrossRef]
- Bangar, P.; Chaudhury, A.; Tiwari, B.; Kumur, S.; Kumari, R.; Bhat, K.V. Morphophysiological and biochemical response of mungbean [Vigna radiata (L.) Wilczek] varieties at different developmental stages under drought stress. Turk. J. Biol. 2019, 43, 58–69. [Google Scholar] [CrossRef]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.N.; Wang, L.C. Chilling and drought stresses in crop plants: Implications, cross talk, and potential management opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef]
- Hayatu, M.; Muhammad, S.Y.; Abdu, H.U. Effect of water stress on the leaf relative water content and yield of some cowpea (Vigna unguiculata (L) genotype. Int. J. Sci. Technol. Res. 2014, 3, 148–152. [Google Scholar]
- Baroowa, B.; Gogoi, N.; Farooq, M. Changes in physiological, biochemical and antioxidant enzyme activities of green gram (Vigna radiata L.) genotypes under drought. Acta Physiol. Plant. 2016, 38, 219. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Sankar, B.; Murali, P.V.; Gomathinayagam, M.; Lakshmanan, G.M.A.; Panneerselvam, R. Water deficit stress effects on reactive oxygen metabolism in Catharanthus roseus; impacts on ajmalicine accumulation. Colloids Surf. B Biointerfaces 2008, 62, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. In Sustainable Agriculture; Springer: Dordrecht, The Netherlands, 2009; pp. 153–188. [Google Scholar]
- Siddiqui, M.H.; Khan, M.N.; Singh, V.P. Hot and dry: How plants can thrive in future climates. Plant Cell Rep. 2022, 41, 497–499. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, M.; Li, J.; Yahya, M.; Wang, M.; Ali, A.; Cheng, A.; Ma, C. Grain legumes and fear of salt stress: Focus on mechanisms and management strategies. Int. J. Mol. Sci. 2019, 20, 799. [Google Scholar] [CrossRef] [Green Version]
- Rahdari, P.; Hosseini, S.M.; Tavakoli, S. The studying effect of drought stress on germination, proline, sugar, lipid, protein and chlorophyll content in purslane (Portulaca oleracea L.) leaves. J. Med. Plant Res. 2012, 6, 1539–1547. [Google Scholar]
- Anjum, S.A.; Xie, X.Y.; Wang, L.C.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Zare, M.; Dehghani, B.; Alizadeh, O.; Azarpanah, A. The evaluation of various agronomic traits of mungbean (Vigna radiata L.) genotypes under drought stress and non-stress conditions. Int. J. Farming Allied Sci. 2013, 2, 764–770. [Google Scholar]
- BARC. Bangladesh Agricultural Research Council. Fertilizer Recommendation Guide; BARC: Farmgate, Dhaka, Bangladesh, 2018. [Google Scholar]
- Barrs, H.; Weatherley, P.A. Re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Sangakkara, U.R.; Hartwig, U.A.; Nösberger, J. Response of root branching and shoot water potentials of french beans (Phaseolus vulgaris L.) to soil moisture and fertilizer potassium. J. Agron. Crop Sci. 1996, 177, 165–173. [Google Scholar] [CrossRef]
- Fadeels, A.A. Location and properties of chloroplasts and pigment determination in roots. Physiol. Plant. 1962, 15, 130–147. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Imtiaz, A.A.; Shahriar, S.A.; Baque, M.A.; Eaty, M.N.K.; Falguni, M.R. Screening of Mungbean Genotypes under Polyethylene Glycol (PEG) Induced Drought Stress Condition. Annu. Res. Rev. Biol. 2020, 35, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.R.; Kamal, M.M.; Alam, M.A.; Hossain, J.; Soufan, W.; Skalicky, M.; Brestic, M.; Habib-ur-Rahman, M.; EL Sabagh, A.; Islam, M.S. Physiochemical Changes of Mung Bean [Vigna radiata (L.) R. Wilczek] in Responses to Varying Irrigation Regimes. Horticulturae 2021, 7, 565. [Google Scholar] [CrossRef]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Rao, D.E.; Chaitanya, K.V. Photosynthesis and antioxidative defense mechanisms in deciphering drought stress tolerance of crop plants. Biol. Plant. 2016, 60, 201–218. [Google Scholar] [CrossRef]
- Yang, F.; Miao, L.F. Adaptive responses to progressive drought stress in two poplar species originating from different altitudes. Silva Fenn. 2010, 44, 23–37. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Jin, Z.Q.; Zhao, J.; Zhang, G.; Wu, F. Physiological and biochemical responses to drought stress in cultivated and Tibetan wild barley. Plant Growth Regul. 2015, 75, 567–574. [Google Scholar] [CrossRef]
- Chowdhury, J.A.; Karim, M.A.; Khaliq, Q.A.; Ahmed, A.U.; Mondol, A.M. Effect of drought stress on water relation traits of four soybean genotypes. SAARC J. Agric. 2017, 15, 163–175. [Google Scholar] [CrossRef] [Green Version]
- Zhu, R.; Wu, F.Y.; Zhou, S.; Hu, T.; Huang, J.; Gao, Y. Cumulative effects of drought-flood abrupt alternation on the photosynthetic characteristics of rice. Environ. Exp. Bot. 2020, 169, 103901. [Google Scholar] [CrossRef]
- Alderfasi, A.A.; Alzarqaa, A.A.; AL-Yahya, F.A.; Roushdy, S.S.; Dawabah, A.A.; Alhammad, B.A. Effect of combined biotic and abiotic stress on some physiological aspects and antioxidant enzymatic activity in mungbean (Vigna radiata L.). Afr. J. Agric. Res. 2017, 12, 700–705. [Google Scholar]
- Farooq, M.; Kobayashi, N.; Wahid, A.; Ito, O.; Basra, S.M.A. Strategies for producing more rice with less water. Adv. Agron. 2009, 101, 351–388. [Google Scholar]
- Mishra, S.S.; Behera, P.K.; Kumar, V.; Lenka, S.K.; Panda, D. Physiological characterization and allelic diversity of selected drought tolerant traditional rice (Oryza sativa L.) landraces of Koraput, India. Physiol. Mol. Biol. Plants. 2018, 24, 1035–1046. [Google Scholar] [CrossRef]
- Alsamadany, H. Physiological, biochemical and molecular evaluation of mungbean genotypes for agronomical yield under drought and salinity stresses in the presence of humic acid. Saudi J. Biol. Sci. 2022, 29, 103385. [Google Scholar] [CrossRef]
- Meena, S.K.; Pandey, R.; Sharma, S.; Kumar, T.; Singh, M.P.; Dikshit, H.K. Physiological basis of combined stress tolerance to low phosphorus and drought in a diverse set of mungbean germplasm. Agronomy 2021, 11, 99. [Google Scholar] [CrossRef]
- Panda, D.; Mishra, S.S.; Behera, P.K. Drought tolerance in rice: Focus on recent mechanisms and approaches. Rice Sci. 2021, 28, 119–132. [Google Scholar] [CrossRef]
- Baroowa, B.; Gogoi, N. Morpho-physiological and yield responses of Black gram (Vigna mungo L.) and Green gram (Vigna radiata L.) genotypes under drought at different growth stages. Res. J. Recent Sci. 2016, 2277, 2502. [Google Scholar]
- Alzarqaa, A.; Roushdy, S.; Alderfasi, A.; Al-Yahya, F.; Dawabah, A. The physiological response of mungbean (Vigna radiata) to water deficit stress and Meloidogyne javanica infection. In Sustainable Irrigation and Drainage V: Management, Technologies and Policies; WIT Press: Billerica, MA, USA, 2014; Volume 185, pp. 89–100. [Google Scholar]
- Abdelaal, K.A.; Attia, K.A.; Alamery, S.F.; El-Afry, M.M.; Ghazy, A.I.; Tantawy, D.S.; Al-Doss, A.A.; El-Shawy, E.S.E.M.; Abu-Elsaoud, A.; Hafez, Y.M. Exogenous application of proline and salicylic acid can mitigate the injurious impacts of drought stress on barley plants associated with physiological and histological characters. Sustainability 2020, 12, 1736. [Google Scholar] [CrossRef] [Green Version]
- Kusvuran, S.; Dasgan, H.Y. Effects of drought stress on physiological and biochemical changes in Phaseolus vulgaris L. Legume Res. Int. J. 2017, 40, 55–62. [Google Scholar]
- Mafakheri, A.; Siosemardeh, A.F.; Bahramnejad, B.; Struik, P.C.; Sohrabi, Y. Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Aust. J. Crop Sci. 2010, 4, 580. [Google Scholar]
- Semida, W.M.; Abdelkhalik, A.; Rady, M.O.; Marey, R.A.; El-Mageed, T.A.A. Exogenously applied proline enhances growth and productivity of drought stressed onion by improving photosynthetic efficiency, water use efficiency and up-regulating osmoprotectants. Sci. Hortic. 2020, 272, 109580. [Google Scholar] [CrossRef]
- Islam, M.R.; Kamal, M.M.; Hossain, M.F.; Hossain, J.; Azam, M.G.; Akhter, M.M.; Hasan, M.K.; Al-Ashkar, I.; Almutairi, K.F.; EL Sabagh, A.; et al. Drought Tolerance in Mung Bean is Associated with the Genotypic Divergence, Regulation of Proline, Photosynthetic Pigment and Water Relation. Phyton Int. J. Exp. Bot. 2022, 92, 955–981. [Google Scholar]
- Kabbadj, A.; Makoudi, B.; Mouradi, M.; Pauly, N.; Frendo, P.; Ghoulam, C. Physiological and biochemical responses involved in water deficit tolerance of nitrogen-fixing Vicia faba. PLoS ONE 2017, 12, e0190284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.N.; AlSolami, M.A.; Basahi, R.A.; Siddiqui, M.H.; Al-Huqail, A.A.; Abbas, Z.K.; Siddiqui, Z.H.; Ali, H.M.; Khan, F. Nitric oxide is involved in nano-titanium dioxide-induced activation of antioxidant defense system and accumulation of osmolytes under water-deficit stress in Vicia faba L. Ecotoxicol. Environ. Saf. 2020, 190, 110152. [Google Scholar] [CrossRef]
- Hare, P.D.; Cress, W.A.; Van Staden, J. Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ. 1998, 21, 535–553. [Google Scholar] [CrossRef]
- McNeil, S.D.; Nuccio, M.L.; Hanson, A.D. Betaines and related osmoprotectants: Targets for metabolic engineering of stress resistance. Plant Physiol. 1999, 120, 945–950. [Google Scholar] [CrossRef] [Green Version]
- Allen, D.J.; McKee, I.F.; Farage, P.K.; Baker, N.R. Analysis of limitations to CO2 assimilation on exposure of leaves of two Brassica napus cultivars to UV-B. Plant Cell Environ. 1997, 20, 633–640. [Google Scholar] [CrossRef]
- Saha, S.; Begum, H.H.; Nasrin, S. Effects of drought stress on growth and accumulation of proline in five rice varieties (Oryza Sativa L.). J. Asiat. Soc. Bangladesh Sci. 2019, 45, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Yaish, M.W. Proline accumulation is a general response to abiotic stress in the date palm tree (Phoenix dactylifera L.). Genet. Mol. Res. 2015, 14, 9943–9950. [Google Scholar] [CrossRef]
- Uddin, S.; Parvin, S.; Awal, M.A. Morpho-physiological aspects of mungbean (Vigna radiata L.) in response to water stress. Int. J. Agric. Sci. Res. 2013, 3, 137–148. [Google Scholar]
- Mirzaei, A.; Naseri, R.; Vafa, P.; Moradi, M. Effects of Drought Stress on Qualitative and Quantitative Traits of Mungbean. World Acad. Sci. Eng. Technol. Int. J. Agric. Biosyst. Eng. 2014, 8, 148–152. [Google Scholar]
- Allahmoradi, P.; Ghobadi, M.; Taherabadi, S.; Taherabadi, S. Physiological aspects of mungbean (Vigna radiata L.) in response to drought stress. In Proceedings of the International Conference on Food Engineering and Biotechnology, IPCBEE, Bangkok, Thailand, 7–9 May 2011; 2011; pp. 272–275. [Google Scholar]
- Ratnasekera, D.; Subhashi, A.P.T. Geographic variation of wild rice; Oryza nivara (SD Sharma and Shastry) populations in Sri Lanka. Int. J. Trop. Agric. 2015, 33, 3235–3238. [Google Scholar]
- Eskandari, H.; Kazemi, K.K. Response of different bread wheat (Triticum aestivum L.) genotypes to post-anthesis water deficit. Not. Sci. Biol. 2010, 2, 49–52. [Google Scholar] [CrossRef] [Green Version]
- Thirunavukkarasu, N.; Sharma, R.; Singh, N.; Shiriga, K.; Mohan, S.; Mittal, S.; Gupta, H.S. Genome wide expression and functional interactions of genes under drought stress in maize. Int. J. Genom. 2017, 2017, 2568706. [Google Scholar]
- Naresh, R.K.; Singh, S.P.; Dwivedi, A.; Kumar, V. Effects of water stress on physiological processes and yield attributes of different mungbean (L.) varieties. Afr. J. Biochem. Res. 2013, 7, 55–62. [Google Scholar]
- Raina, S.K.; Govindasamy, V.; Kumar, M.; Singh, A.K.; Rane, J.; Minhas, P.S. Genetic variation in physiological responses of mungbeans (Vigna radiata (L.) Wilczek) to drought. Acta Physiol. Plant. 2016, 38, 263. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Genotypes | Relative Water Content (%) | Water Saturation Deficit (%) | Water Retention Capacity | Water Uptake Capacity | ||||
|---|---|---|---|---|---|---|---|---|---|
| First Year | Second Year | First Year | Second Year | First Year | Second Year | First Year | Second Year | ||
| Water regime A | G43 | 85.21 b | 83.65 d | 6.09 gh | 6.45 ghi | 4.61 a | 4.56 bcde | 0.30 ij | 0.30 f |
| G88 | 84.33 c | 85.26 c | 6.21 gh | 6.17 i | 4.59 ab | 4.70 a | 0.31 i | 0.31 f | |
| G108 | 85.61 b | 86.62 ab | 6.55 g | 6.69 gh | 4.61 a | 4.63 abcd | 0.31 i | 0.31 f | |
| G141 | 87.29 a | 86.62 ab | 6.39 g | 6.44 ghi | 4.63 a | 4.55 cde | 0.31 i | 0.31 f | |
| G164 | 85.36 b | 85.66 bc | 6.41 g | 6.76 g | 4.61 a | 4.66 ab | 0.30 ij | 0.32 f | |
| G186 | 84.69 bc | 86.23 bc | 6.05 gh | 6.52 ghi | 4.57 ab | 4.56 bcde | 0.31 i | 0.29 f | |
| BU mug 5 | 86.05 ab | 85.61 bc | 6.56 g | 6.31 hi | 4.61 a | 4.52 de | 0.32 i | 0.32 f | |
| Binamoog-8 | 85.61 b | 87.52 a | 6.41 g | 6.35 ghi | 4.55 b | 4.65 abc | 0.30 ij | 0.31 f | |
| Water regime B | G43 | 60.41 h | 61.33 h | 23.41 a | 24.59 a | 3.48 f | 3.59 i | 0.968 a | 0.989 a |
| G88 | 80.22 e | 79.36 f | 12.51 f | 13.76 f | 4.36 c | 4.51 e | 0.501 h | 0.526 e | |
| G108 | 80.62 e | 80.95 e | 14.22 e | 15.84 c | 4.32 c | 4.38 f | 0.562 e | 0.559 d | |
| G141 | 82.61 d | 81.26 e | 14.02 e | 14.40 e | 4.36 c | 4.34 fg | 0.549 f | 0.558 d | |
| G164 | 78.41 f | 79.35 f | 15.11 c | 15.63 c | 4.17 de | 4.26 gh | 0.638 bc | 0.639 b | |
| G186 | 77.36 g | 75.32 g | 15.99 b | 16.71 b | 4.13 e | 4.16 h | 0.629 b | 0.621 b | |
| BU mug 5 | 77.69 g | 76.12 g | 14.81 cd | 15.12 d | 4.30 cd | 4.29 fg | 0.579 d | 0.590 c | |
| Binamoog-8 | 80.28 e | 81.76 e | 14.69 cd | 14.37 e | 4.23 d | 4.39 f | 0.541 g | 0.549 de | |
| CV(%) | 4.36 | 1.91 | 5.16 | 2.53 | 4.82 | 1.76 | 3.81 | 4.47 | |
| SE | 0.8153 | 0.5266 | 0.5381 | 0.2036 | 0.0714 | 0.0552 | 0.0151 | 0.0148 | |
| Photosynthetic Rate (µmol m−2 s−1) | Stomatal Conductance (mmol m−2 s−1) | Transpiration Rate (mmol m−2 s−1 ) | Photosynthetic Water Use Efficiency (Pn/Tr) | Total Chlorophyll Content (mg g−1 ) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatments | Genotypes | First Year | Second Year | First Year | Second Year | First Year | Second Year | First Year | Second Year | First Year | Second Year |
| Water regime A | G43 | 16.24 e | 16.08 e | 0.285 a | 0.286 a | 4.48 bc | 4.65 bc | 3.63 g | 3.46 ef | 3.44 c | 3.37 d |
| G88 | 16.61 b | 16.87 b | 0.285 a | 0.285 a | 4.52 ab | 4.71 ab | 3.67 g | 3.58 e | 3.52 a | 3.63 a | |
| G108 | 16.55 bc | 16.78 bc | 0.286 a | 0.287 a | 4.49 abc | 4.60 c | 3.69 g | 3.65 e | 3.41 c | 3.44 c | |
| G141 | 16.91 a | 17.55 a | 0.286 a | 0.283 ab | 4.50 ab | 4.67 bc | 3.76 g | 3.76 e | 3.47 b | 3.51 b | |
| G164 | 15.88 g | 16.39 d | 0.281 ab | 0.270 cd | 4.56 a | 4.70 ab | 3.48 gh | 3.49 ef | 3.51 a | 3.63 a | |
| G186 | 16.42 d | 16.66 c | 0.283 ab | 0.287 a | 4.61 a | 4.83 a | 3.56 g | 3.45 ef | 3.42 c | 3.35 d | |
| BU mug 5 | 16.53 c | 16.29 d | 0.275 b | 0.277 bc | 4.63 a | 4.69 b | 3.57 g | 3.47 ef | 3.42 c | 3.37 d | |
| Binamoog-8 | 16.11 f | 15.99 e | 0.268 c | 0.267 d | 4.59 a | 4.61 c | 3.51 gh | 3.47 ef | 3.47 b | 3.54 b | |
| Water regime B | G43 | 6.23 o | 5.13 l | 0.176 i | 0.182 i | 0.87 g | 0.85 g | 7.16 f | 6.04 cd | 1.62 h | 1.47 k |
| G88 | 14.91 i | 13.85 g | 0.215 de | 0.219 ef | 1.61 d | 1.60 d | 9.26 a | 9.69 a | 3.21 d | 3.29 e | |
| G108 | 13.26 j | 12.78 h | 0.206 f | 0.214 f | 1.48 f | 1.52 de | 8.96 b | 8.41 ab | 3.04 ef | 3.09 h | |
| G141 | 14.99 h | 15.62 f | 0.221 d | 0.225 e | 1.62 d | 1.61 d | 9.25 a | 9.70 a | 3.16 de | 3.22 f | |
| G164 | 11.34 n | 10.51 k | 0.201 fg | 0.190 h | 1.53 ef | 1.54 de | 7.41 e | 6.41 c | 2.96 fg | 2.99 i | |
| G186 | 11.51 m | 10.79 j | 0.191 gh | 0.189 h | 1.58 de | 1.55 de | 7.28 ef | 6.17 bc | 2.91 g | 2.86 j | |
| BU mug 5 | 12.36 k | 11.87 i | 0.206 f | 0.212 f | 1.49 ef | 1.53 de | 8.30 c | 7.76 b | 2.99 f | 2.99 i | |
| Binamoog-8 | 12.11 l | 11.94 i | 0.197 g | 0.200 g | 1.57 de | 1.61 de | 7.71 d | 7.42 b | 3.06 e | 3.15 g | |
| CV(%) | 3.29 | 1.77 | 5.26 | 2.03 | 4.81 | 1.60 | 4.37 | 2.63 | 3.91 | 1.62 | |
| SE | 0.0631 | 0.0776 | 0.00416 | 0.00339 | 0.117 | 0.266 | 0.231 | 0.536 | 0.068 | 0.140 | |
| Treatments | Genotypes | Plant Height (cm) | No. of Branches Plant−1 | No. of Pods Plant−1 | Pod Length (cm) | No. of Seeds Pod−1 | 1000-Seeds Weight (g) | Seed Yield Plant−1 (g) |
|---|---|---|---|---|---|---|---|---|
| Water regime A | G43 | 52.85 fg | 2.25 c | 16.65 g | 7.51 abc | 6.93 cd | 45.80 c | 12.05 d |
| G88 | 58.45 de | 2.75 b | 22.63 bc | 7.87 a | 7.20 c | 40.25 d | 14.48 a | |
| G108 | 61.08 cd | 3.13 a | 21.88 cd | 7.88 a | 6.93 cd | 51.37 a | 13.32 b | |
| G141 | 76.19 a | 2.75 b | 28.88 a | 7.48 abcd | 6.90 cd | 37.79 ef | 13.06 b | |
| G164 | 69.08 b | 2.67 b | 23.25 b | 7.25 cdef | 8.86 a | 37.43 f | 12.81 b | |
| G186 | 52.50 fg | 3.00 a | 16.75 fg | 7.04 efg | 6.98 cd | 48.92 b | 12.72 bc | |
| BU mug 5 | 57.64 de | 2.68 b | 18.00 e | 7.73 ab | 6.13 ef | 50.69 ab | 14.17 a | |
| Binamoog-8 | 51.29 fg | 2.68 b | 21.00 d | 7.38 bcde | 7.20 c | 46.23 c | 13.05 b | |
| Water regime B | G43 | 35.29 j | 1.75 d | 13.13 i | 5.99 h | 5.28 h | 40.76 d | 6.18 g |
| G88 | 54.84 ef | 2.60 b | 21.30 d | 7.63 abc | 6.80 cd | 39.86 de | 12.11 cd | |
| G108 | 53.61 f | 2.71 b | 18.75 e | 7.62 abc | 6.51de | 49.82 ab | 11.11 e | |
| G141 | 68.96 b | 2.58 b | 28.50 a | 6.97 fg | 6.55 de | 37.42 f | 11.86 d | |
| G164 | 63.09 c | 2.18 c | 18.88 e | 6.95 fg | 7.35 b | 37.05 f | 10.29 f | |
| G186 | 49.24 gh | 2.58 b | 15.75 gh | 6.63 g | 6.51 de | 45.44 c | 10.06 f | |
| BU mug 5 | 45.98 hi | 2.18 c | 15.18 h | 7.07 def | 5.72 fgh | 49.62 ab | 9.81 f | |
| Binamoog-8 | 44.55 i | 2.19 c | 17.88 ef | 6.89 fg | 6.47 de | 44.75 c | 10.39 f | |
| CV(%) | 3.99 | 4.62 | 3.57 | 3.40 | 4.80 | 3.20 | 3.36 | |
| SE | 1.187 | 0.0678 | 0.4103 | 0.1422 | 0.1847 | 0.8128 | 0.2268 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamanna, T.; Islam, M.M.; Chaity, A.R.; Shams, S.-N.-U.; Rasel, M.A.; Haque, M.M.; Miah, M.G.; Alamri, S.; Murata, Y. Water Relation, Gas Exchange Characteristics and Yield Performance of Selected Mungbean Genotypes under Low Soil Moisture Condition. Agronomy 2023, 13, 1068. https://doi.org/10.3390/agronomy13041068
Tamanna T, Islam MM, Chaity AR, Shams S-N-U, Rasel MA, Haque MM, Miah MG, Alamri S, Murata Y. Water Relation, Gas Exchange Characteristics and Yield Performance of Selected Mungbean Genotypes under Low Soil Moisture Condition. Agronomy. 2023; 13(4):1068. https://doi.org/10.3390/agronomy13041068
Chicago/Turabian StyleTamanna, Tahmina, Md. Moshiul Islam, Arpita Roy Chaity, Shahjadi-Nur-Us Shams, Md. Asadujjaman Rasel, M. Moynul Haque, Md. Giashuddin Miah, Saud Alamri, and Yoshiyuki Murata. 2023. "Water Relation, Gas Exchange Characteristics and Yield Performance of Selected Mungbean Genotypes under Low Soil Moisture Condition" Agronomy 13, no. 4: 1068. https://doi.org/10.3390/agronomy13041068