
Three-Year Survey of Fusarium Multi-Metabolites/Mycotoxins Contamination in Wheat Samples in Potentially Epidemic FHB Conditions

Abstract
:1. Introduction
2. Materials and Methods
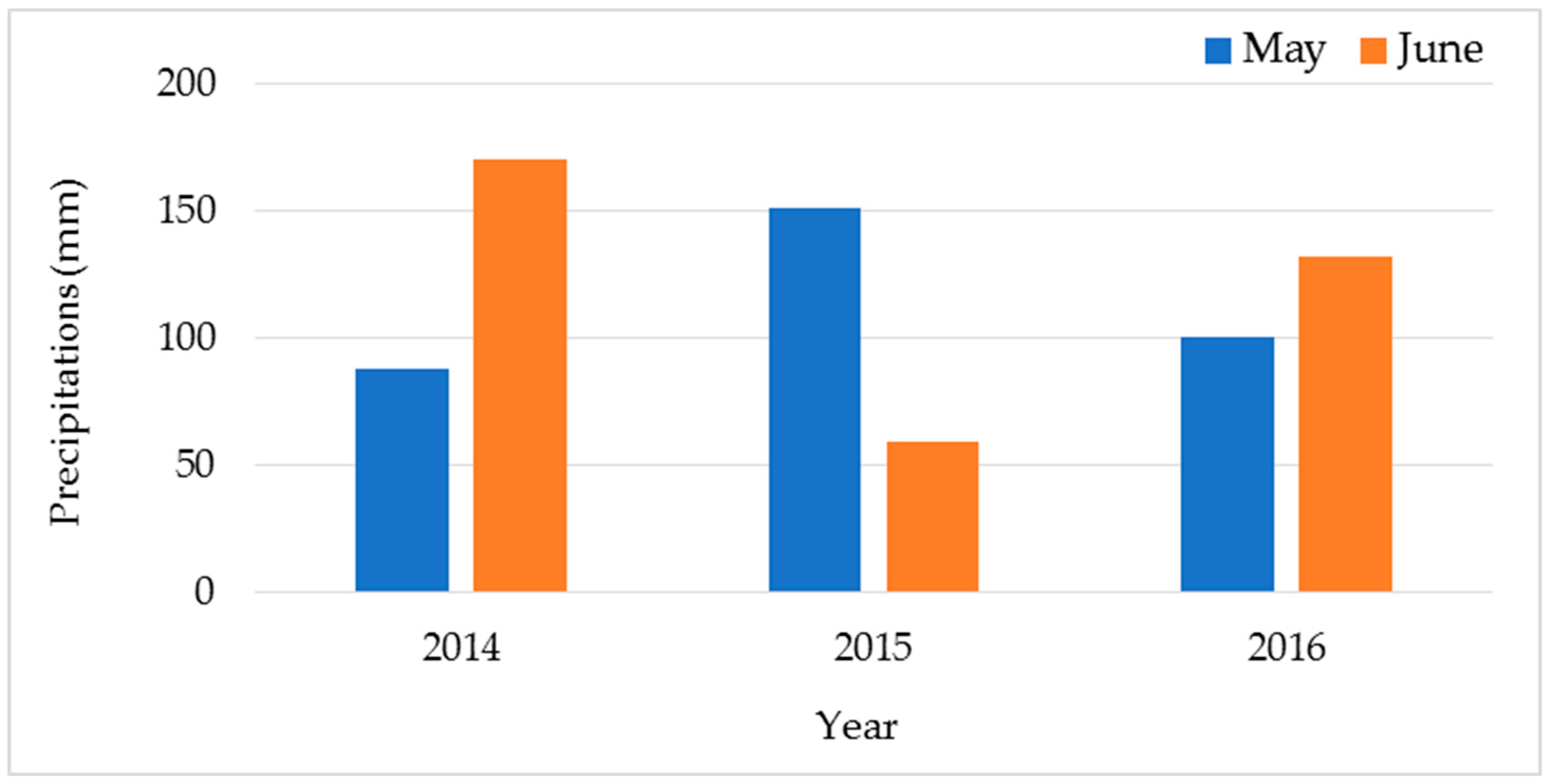
2.1. Wheat Material and Field Conditions
2.2. Inoculum Production and Inoculation
2.3. Disease Assessment
2.4. Mycotoxin Analysis
2.5. Statistical Analysis
3. Results
3.1. Fusarium Head Blight Assessment
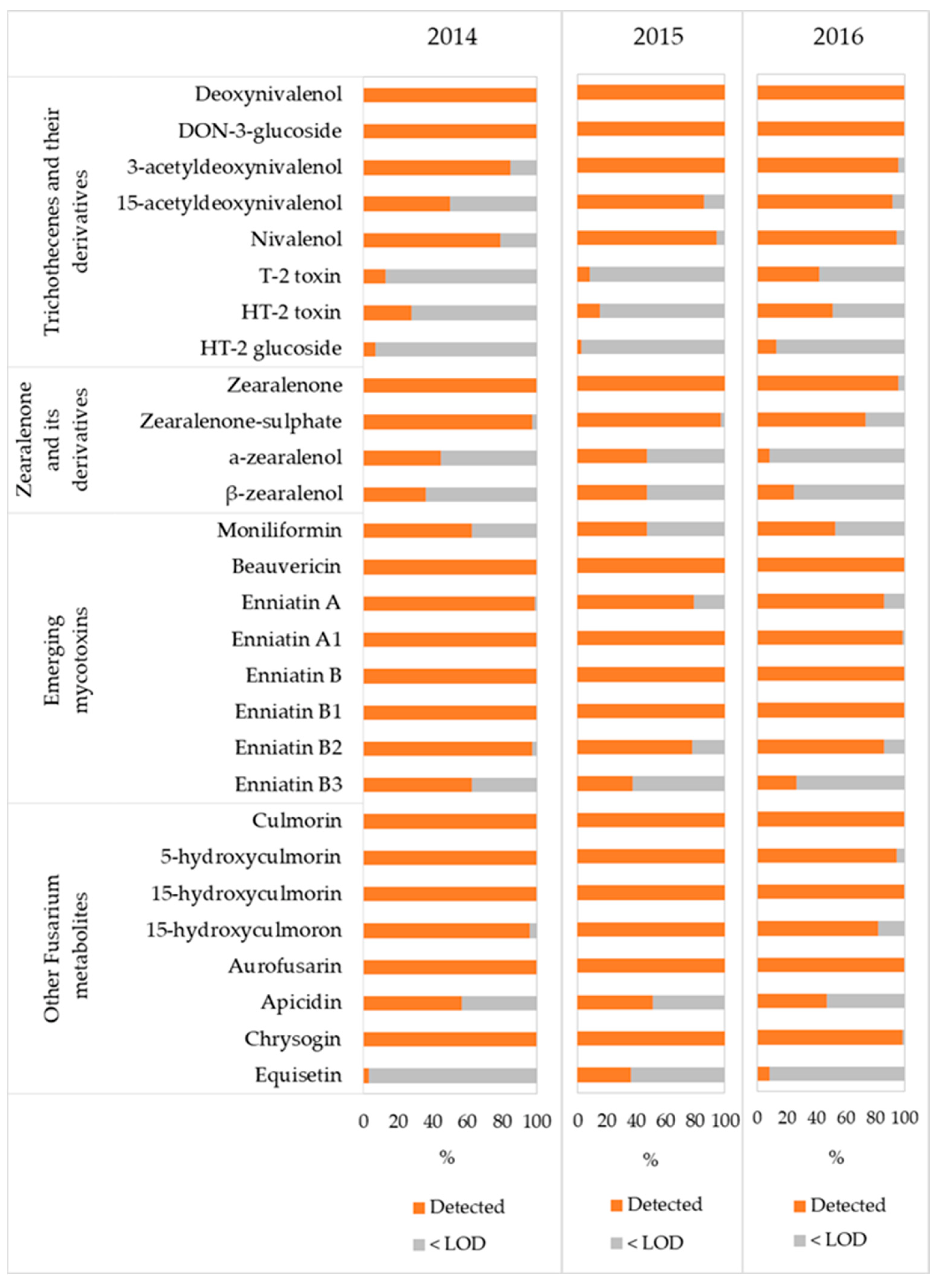
3.2. Trithothecenes and Their Derivatives
3.3. Zearelenone, Zearelenone-Sulphate, α- and β-Zearalenol
3.4. Moniliformin, Beauvericin and Enniatins
3.5. Other Fusarium Metabolites
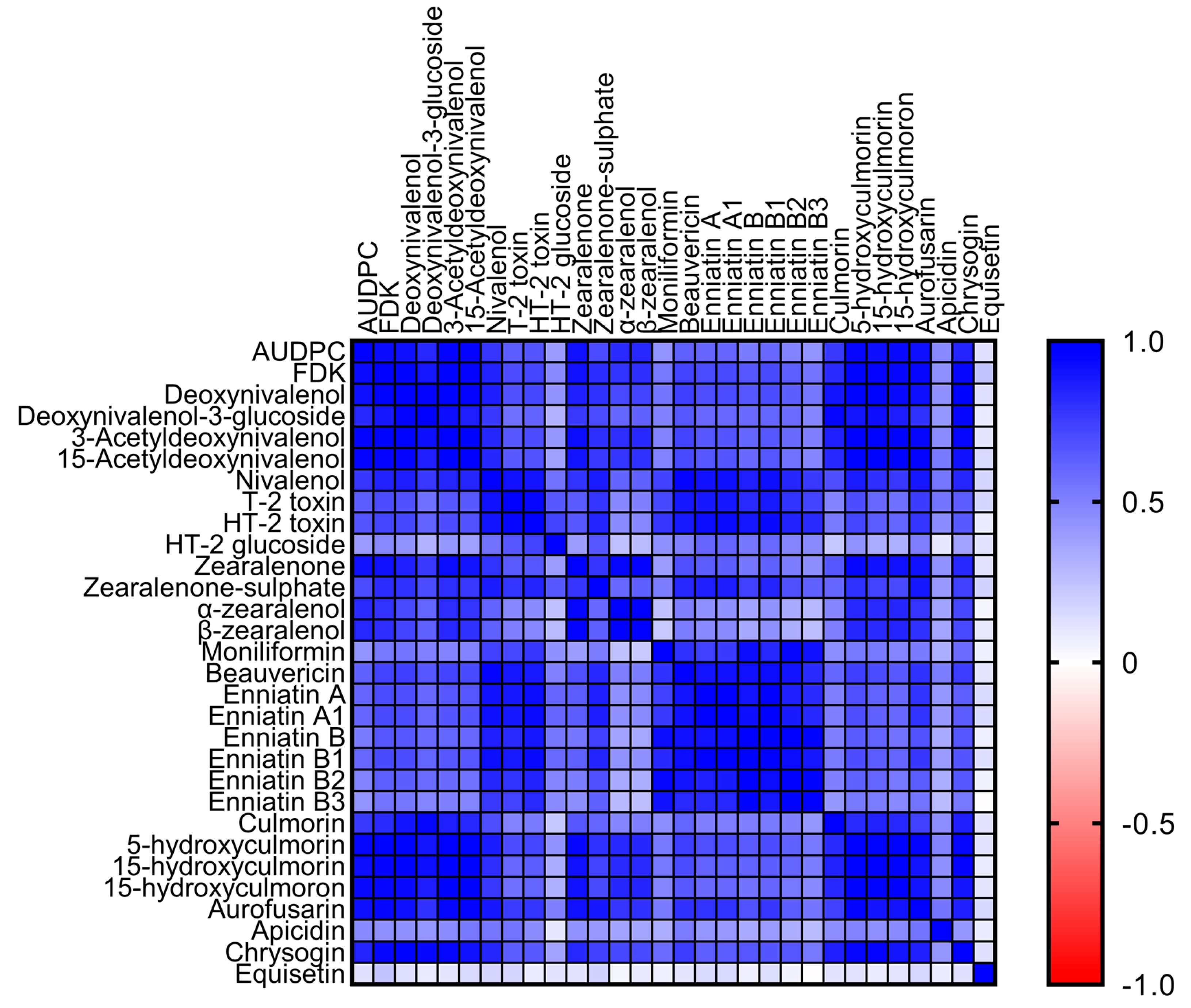
3.6. Co-Occurrence and Correlation of Fusarium Mycotoxins/Metabolites
4. Discussion
4.1. Trichothecenes and Their Modified Forms
4.2. Zearalenone and Its Derivatives
4.3. Emerging Mycotoxins
4.4. Other Fusarium Metabolites
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Spanic, V.; Marcek, T.; Abicic, I.; Sarkanj, B. Effects of Fusarium head blight on wheat grain and malt infected by Fusarium culmorum. Toxins 2018, 10, 17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Boyle, K.; Brûlé-Babel, A.L.; Fedak, G.; Gao, P.; Robleh Djama, Z.; Polley, B.; Cuthbert, R.D.; Randhawa, H.S.; Jiang, F.; et al. Genetic characterization of multiple components contributing to Fusarium Head Blight Resistance of FL62R1, a canadian bread wheat developed using systemic breeding. Front. Plant Sci. 2020, 26, 11–580833. [Google Scholar] [CrossRef] [PubMed]
- Mesterhazy, A.; Toth, B.; Varga, M.; Bartok, T.; Szabo-Hever, A.; Farady, L.; Lehoczki-Krsjak, S. Role of fungicides, application of nozzle types, and the resistance level of wheat varieties in the control of Fusarium head blight and deoxynivalenol. Toxins 2011, 3, 1453–1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spanic, V.; Viljevac Vuletic, M.; Horvat, D.; Sarkanj, B.; Drezner, G.; Zdunic, Z. Changes in antioxidant system during grain development of wheat (Triticum aestivum L.) and relationship with protein composition under FHB stress. Pathogens 2020, 9, 17. [Google Scholar] [CrossRef] [Green Version]
- Wilde, F.; Miedaner, T. Selection for Fusarium head blight resistance in early generations reduces the deoxynivalenol (DON) content in grain of winter and spring wheat. Plant Breed 2006, 125, 96–98. [Google Scholar] [CrossRef]
- Sunic, K.; Kovac, T.; Loncaric, A.; Babic, J.; Sulyok, M.; Krska, R.; Drezner, G.; Spanic, V. Fusarium secondary metabolite content in naturally produced and artificially provoked FHB pressure in winter wheat. Agronomy 2021, 11, 2239. [Google Scholar] [CrossRef]
- Bushnell, W.R.; Hazen, B.E.; Pritsch, C. Histology and physiology of Fusarium head blight. In Fusarium Head Blight of Wheat and Barley, 1st ed.; Leonard, K.J., Bushnell, W.R., Eds.; The American Phytopathological Society: St. Paul, MN, USA, 2003; Chapter 3; pp. 44–83. [Google Scholar]
- Rodrigues, I.; Naehrer, K. A three-year survey on the worldwide occurrence of mycotoxins in feedstuffs and feed. Toxins 2012, 4, 663–675. [Google Scholar] [CrossRef]
- Cheli, F.; Pinotti, L.; Rossi, L.; Dell’Orto, V. Effect of milling procedures on mycotoxin distribution in wheat fractions: A review. LWT Food Sci. Technol. 2013, 54, 307–314. [Google Scholar] [CrossRef]
- Shank, R.A.; Foroud, N.A.; Hazendonk, P.; Eudes, F.; Blackwell, B.A. Current and future experimental strategies for structural analysis of trichothecene mycotoxins—A prospectus. Toxins 2011, 3, 1518–1553. [Google Scholar] [CrossRef] [Green Version]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [Green Version]
- Lemos, A.C.; de Borba, V.S.; Badiale-Furlong, E. The impact of wheat-based food processing on the level of trichothecenes and their modified forms. Trends Food Sci. Technol. 2021, 111, 89–99. [Google Scholar] [CrossRef]
- Stanciu, O.; Juan, C.; Berrada, H.; Miere, D.; Loghin, F.; Mañes, J. Study on trichothecene and zearalenone presence in Romanian wheat relative to weather conditions. Toxins 2019, 11, 163. [Google Scholar] [CrossRef] [Green Version]
- Commission Regulation (EC) No 1881/2006 of 19 December 2006 on the Setting Maximum Levels for Certain Contaminants in Foodstuffs. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32006R1881 (accessed on 28 January 2023).
- Commission Recommendation (ER) No 2013/165/EU of 27 March 2013 on the Presence of T-2 and HT-2 Toxin in Cereals and Cereal Products. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32013H0165 (accessed on 28 January 2023).
- EFSA. Panel on Contaminants in the Food Chain. Scientific Opinion on risks for animal and public health related to the presence of nivalenol in food and feed. EFSA J 2013, 11, 3262. [Google Scholar] [CrossRef]
- Berthiller, F.; Crews, C.; Dall’Asta, C.; De Saeger, S.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Seefelder, W.; Speijers, G.; Strokaet, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef] [PubMed]
- Ekwomadu, T.I.; Akinola, S.A.; Mwanza, M. Fusarium mycotoxins, their metabolites (free, emerging, and masked), food safety concerns, and health impacts. Int. J. Environ. Res. Public Health 2021, 18, 11741. [Google Scholar] [CrossRef]
- Broekaert, N.; Devreese, M.; de Baere, S.; de Backer, P.; Croubels, S. Modified Fusarium mycotoxins unmasked: From occurrence in cereals to animal and human excretion. Food Chem. Toxicol. 2015, 80, 17–31. [Google Scholar] [CrossRef]
- Iqbal, S.Z. Mycotoxins in food, recent development in food analysis and future challenges: A review. Curr. Opin. Food Sci. 2021, 42, 237–247. [Google Scholar] [CrossRef]
- Berthiller, F.; Dall’Asta, C.; Schuhmacher, R.; Lemmens, M.; Adam, G.; Krska, R. Masked mycotoxins: Determination of a deoxynivalenolglucoside in artificially and naturally contaminated wheat by liquid chromatography—Tandem mass spectrometry. J. Agric. Food Chem. 2005, 53, 3421–3425. [Google Scholar] [CrossRef]
- Borzekowski, A.; Drewitz, T.; Keller, J.; Pfeifer, D.; Kunte, H.J.; Koch, M.; Rohn, S.; Maul, R. Biosynthesis and characterization of zearalenone-14-sulfate, zearalenone-14-glucoside and zearalenone-16-glucoside Using Common Fungal Strains. Toxins 2018, 10, 104. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Woo, S.Y.; Tian, F.; Song, J.; Michlmayr, H.; Kim, J.B.; Chun, H.S. Occurrence of deoxynivalenol, nivalenol, and their glucosides in Korean market foods and estimation of their population exposure through food consumption. Toxins 2020, 12, 89. [Google Scholar] [CrossRef] [Green Version]
- Ovando-Martínez, M.; Ozsisli, B.; Anderson, J.; Whitney, K.; Ohm, J.B.; Simsek, S. Analysis of deoxynivalenol and deoxynivalenol-3-glucoside in hard red spring wheat inoculated with Fusarium Graminearum. Toxins 2013, 5, 2522–2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spanic, V.; Katanic, Z.; Sulyok, M.; Krska, R.; Puskas, K.; Vida, G.; Drezner, G.; Sarkanj, B. Multiple fungal metabolites including mycotoxin formation in naturally infected and Fusarium inoculated wheat samples. Microorganisms 2020, 8, 578. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, H.; Ohmichi, K.; Sakamoto, S.; Sago, Y.; Kushiro, M.; Nagashima, H.; Yoshida, M.; Nakajima, T. Detection of a new Fusarium masked mycotoxin in wheat grain by high-resolution LC–Orbitrap™ MS. Food Addit. Contam. A 2011, 28, 1447–1456. [Google Scholar] [CrossRef] [PubMed]
- Prosperini, A.; Berrada, H.; Ruiz, M.J.; Caloni, F.; Coccini, T.; Spicer, L.J.; Perego, M.C.; Lafranconi, A. A Review of the mycotoxin enniatin B. Front. Public Health 2017, 5, 304. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Blanco, C.; Font, G.; Ruiz, M.J. Interaction effects of enniatin B, deoxinivalenol and alternariol in Caco-2 cells. Toxicol. Lett. 2016, 241, 38–48. [Google Scholar] [CrossRef]
- Nazari, F.; Sulyok, M.; Kobarfard, F.; Yazdanpanah, H.; Krska, R. Evaluation of emerging Fusarium mycotoxins beauvericin, enniatins, fusaproliferin and moniliformin in domestic rice in Iran. Iran J. Pharm. Res. 2015, 14, e125357. [Google Scholar] [CrossRef]
- Wu, Q.; Patocka, J.; Nepovimova, E.; Kuca, K. A Review on the synthesis and bioactivity aspects of beauvericin, a Fusarium Mycotoxin. Front. Pharmacol. 2018, 9, 1338. [Google Scholar] [CrossRef] [Green Version]
- Kuang, Q.-X.; Lei, L.-R.; Li, Q.-Z.; Peng, W.; Wang, Y.-M.; Dai, Y.-F.; Wang, D.; Gu, Y.-C.; Deng, Y.; Guo, D.-L. Investigation of the anti-inflammatory activity of fusaproliferin analogues guided by transcriptome analysis. Front. Pharmacol. 2022, 13, 881182. [Google Scholar] [CrossRef]
- Ceranic, A.; Svoboda, T.; Berthiller, F.; Sulyok, M.; Samson, J.M.; Güldener, U.; Schuhmacher, R.; Adam, G. Identification and functional characterization of the gene cluster responsible for fusaproliferin biosynthesis in Fusarium proliferatum. Toxins 2021, 13, 468. [Google Scholar] [CrossRef]
- Bertuzzi, T.; Giorni, P.; Rastelli, S.; Vaccino, P.; Lanzanova, C.; Locatelli, S. Co-Occurrence of moniliformin and regulated Fusarium toxins in maize and wheat grown in Italy. Molecules 2020, 25, 2440. [Google Scholar] [CrossRef]
- Bouafifssa, Y.; Manyes, L.; Rahouti, M.; Mañes, J.; Berrada, H.; Zinedine, A.; Fernández-Franzón, M. Multi-occurrence of twenty mycotoxins in pasta and a risk assessment in the Moroccan population. Toxins 2018, 10, 432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahdjoubi, C.K.; Arroyo-Manzanares, N.; Hamini-Kadar, N.; García-Campaña, A.M.; Mebrouk, K.; Gámiz-Gracia, L. Multi-mycotoxin occurrence and exposure assessment approach in foodstuffs from Algeria. Toxins 2020, 12, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siuda, R.; Grabowski, A.; Lenc, L.; Ralcewicz, M.; Spychaj-Fabisiak, E. Influence of the degree of fusariosis on technological traits of wheat grain. Int. J. Food Sci. Technol. 2010, 45, 2596–2604. [Google Scholar] [CrossRef]
- Spanic, V.; Dvojkovic, K.; Babic, J.; Drezner, G.; Zdunic, Z. Fusarium head blight infestation in relation to winter wheat end-use quality—A three-year study. Agronomy 2021, 11, 1648. [Google Scholar] [CrossRef]
- Malachova, A.; Dzuman, Z.; Veprikova, Z.; Vaclavikova, M.; Zachariasova, M.; Hajslova, J. Deoxynivalenol, deoxynivalenol-3-glucoside, and enniatins: The major mycotoxins found in cereal-based products on the Czech market. J. Agric. Food Chem. 2011, 59, 12990–12997. [Google Scholar] [CrossRef] [PubMed]
- Ryu, D.; Hanna, M.A.; Bullerman, L.B. Stability of zearalenone during extrusion of corn grits. J. Food Prot. 1999, 62, 1482–1484. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A.E.; Manandhar, G.; Plattner, R.D.; Maragos, C.M.; Shrestha, K.; McCormick, S.P. Occurrence of Fusarium species and mycotoxins in Nepalese maize and wheat and the effect of traditional processing methods on mycotoxin levels. J. Agric. Food Chem. 2000, 48, 1377–1383. [Google Scholar] [CrossRef]
- Spanic, V.; Lemmens, M.; Drezner, G. Variability of components of fusarium head blight resistance among wheat genotypes. Cereal. Res. Commun. 2013, 41, 420–430. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/STAT 9.2 Users Guide; SAS Inc.: Cary, NC, USA, 2009. [Google Scholar]
- Motulsky, H.J. Analyzing Data with GraphPad Prism; GraphPad Software Inc.: San Diego, CA, USA, 1999. [Google Scholar]
- Awuchi, C.G.; Ondari, E.N.; Ogbonna, C.U.; Upadhyay, A.K.; Baran, K.; Okpala, C.O.R.; Korzeniowska, M.; Guiné, R.P.F. Mycotoxins affecting animals, foods, humans, and plants: Types, occurrence, toxicities, action mechanisms, prevention, and detoxification strategies—A Revisit. Foods 2021, 10, 1279. [Google Scholar] [CrossRef]
- Wegulo, S. Factors influencing deoxinivalenol accumulation in small grain cereals. Toxins 2012, 4, 1157–1180. [Google Scholar] [CrossRef]
- Alizadeh, A.; Braber, S.; Akbari, P.; Kraneveld, A.; Garssen, J.; Fink-Gremmels, J. Deoxynivalenol and its modified forms: Are there major differences? Toxins 2016, 8, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toxicology and Occurrence of Nivalenol, Fusarenon X, Diacetoxyscirpenol, Neosolaniol and 3- and 15-Acetyldeoxynivalenol: A Review of Six Trichothecenes. RIVM Report 388802024/2002. Available online: https://www.rivm.nl/bibliotheek/rapporten/388802024.pdf (accessed on 15 January 2023).
- Sugita-Konishi, Y.; Park, B.J.; Kobayashi-Hattori, K.; Tanaka, T.; Chonan, T.; Yoshikawa, K.; Kumagai, S. Effect of cooking process on the deoxynivalenol content and its subsequent cytotoxicity in wheat products. Biosci. Biotechnol. Biochem. 2006, 70, 1764–1768. [Google Scholar] [CrossRef] [Green Version]
- Castañares, E.; Albuquerque, D.R.; Dinolfo, M.I.; Pinto, V.F.; Patriarca, A.; Stenglein, S.A. Trichothecene genotypes and production profiles of Fusarium graminearum isolates obtained from barley cultivated in Argentina. Int. J. Food Microbiol. 2014, 179, 57–63. [Google Scholar] [CrossRef]
- Turner, P.C. Deoxynivalenol and nivalenol occurrence and exposure assessment. World Mycotoxin J. 2010, 3, 315–321. [Google Scholar] [CrossRef]
- Nathanail, A.V.; Syvähuoko, J.; Malachová, A.; Jestoi, M.; Varga, E.; Michlmayr, H.; Adam, G.; Sieviläinen, E.; Berthiller, F.; Peltonen, K. Simultaneous determination of major type A and B trichothecenes, zearalenone and certain modified metabolites in Finnish cereal grains with a novel liquid chromatography-tandem mass spectrometric method. Anal. Bioanal. Chem. 2015, 407, 4745–4755. [Google Scholar] [CrossRef] [Green Version]
- Berthiller, F.; Krska, R.; Domig, K.J.; Kneifel, W.; Juge, N.; Schuhmacher, R.; Adam, G. Hydrolytic fate of deoxynivalenol-3-glucoside during digestion. Toxicol. Lett. 2011, 206, 264–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spanic, V.; Zdunic, Z.; Drezner, G.; Sarkanj, B. The pressure of Fusarium disease and its relation with mycotoxins in the wheat grain and malt. Toxins 2019, 11, 198. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Yin, X.; Dong, J.; Yang, Q.; Wu, Y.; Gong, Z. Transcriptome analysis of Caco-2 cells upon the exposure of mycotoxin deoxynivalenol and its acetylated derivatives. Toxins 2021, 13, 167. [Google Scholar] [CrossRef] [PubMed]
- Springler, A.; Hessenberger, S.; Reisinger, N.; Kern, C.; Nagl, V.; Schatzmayr, G.; Mayer, E. Deoxynivalenol and its metabolite deepoxy-deoxynivalenol: Multi-parameter analysis for the evaluation of cytotoxicity and cellular effects. Mycotoxin. Res. 2017, 33, 25–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagashima, H. Deoxynivalenol and nivalenol toxicities in cultured cells: A review of comparative studies. Food Saf. 2018, 6, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Zingales, V.; Fernández-Franzón, M.; Ruiz, M.-J. Occurrence, mitigation and in vitro cytotoxicity of nivalenol, a type B trichothecene mycotoxin–Updates from the last decade (2010–2020). Food Chem. Toxicol. 2021, 152, 112182. [Google Scholar] [CrossRef] [PubMed]
- Bryła, M.; Pierzgalski, A.; Zapaśnik, A.; Uwineza, P.A.; Ksieniewicz-Woźniak, E.; Modrzewska, M.; Waśkiewicz, A. Recent research on Fusarium mycotoxins in maize-A Review. Foods 2022, 11, 3465. [Google Scholar] [CrossRef]
- EFSA. Panel on Contaminants in the Food Chain. Appropriateness to set a group health based guidance value for T2 and HT2 toxin and its modified forms. EFSA J. 2017, 15, e04655. [Google Scholar] [CrossRef]
- Hjelkrem, A.G.R.; Aamot, H.U.; Brodal, G.; Strand, E.C.; Torp, T.; Edwards, S.G.; Dill-Macky, R.; Hofgaard, I.S. HT-2 and T-2 toxins in Norwegian oat grains related to weather conditions at different growth stages. Eur. J. Plant Pathol. 2018, 151, 501–514. [Google Scholar] [CrossRef] [Green Version]
- Poor, M.; Kunsagi-Mate, S.; Sali, N.; Koszegi, T.; Szente, L.; Peles-Lemli, B. Interactions of zearalenone with native and chemically modified cyclodextrins and their potential utilization. J. Photochem. Photobiol. B 2015, 151, 63–68. [Google Scholar] [CrossRef]
- Tan, H.; Zhou, H.; Guo, T.; Zhang, Y.; Ma, L. Zein-bound zearalenone: A hidden mycotoxin found in maize and maize-products. Food Control 2021, 124, 107903. [Google Scholar] [CrossRef]
- Keller, J.; Borzekowski, A.; Haase, H.; Menzel, R.; Rueß, L.; Koch, M. Toxicity assay for citrinin, zearalenone and zearalenone-14-sulfate using the nematode Caenorhabditis elegans as Model Organism. Toxins 2018, 10, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez Pereyra, M.L.; Sulyok, M.; Baralla, V.; Dalcero, A.M.; Krska, R.; Chulze, S.N.; Cavaglieri, L.R. Evaluation of zearalenone, α-zearalenol, β-zearalenol, zearalenone 4-sulfate and β-zearalenol 4-glucoside levels during the ensiling process. World Mycotoxin J. 2014, 7, 291–295. [Google Scholar] [CrossRef]
- Shkhaliyeva, I.; Teker, T.; Albayrak, G. Masked mycotoxins of deoxynivalenol and zearalenone—Unpredicted toxicity. Biomed. J. Sci. Tech. Res. 2020, 29, 22288–22293. [Google Scholar] [CrossRef]
- Vogelgsang, S.; Musa, T.; Bänziger, I.; Kägi, A.; Bucheli, T.D.; Wettstein, F.E.; Pasquali, M.; Forrer, H.R. Fusarium mycotoxins in Swiss wheat: A survey of growers’ samples between 2007 and 2014 shows strong year and minor geographic effects. Toxins 2017, 9, 246. [Google Scholar] [CrossRef] [Green Version]
- Ben Salah-Abbès, J.; Belgacem, H.; Ezzdini, K.; Abdel-Wahhab, M.A.; Abbès, S. Zearalenone nephrotoxicity: DNA fragmentation, apoptotic gene expression and oxidative stress protected by Lactobacillus plantarum MON03. Toxicon 2020, 175, 28–35. [Google Scholar] [CrossRef]
- Abid-Essefi, S.C.; Bouaziz, E.; El Golli-Bennour, Z.; Ouanes, H.B. Comparative study of toxic effects of zearalenone and its two major metabolites α-zearalenol and β-zearalenol on cultured human Caco-2 cells. J. Biochem. Mol. Toxicol. 2009, 23, 233–243. [Google Scholar] [CrossRef]
- Hietaniemi, V.; Ramo, S.; Yli-Mattila, T.; Jestoi, M.; Peltonen, S.; Kartio, M.; Sievilainen, E.; Koivisto, T.; Parikka, P. Updated survey of Fusarium species and toxins in Finnish cereal grains. Food Addit. Contam. A 2016, 33, 831–848. [Google Scholar] [CrossRef]
- Waśkiewicz, A.; Łukasz, S. Mycotoxins biosynthesized by plant-derived Fusarium isolates. Arh. Hig. Rada Toksikol. 2013, 63, 437–446. [Google Scholar] [CrossRef] [Green Version]
- Logrieco, A.; Moretti, A.; Castella, G.; Kostecki, M.; Golinski, P.; Ritieni, A.; Chelkowski, J. Beauvericin production by Fusarium species. Appl. Environ. Microbiol. 1998, 64, 3084–3088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Křížová, L.; Dadáková, K.; Dvořáčková, M.; Kašparovský, T. Feedborne mycotoxins beauvericin and enniatins and livestock animals. Toxins 2021, 13, 32. [Google Scholar] [CrossRef] [PubMed]
- Agahi, F.; Álvarez-Ortega, N.; Font, G.; Juan-García, A.; Juan, C. Oxidative stress, glutathione, and gene expression as key indicators in SH-SY5Y cells exposed to zearalenone metabolites and beauvericin. Toxicol. Lett. 2020, 334, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Cimbalo, A.; Alonso-Garrido, M.; Font, G.; Frangiamone, M.; Manyes, L. Transcriptional changes after enniatins A, A1, B and B1 ingestion in rat stomach, liver, kidney and lower intestine. Foods 2021, 10, 1630. [Google Scholar] [CrossRef]
- Reisinger, N.; Schürer-Waldheim, S.; Mayer, E.; Debevere, S.; Antonissen, G.; Sulyok, M.; Nagl, V. Mycotoxin occurrence in maize silage—A neglected risk for bovine gut health? Toxins 2019, 11, 577. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Scientific Opinion on the risks to human and animal health related to the presence of beauvericin and enniatins in food and feed. EFSA J. 2014, 12, 3802. [Google Scholar] [CrossRef]
- Weber, J.; Vaclavikova, M.; Wiesenberger, G.; Haider, M.; Hametner, C.; Fröhlich, J.; Berthiller, F.; Adam, G.; Mikula, H.; Fruhmann, P. Chemical synthesis of culmorin metabolites and their biologic role in culmorin and acetyl-culmorin treated wheat cells. Org. Biomol. Chem. 2018, 16, 2043–2048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlig, S.; Eriksen, G.S.; Hofgaard, I.S.; Krska, R.; Beltrán, E.; Sulyok, M. Faces of a changing climate: Semi-quantitative multi-mycotoxin analysis of grain grown in exceptional climatic conditions in Norway. Toxins 2013, 5, 1682–1697. [Google Scholar] [CrossRef]
- Wipfler, R.; McCormick, S.P.; Proctor, R.; Teresi, J.; Hao, G.; Ward, T.; Alexander, N.; Vaughan, M.M. Synergistic phytotoxic effects of culmorin and trichothecene mycotoxins. Toxins 2019, 11, 555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woelflingseder, L.; Warth, B.; Vierheilig, I.; Schwartz-Zimmermann, H.; Hametner, C.; Nagl, V.; Novak, B.; Sarkanj, B.; Berthiller, F.; Adam, G. The Fusarium metabolite culmorin suppresses the in vitro glucuronidation of deoxynivalenol. Arch. Toxicol. 2019, 93, 1729–1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Streit, E.; Schwab, C.; Sulyok, M.; Naehrer, K.; Krska, R.; Schatzmayr, G. Multi-mycotoxin screening reveals the occurrence of 139 different secondary metabolites in feed and feed ingredients. Toxins 2013, 5, 504–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarolim, K.; Wolters, K.; Woelflingseder, L.; Pahlke, G.; Beisl, J.; Puntscher, H.; Braun, D.; Sulyok, M.; Warth, B.; Marko, D. The secondary Fusarium metabolite aurofusarin induces oxidative stress, cytotoxicity and genotoxicity in human colon cells. Toxicol. Lett. 2018, 284, 170–183. [Google Scholar] [CrossRef]
- Khoshal, A.K.; Novak, B.; Martin, P.G.P.; Jenkins, T.; Neves, M.; Schatzmayr, G.; Oswald, I.P.; Pinton, P. Co-occurrence of DON and emerging mycotoxins in worldwide finished pig feed and their combined toxicity in intestinal cells. Toxins 2019, 11, 727. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
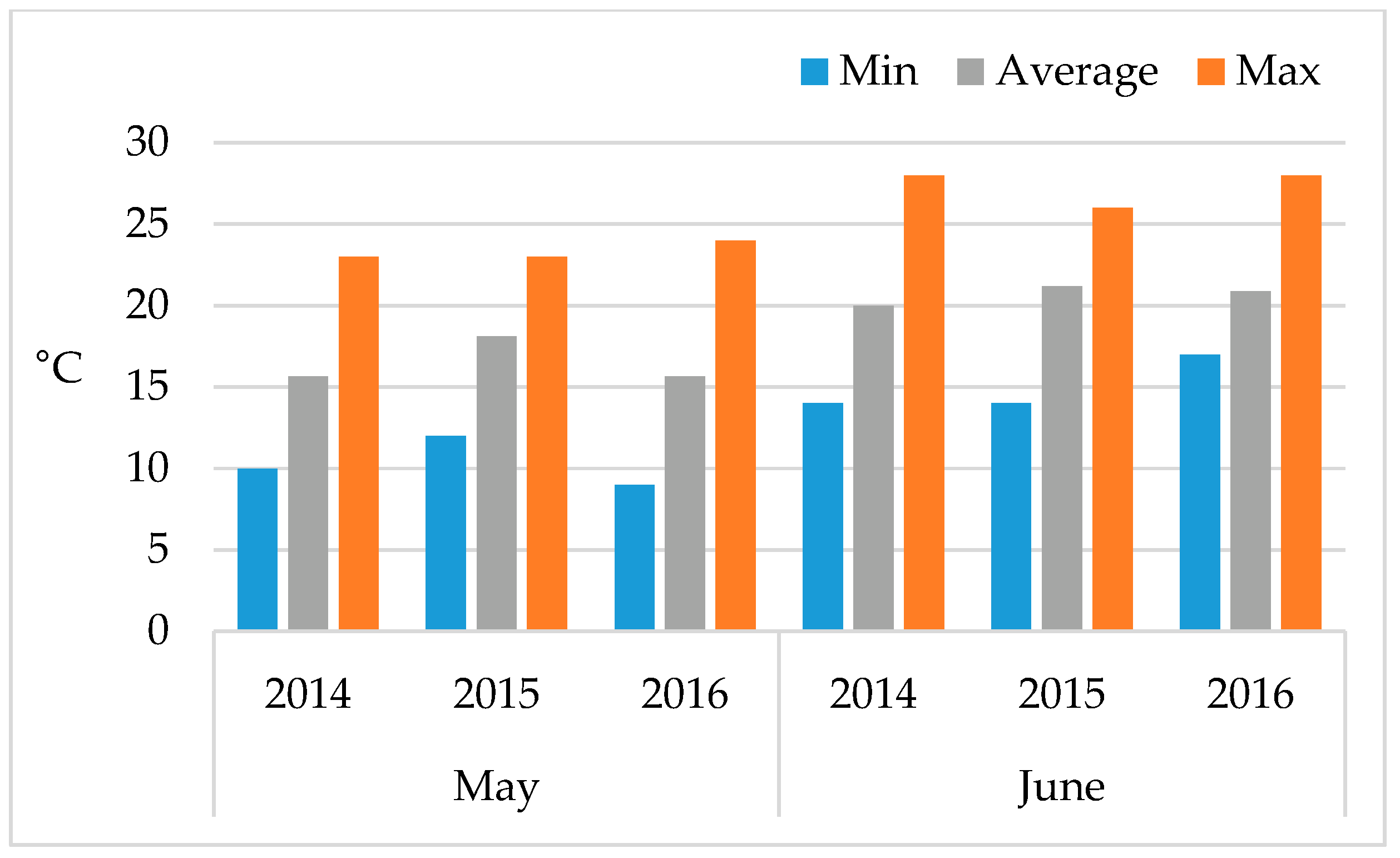{kind=link}
{kind=link}
{kind=link}
| Year (Y) | Genotype (G) | G × Y | |||||
|---|---|---|---|---|---|---|---|
| Trait | F | F Sign. | F | F Sign. | F | F Sign. | |
| AUDPC for general resistance | 1.5 | ns | 24.8 | ** | 1.0 | ns | |
| Fusarium damaged kernels | 53.6 | ** | 17.2 | ** | 1.7 | ** | |
| Trichothecenes and their derivatives | Deoxynivalenol | 66.0 | ** | 16.8 | ** | 1.1 | ns |
| DON-3-glucoside | 96.3 | ** | 13.5 | ** | 1.7 | ** | |
| 3-acetyldeoxynivalenol | 75.5 | ** | 6.3 | ** | 0.9 | ns | |
| 15-acetyldeoxynivalenol | 72.9 | ** | 7.2 | ** | 1.5 | * | |
| Nivalenol | 29.7 | ** | 13.4 | ** | 1.2 | ns | |
| T-2 toxin | 10.9 | ** | 1.4 | ns | 0.9 | ns | |
| HT-2 toxin | 9.6 | ** | 3.0 | ** | 0.9 | ns | |
| HT-2 glucoside | 1.9 | ns | 1.3 | ns | 0.8 | ns | |
| Zearelenoneand its derivatives | Zearalenone | 14.0 | ** | 10.9 | ** | 1.0 | ns |
| Zearalenone-sulphate | 51.3 | ** | 11.1 | ** | 2.7 | ** | |
| α-zearalenol | 6.3 | ** | 2.5 | ** | 0.8 | ns | |
| β-zearalenol | 4.4 | * | 2.8 | ** | 0.6 | ns | |
| Emerging mycotoxins | Moniliformin | 1.0 | ns | 2.9 | ** | 0.6 | ns |
| Beauvericin | 81.3 | ** | 8.7 | ** | 2.4 | ** | |
| Enniatin A | 17.6 | ** | 4.9 | ** | 1.6 | * | |
| Enniatin A1 | 18.9 | ** | 5.4 | ** | 1.8 | ** | |
| Enniatin B | 7.8 | ** | 4.7 | ** | 1.1 | ns | |
| Enniatin B1 | 17.4 | ** | 6.0 | ** | 1.6 | * | |
| Enniatin B2 | 5.3 | ** | 4.1 | ** | 0.9 | ns | |
| Enniatin B3 | 2.6 | ns | 2.8 | ** | 0.9 | ns | |
| Other Fusarium metabolites | Culmorin | 134.5 | ** | 15.6 | ** | 4.9 | ** |
| 5-hydroxyculmorin | 11.9 | ** | 4.1 | ** | 0.7 | ns | |
| 15-hydroxyculmorin | 12.5 | ** | 4.0 | ** | 0.6 | ns | |
| 15-hydroxyculmoron | 20.2 | ** | 3.5 | ** | 0.7 | ns | |
| Aurofusarin | 154.6 | ** | 16.2 | ** | 5.9 | ** | |
| Apicidin | 3.0 | ** | 1.8 | * | 0.7 | ns | |
| Chrysogin | 10.8 | ** | 6.3 | ** | 0.8 | ns | |
| Equisetin | 3.3 | * | 1.1 | ns | 0.9 | ns | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spanic, V.; Maricevic, M.; Ikic, I.; Sulyok, M.; Sarcevic, H. Three-Year Survey of Fusarium Multi-Metabolites/Mycotoxins Contamination in Wheat Samples in Potentially Epidemic FHB Conditions. Agronomy 2023, 13, 805. https://doi.org/10.3390/agronomy13030805
Spanic V, Maricevic M, Ikic I, Sulyok M, Sarcevic H. Three-Year Survey of Fusarium Multi-Metabolites/Mycotoxins Contamination in Wheat Samples in Potentially Epidemic FHB Conditions. Agronomy. 2023; 13(3):805. https://doi.org/10.3390/agronomy13030805
Chicago/Turabian StyleSpanic, Valentina, Marko Maricevic, Ivica Ikic, Michael Sulyok, and Hrvoje Sarcevic. 2023. "Three-Year Survey of Fusarium Multi-Metabolites/Mycotoxins Contamination in Wheat Samples in Potentially Epidemic FHB Conditions" Agronomy 13, no. 3: 805. https://doi.org/10.3390/agronomy13030805